Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth


Abstract
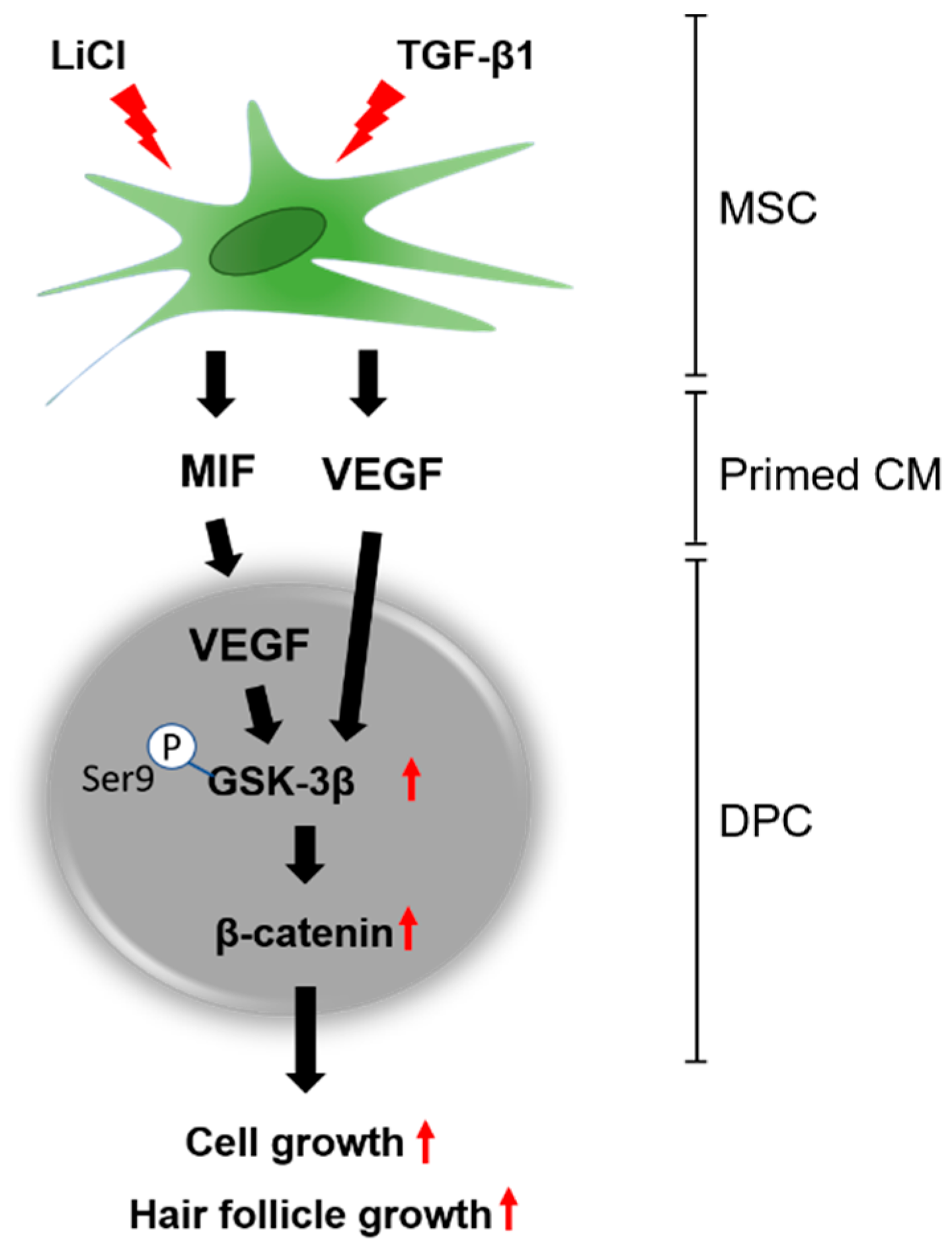
:1. Introduction
2. Materials and Methods
2.1. hUCB-MSC Culture and CM Preparation
2.2. Culture of Follicle Dermal Papilla Cells
2.3. Cell Viability Assay
2.4. Western Blot Analysis
2.5. Growth Factor Array
2.6. L507 Antibody Array
2.7. ELISA
2.8. Neutralizing Antibody
2.9. Clinical Study
2.10. Flow Cytometry
2.11. Statistical Analysis
3. Results
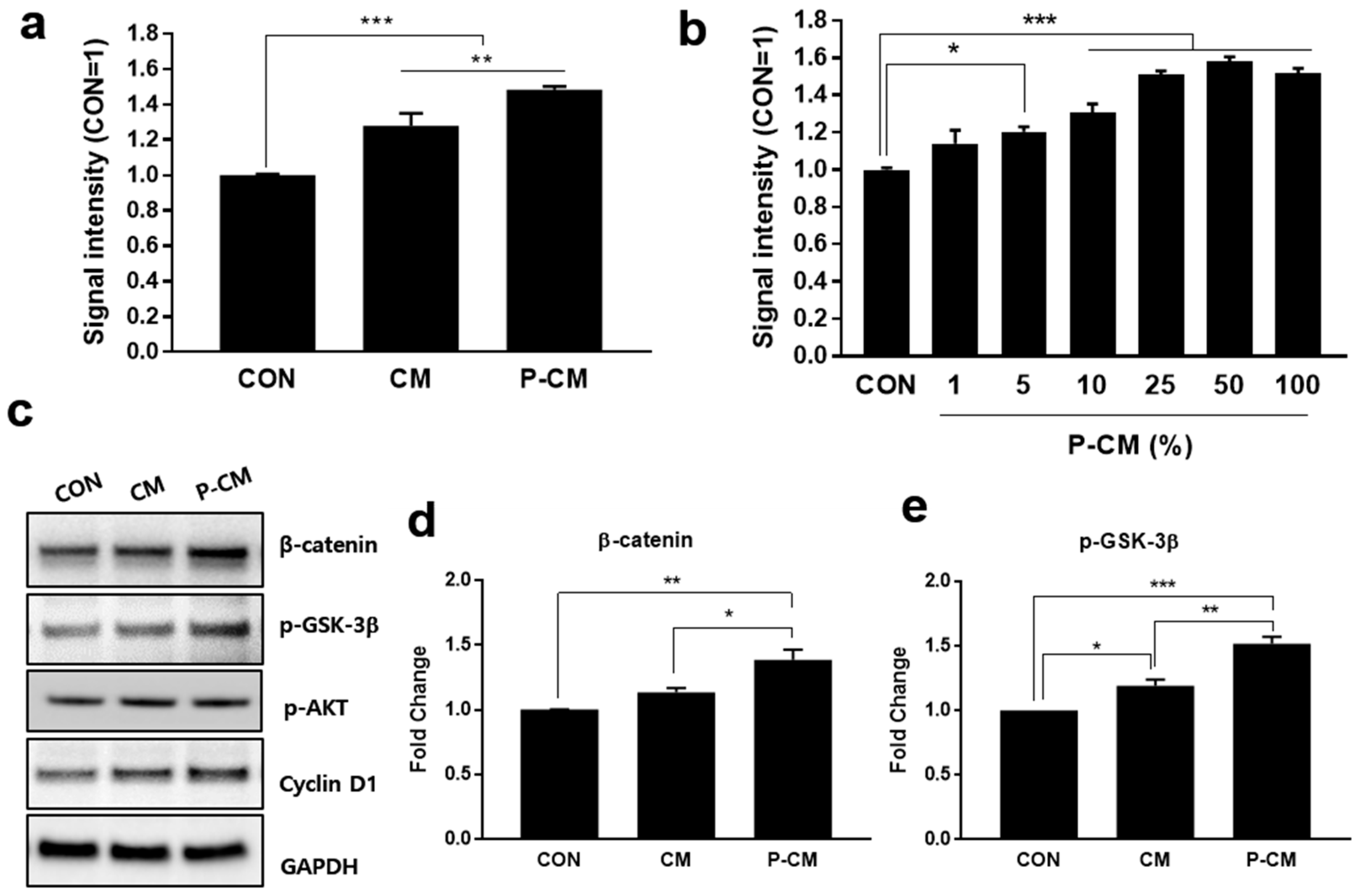
3.1. Primed CM Promoted the Cell Viability of DPCs
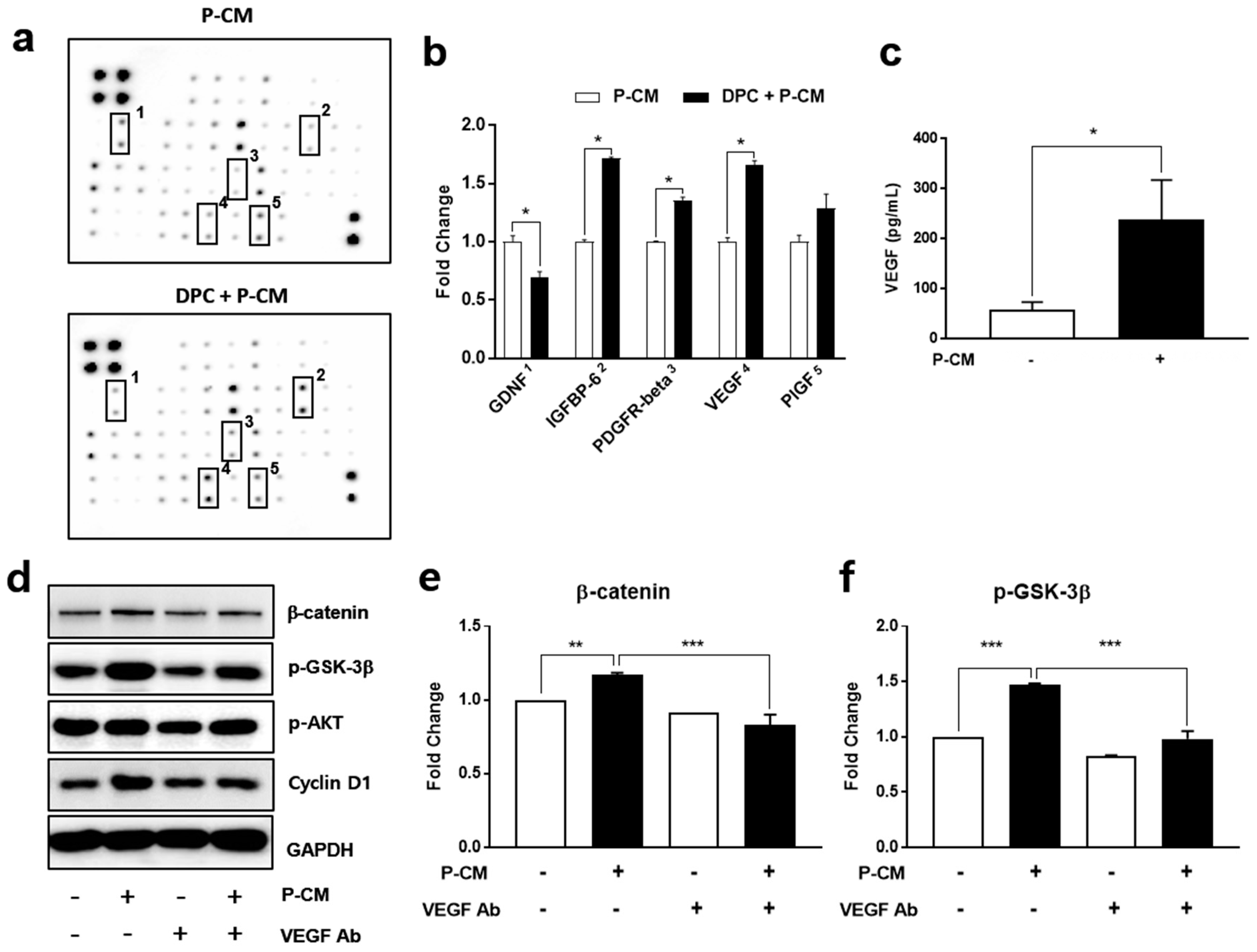
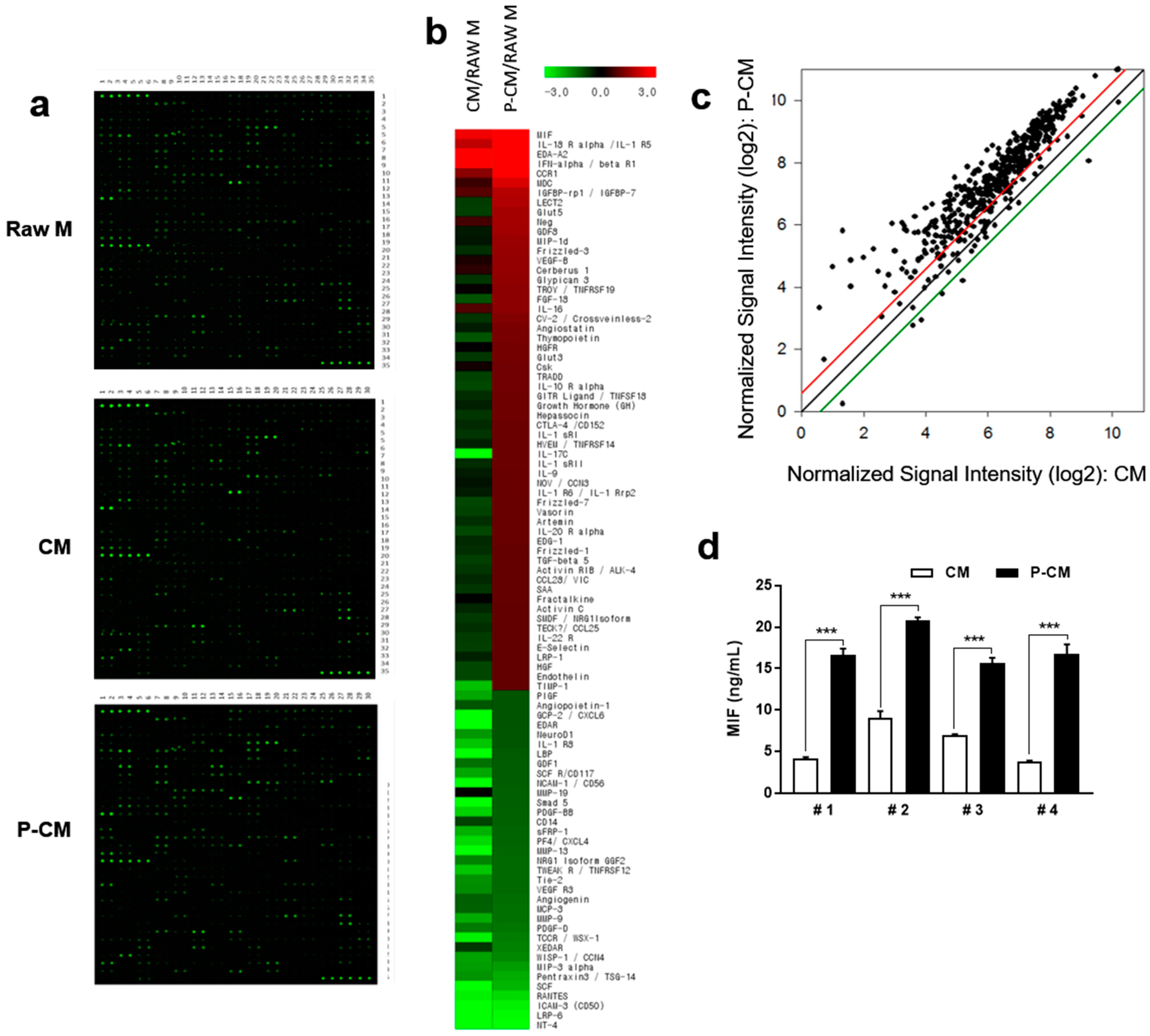
3.2. P-CM Enhanced the Secretion of Growth Factors in DPCs
3.3. MIF Expression was Increased in MSCs with Priming
3.4. MIF-Regulated P-CM Induced VEGF Secretion
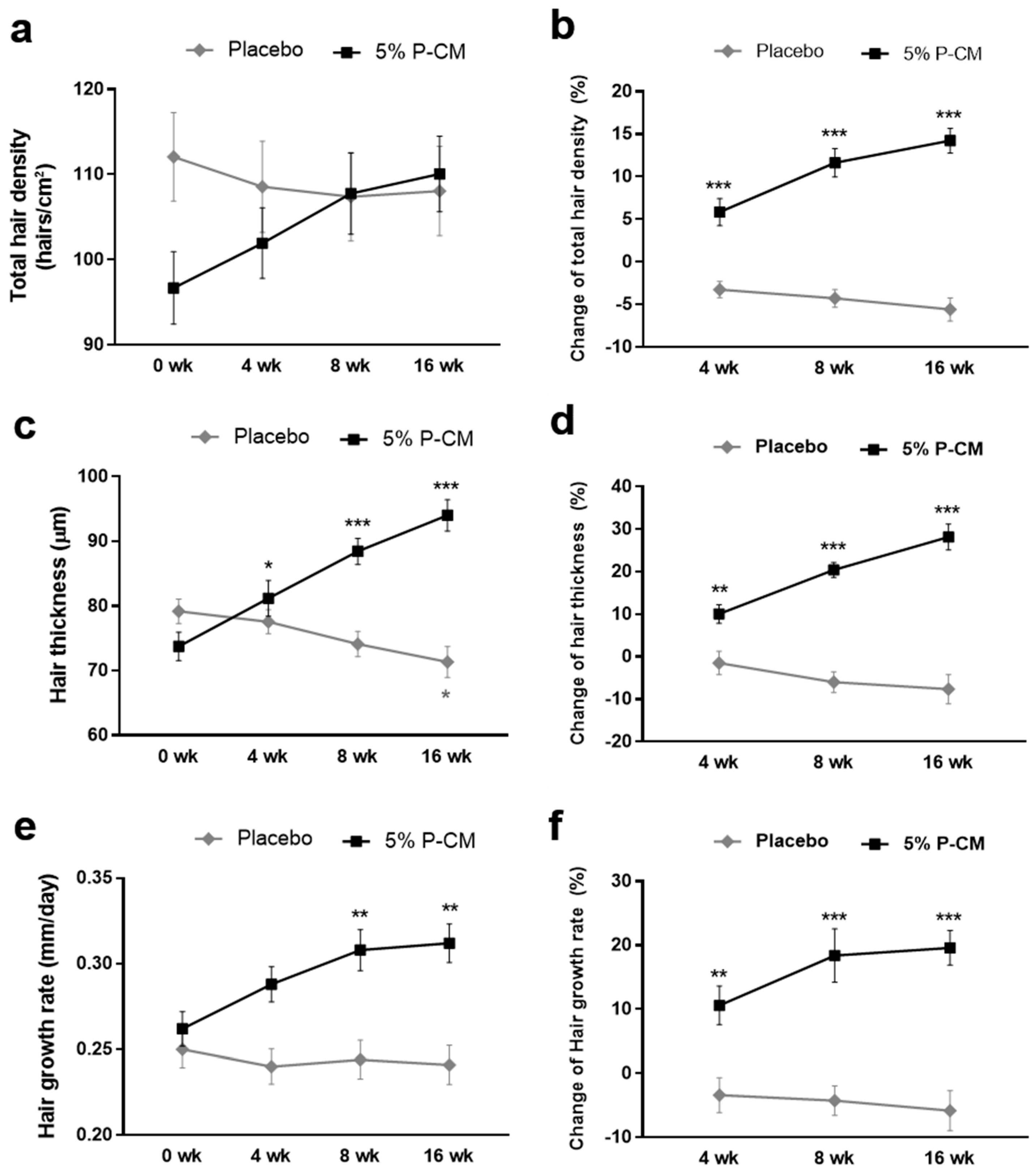
3.5. Topical Agents Containing P-CM Improved Androgenetic Alopecia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nalluri, R.; Harries, M. Alopecia in general medicine. Clin. Med. 2016, 16, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vary, J.C., Jr. Selected Disorders of Skin Appendages—Acne, Alopecia, Hyperhidrosis. Med. Clin. N. Am. 2015, 99, 1195–1211. [Google Scholar] [CrossRef]
- Price, V.H. Treatment of hair loss. N. Engl. J. Med. 1999, 341, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Wipf, A.; Boysen, N.; Hordinsky, M.K.; Dando, E.E.; Sadick, N.; Farah, R.S. The rise of transcutaneous drug delivery for the management of alopecia: A review of existing literature and an eye towards the future. J. Cosmet. Laser Ther. 2019, 21, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Alsalhi, W.; Alalola, A.; Randolph, M.; Gwillim, E.; Tosti, A. Novel drug delivery approaches for the management of hair loss. Expert Opin. Drug Deliv. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sattur, S.S. A review of surgical methods (excluding hair transplantation) and their role in hair loss management today. J. Cutan. Aesthet. Surg. 2011, 4, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Cantisani, C.; Melis, L.; Iorio, A.; Scali, E.; Calvieri, S. Minoxidil use in dermatology, side effects and recent patents. Recent Pat. Inflamm. Allergy Drug Discov. 2012, 6, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Irwig, M.S.; Kolukula, S. Persistent sexual side effects of finasteride for male pattern hair loss. J. Sex. Med. 2011, 8, 1747–1753. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.C.; Cotsarelis, G. Review of hair follicle dermal cells. J. Dermatol. Sci. 2010, 57, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, W.; Abbasi, S.; Hagner, A.; Raharjo, E.; Kumar, R.; Hotta, A.; Magness, S.; Metzger, D.; Biernaskie, J. Hair follicle dermal stem cells regenerate the dermal sheath, repopulate the dermal papilla, and modulate hair type. Dev. Cell 2014, 31, 543–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botchkarev, V.A.; Kishimoto, J. Molecular control of epithelial-mesenchymal interactions during hair follicle cycling. J. Investig. Dermatol. Symp. Proc. 2003, 8, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sennett, R.; Rendl, M. Mesenchymal-epithelial interactions during hair follicle morphogenesis and cycling. Semin. Cell Dev. Biol. 2012, 23, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solanas, G.; Benitah, S.A. Regenerating the skin: A task for the heterogeneous stem cell pool and surrounding niche. Nat. Rev. Mol. Cell Biol. 2013, 14, 737–748. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Shi, C.; Wang, Y.; Yang, J.; Yang, T. Regulatory effect of β-catenin on proliferation of hair follicle stem cells involves PI3K/Akt pathway. J. Appl. Biomed. 2013, 11, 131–141. [Google Scholar] [CrossRef]
- Gilhar, A.; Etzioni, A.; Paus, R. Alopecia areata. N. Engl. J. Med. 2012, 366, 1515–1525. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Xie, N.; Li, W.; Yuan, B.; Shi, Y.; Wang, Y. Immunobiology of mesenchymal stem cells. Cell Death Differ. 2014, 21, 216–225. [Google Scholar] [CrossRef]
- Allan, D.S. Using umbilical cord blood for regenerative therapy: Proof or promise? Stem Cells 2020. [Google Scholar] [CrossRef]
- Berglund, S.; Magalhaes, I.; Gaballa, A.; Vanherberghen, B.; Uhlin, M. Advances in umbilical cord blood cell therapy: The present and the future. Expert Opin. Biol. Ther. 2017, 17, 691–699. [Google Scholar] [CrossRef]
- Bak, D.H.; Choi, M.J.; Kim, S.R.; Lee, B.C.; Kim, J.M.; Jeon, E.S.; Oh, W.; Lim, E.S.; Park, B.C.; Kim, M.J.; et al. Human umbilical cord blood mesenchymal stem cells engineered to overexpress growth factors accelerate outcomes in hair growth. Korean J. Physiol. Pharmacol. 2018, 22, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Pawitan, J.A. Prospect of stem cell conditioned medium in regenerative medicine. BioMed Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, H.; Suga, H. Hair Regeneration Treatment Using Adipose-Derived Stem Cell Conditioned Medium: Follow-up With Trichograms. Eplasty 2015, 15, e10. [Google Scholar] [PubMed]
- Dong, L.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Tong, C.; Hou, Q.; Han, Q.; Zhao, Y.; Liu, H.; et al. Treatment of MSCs with Wnt1a-conditioned medium activates DP cells and promotes hair follicle regrowth. Sci. Rep. 2014, 4, 5432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.S.; Kim, W.S.; Choi, J.S.; Kim, H.K.; Won, J.H.; Ohkubo, F.; Fukuoka, H. Hair growth stimulated by conditioned medium of adipose-derived stem cells is enhanced by hypoxia: Evidence of increased growth factor secretion. Biomed. Res. 2010, 31, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Won, C.H.; Yoo, H.G.; Kwon, O.S.; Sung, M.Y.; Kang, Y.J.; Chung, J.H.; Park, B.S.; Sung, J.H.; Kim, W.S.; Kim, K.H. Hair growth promoting effects of adipose tissue-derived stem cells. J. Dermatol. Sci. 2010, 57, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, S.L.; Hsiao, S.T.; Peshavariya, H.M.; Lim, S.Y.; Dusting, G.J.; Dilley, R.J. Hypoxic preconditioning enhances survival of human adipose-derived stem cells and conditions endothelial cells in vitro. Stem Cells Dev. 2012, 21, 1887–1896. [Google Scholar] [CrossRef]
- Di Santo, S.; Yang, Z.; Wyler von Ballmoos, M.; Voelzmann, J.; Diehm, N.; Baumgartner, I.; Kalka, C. Novel cell-free strategy for therapeutic angiogenesis: In vitro generated conditioned medium can replace progenitor cell transplantation. PLoS ONE 2009, 4, e5643. [Google Scholar] [CrossRef] [Green Version]
- Song, L.L.; Cui, Y.; Yu, S.J.; Liu, P.G.; He, J.F. TGF-beta and HSP70 profiles during transformation of yak hair follicles from the anagen to catagen stage. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef]
- Tsuji, Y.; Denda, S.; Soma, T.; Raftery, L.; Momoi, T.; Hibino, T. A potential suppressor of TGF-beta delays catagen progression in hair follicles. J. Investig. Dermatol. Symp. Proc. 2003, 8, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Yoon, J.; Shin, S.H.; Zahoor, M.; Kim, H.J.; Park, P.J.; Park, W.S.; Min do, S.; Kim, H.Y.; Choi, K.Y. Valproic acid induces hair regeneration in murine model and activates alkaline phosphatase activity in human dermal papilla cells. PLoS ONE 2012, 7, e34152. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Hao, C.M.; Breyer, M.D. Hypertonic stress activates glycogen synthase kinase 3beta-mediated apoptosis of renal medullary interstitial cells, suppressing an NFkappaB-driven cyclooxygenase-2-dependent survival pathway. J. Biol. Chem. 2004, 279, 3949–3955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Huang, L.D.; Jiang, Y.M.; Manji, H.K. The mood-stabilizing agent valproate inhibits the activity of glycogen synthase kinase-3. J. Neurochem. 1999, 72, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Kim, M.; Bae, Y.K.; Kim, G.H.; Choi, S.J.; Oh, W.; Um, S.; Jin, H.J. Decorin Secreted by Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Induces Macrophage Polarization via CD44 to Repair Hyperoxic Lung Injury. Int. J. Mol. Sci. 2019, 20, 4815. [Google Scholar] [CrossRef] [Green Version]
- Chikhalkar, S.; Jerajani, H.; Madke, B. Evaluation of utility of phenol in alopecia areata. Int. J. Trichology 2013, 5, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Yang, Z.; Wu, Y.; Denslin, V.; Yu, C.C.; Tee, C.A.; Lim, C.T.; Han, J.; Lee, E.H. Label-free separation of mesenchymal stem cell subpopulations with distinct differentiation potencies and paracrine effects. Biomaterials 2020, 240, 119881. [Google Scholar] [CrossRef]
- Peng, Y.; Xuan, M.; Zou, J.; Liu, H.; Zhuo, Z.; Wan, Y.; Cheng, B. Freeze-dried rat bone marrow mesenchymal stem cell paracrine factors: A simplified novel material for skin wound therapy. Tissue Eng. Part A 2015, 21, 1036–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvolini, E.; Orciani, M.; Vignini, A.; Mattioli-Belmonte, M.; Mazzanti, L.; Di Primio, R. Skin-derived mesenchymal stem cells (S-MSCs) induce endothelial cell activation by paracrine mechanisms. Exp. Dermatol. 2010, 19, 848–850. [Google Scholar] [CrossRef]
- Golchin, A.; Farahany, T.Z.; Khojasteh, A.; Soleimanifar, F.; Ardeshirylajimi, A. The Clinical Trials of Mesenchymal Stem Cell Therapy in Skin Diseases: An Update and Concise Review. Curr. Stem Cell Res. Ther. 2019, 14, 22–33. [Google Scholar] [CrossRef]
- Zimber, M.P.; Ziering, C.; Zeigler, F.; Hubka, M.; Mansbridge, J.N.; Baumgartner, M.; Hubka, K.; Kellar, R.; Perez-Meza, D.; Sadick, N.; et al. Hair regrowth following a Wnt- and follistatin containing treatment: Safety and efficacy in a first-in-man phase 1 clinical trial. J. Drugs Dermatol. 2011, 10, 1308–1312. [Google Scholar]
- Inui, S.; Itami, S. Androgen receptor transactivity is potentiated by TGF-beta1 through Smad3 but checked by its coactivator Hic-5/ARA55 in balding dermal papilla cells. J. Dermatol. Sci. 2011, 64, 149–151. [Google Scholar] [CrossRef]
- Qi, W.; Gao, S.; Chu, J.; Zhou, L.; Wang, Z. Negative androgen-response elements mediate androgen-dependent transcriptional inhibition of TGF-beta1 and CDK2 promoters in the prostate gland. J. Androl. 2012, 33, 27–36. [Google Scholar] [CrossRef]
- Inui, S.; Fukuzato, Y.; Nakajima, T.; Yoshikawa, K.; Itami, S. Androgen-inducible TGF-beta1 from balding dermal papilla cells inhibits epithelial cell growth: A clue to understand paradoxical effects of androgen on human hair growth. FASEB J. 2002, 16, 1967–1969. [Google Scholar] [CrossRef]
- Choi, Y.M.; Choi, S.Y.; Kim, H.; Kim, J.; Ki, M.S.; An, I.-S.; Jung, J. TGFβ family mimetic peptide promotes proliferation of human hair follicle dermal papilla cells and hair growth in C57BL/6 mice. Biomed. Dermatol. 2018, 2. [Google Scholar] [CrossRef]
- Xia, M.Y.; Zhao, X.Y.; Huang, Q.L.; Sun, H.Y.; Sun, C.; Yuan, J.; He, C.; Sun, Y.; Huang, X.; Kong, W.; et al. Activation of Wnt/beta-catenin signaling by lithium chloride attenuates d-galactose-induced neurodegeneration in the auditory cortex of a rat model of aging. FEBS Open Bio 2017, 7, 759–776. [Google Scholar] [CrossRef]
- Shan, T.; Zhou, C.; Yang, R.; Yan, F.; Zhang, P.; Fu, Y.; Jiang, H. Lithium chloride promotes the odontoblast differentiation of hair follicle neural crest cells by activating Wnt/beta-catenin signaling. Cell Biol. Int. 2015, 39, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Oshimori, N.; Fuchs, E. Paracrine TGF-beta signaling counterbalances BMP-mediated repression in hair follicle stem cell activation. Cell Stem Cell 2012, 10, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.Y.; Yang, L.T. Differential response of epithelial stem cell populations in hair follicles to TGF-beta signaling. Dev. Biol. 2013, 373, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.I.; Kim, S.C.; Kim, M.K.; Boo, H.J.; Kim, E.J.; Im, G.J.; Kim, Y.H.; Hyun, J.W.; Kang, J.H.; Koh, Y.S.; et al. Effects of dihydrotestosterone on rat dermal papilla cells in vitro. Eur. J. Pharmacol. 2015, 757, 74–83. [Google Scholar] [CrossRef]
- Leiros, G.J.; Attorresi, A.I.; Balana, M.E. Hair follicle stem cell differentiation is inhibited through cross-talk between Wnt/beta-catenin and androgen signalling in dermal papilla cells from patients with androgenetic alopecia. Br. J. Dermatol. 2012, 166, 1035–1042. [Google Scholar] [CrossRef]
- Kwon, T.R.; Oh, C.T.; Choi, E.J.; Park, H.M.; Han, H.J.; Ji, H.J.; Kim, B.J. Human placental extract exerts hair growth-promoting effects through the GSK-3beta signaling pathway in human dermal papilla cells. Int. J. Mol. Med. 2015, 36, 1088–1096. [Google Scholar] [CrossRef] [Green Version]
- Lowry, W.E.; Blanpain, C.; Nowak, J.A.; Guasch, G.; Lewis, L.; Fuchs, E. Defining the impact of beta-catenin/Tcf transactivation on epithelial stem cells. Genes Dev. 2005, 19, 1596–1611. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.Y.; Yoon, J.S.; Jo, S.J.; Shin, C.Y.; Shin, J.Y.; Kim, J.I.; Kwon, O.; Kim, K.H. A role of placental growth factor in hair growth. J. Dermatol. Sci. 2014, 74, 125–134. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Bassino, E.; Zanardi, A.; Gasparri, F.; Munaron, L. Effects of the biomimetic peptide Sh-Polypeptide 9 (CG-VEGF) on cocultures of human hair follicle dermal papilla cells and microvascular endothelial cells. Exp. Dermatol. 2016, 25, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Man, X.Y.; Li, C.M.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Lu, Z.F.; Zheng, M. VEGF induces proliferation of human hair follicle dermal papilla cells through VEGFR-2-mediated activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Fernandez, J.G.; Rodriguez, D.A.; Valenzuela, M.; Calderon, C.; Urzua, U.; Munroe, D.; Rosas, C.; Lemus, D.; Diaz, N.; Wright, M.C.; et al. Survivin expression promotes VEGF-induced tumor angiogenesis via PI3K/Akt enhanced beta-catenin/Tcf-Lef dependent transcription. Mol. Cancer 2014, 13, 209. [Google Scholar] [CrossRef] [Green Version]
- Easwaran, V.; Lee, S.H.; Inge, L.; Guo, L.; Goldbeck, C.; Garrett, E.; Wiesmann, M.; Garcia, P.D.; Fuller, J.H.; Chan, V.; et al. beta-Catenin regulates vascular endothelial growth factor expression in colon cancer. Cancer Res. 2003, 63, 3145–3153. [Google Scholar]
- Salem, S.A.; Asaad, M.K.; Elsayed, S.B.; Sehsah, H.M. Evaluation of macrophage migration inhibitory factor (MIF) levels in serum and lesional skin of patients with alopecia areata. Int. J. Dermatol. 2016, 55, 1357–1361. [Google Scholar] [CrossRef]
- Younan, D.N.; Agamia, N.; Elshafei, A.; Ebeid, N. Serum level of macrophage migration inhibitory factor (MIF) in Egyptians with alopecia areata and its relation to the clinical severity of the disease. J. Clin. Lab. Anal. 2015, 29, 74–79. [Google Scholar] [CrossRef]
- Trivedi-Parmar, V.; Jorgensen, W.L. Advances and Insights for Small Molecule Inhibition of Macrophage Migration Inhibitory Factor. J. Med. Chem. 2018, 61, 8104–8119. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated in P-CM Normalized to Raw Medium | Downregulated in P-CM Normalized to Raw Medium | ||||||
|---|---|---|---|---|---|---|---|
| Protein | CM | P-CM | P-CM/CM Ratio | Protein | CM | P-CM | P-CM/CM Ratio |
| Normalized to Raw Medium | Normalized to Raw Medium | ||||||
| MIF | 5.50 | 22.62 | 4.11 | PlGF | 0.26 | 0.50 | 1.96 |
| IL-18 R alpha/IL-1 R5 | 3.87 | 8.90 | 2.30 | Angiopoietin-1 | 0.49 | 0.50 | 1.02 |
| EDA-A2 | 9.72 | 8.10 | 0.83 | GCP-2/CXCL6 | 0.07 | 0.50 | 6.67 |
| IFN-alpha/beta R1 | 14.50 | 7.66 | 0.53 | EDAR | 0.04 | 0.50 | 12.50 |
| CCR1 | 2.64 | 5.97 | 2.26 | NeuroD1 | 0.29 | 0.49 | 1.72 |
| MDC | 1.56 | 4.65 | 2.99 | IL-1 R8 | 0.20 | 0.49 | 2.49 |
| IGFBP-rp1/IGFBP-7 | 1.90 | 3.58 | 1.89 | LBP | 0.05 | 0.48 | 10.35 |
| LECT2 | 0.60 | 3.46 | 5.77 | GDF1 | 0.40 | 0.47 | 1.18 |
| Glut5 | 0.59 | 3.18 | 5.36 | SCF R/CD117 | 0.26 | 0.47 | 1.83 |
| GDF8 | 0.78 | 3.11 | 1.90 | NCAM-1/CD56 | 0.10 | 0.47 | 4.49 |
| MIP-1d | 0.80 | 2.95 | 3.97 | MMP-19 | 1.06 | 0.46 | 0.44 |
| Frizzled-3 | 0.66 | 2.88 | 3.67 | Smad 5 | 0.06 | 0.46 | 7.72 |
| VEGF-B | 1.28 | 2.82 | 4.34 | PDGF-BB | 0.18 | 0.45 | 2.52 |
| Cerberus 1 | 1.44 | 2.79 | 2.21 | CD14 | 0.57 | 0.45 | 0.79 |
| Glypican 3 | 0.64 | 2.75 | 1.94 | sFRP-1 | 0.24 | 0.44 | 1.85 |
| TROY/TNFRSF19 | 1.10 | 2.71 | 4.29 | PF4/CXCL4 | 0.17 | 0.44 | 2.63 |
| FGF-18 | 0.51 | 2.69 | 2.45 | MMP-13 | 0.02 | 0.43 | 25.54 |
| IL-16 | 1.83 | 2.69 | 5.27 | NRG1 | 0.34 | 0.42 | 1.23 |
| Crossveinless-2 | 0.64 | 2.50 | 1.47 | TWEAK R | 0.20 | 0.42 | 2.10 |
| Angiostatin | 0.76 | 2.37 | 3.91 | Tie-2 | 0.31 | 0.42 | 1.36 |
| Thymopoietin | 0.50 | 2.32 | 3.12 | VEGF R3 | 0.32 | 0.42 | 1.32 |
| HGFR | 1.05 | 2.29 | 4.65 | Angiogenin | 0.46 | 0.41 | 0.90 |
| Glut3 | 0.62 | 2.28 | 2.19 | MCP-3 | 0.45 | 0.40 | 0.90 |
| Csk | 1.21 | 2.21 | 3.69 | MMP-9 | 0.25 | 0.40 | 1.59 |
| TRADD | 0.58 | 2.21 | 1.82 | PDGF-D | 0.37 | 0.38 | 1.01 |
| IL-10 R alpha | 0.55 | 2.21 | 3.80 | TCCR/WSX-1 | 0.14 | 0.36 | 2.51 |
| GITR Ligand | 0.68 | 2.19 | 4.01 | XEDAR | 0.62 | 0.35 | 0.56 |
| Growth Hormone | 0.75 | 2.18 | 3.22 | WISP-1/CCN4 | 0.27 | 0.33 | 1.21 |
| Hepassocin | 0.63 | 2.17 | 2.88 | MIP-3 alpha | 0.28 | 0.28 | 1.01 |
| CTLA-4/CD152 | 0.69 | 2.15 | 3.45 | Pentraxin3 | 0.30 | 0.25 | 0.83 |
| IL-1 sRI | 0.64 | 2.15 | 3.10 | SCF | 0.03 | 0.23 | 8.86 |
| HVEM/TNFRSF14 | 0.74 | 2.15 | 3.35 | RANTES | 0.14 | 0.17 | 1.18 |
| IL-17C | 0.10 | 2.14 | 2.92 | ICAM-3 (CD50) | 0.12 | 0.14 | 1.18 |
| IL-1 sRII | 0.68 | 2.14 | 22.31 | LRP-6 | 0.02 | 0.13 | 8.22 |
| IL-9 | 0.76 | 2.13 | 3.16 | NT-4 | 0.10 | 0.11 | 1.18 |
| NOV/CCN3 | 0.82 | 2.13 | 2.81 | ||||
| IL-1 R6/IL-1 Rrp2 | 0.76 | 2.11 | 2.60 | ||||
| Frizzled-7 | 0.56 | 2.10 | 2.76 | ||||
| Vasorin | 0.58 | 2.10 | 3.78 | ||||
| Artemin | 0.67 | 2.09 | 3.64 | ||||
| IL-20 R alpha | 0.56 | 2.09 | 3.12 | ||||
| EDG-1 | 0.69 | 2.09 | 3.71 | ||||
| Frizzled-1(FZD1) | 0.66 | 2.06 | 3.03 | ||||
| TGF-beta 5 | 0.59 | 2.06 | 3.11 | ||||
| Activin RIB/ALK-4 | 0.63 | 2.06 | 3.51 | ||||
| CCL28/VIC | 0.70 | 2.06 | 3.28 | ||||
| SAA | 0.63 | 2.05 | 2.95 | ||||
| Fractalkine | 0.96 | 2.05 | 3.27 | ||||
| Activin C | 0.70 | 2.03 | 2.12 | ||||
| SMDF/NRG1 Isoform | 0.61 | 2.03 | 2.91 | ||||
| TECK/CCL25 | 0.67 | 2.02 | 3.34 | ||||
| IL-22 R | 0.60 | 2.02 | 3.03 | ||||
| E-Selectin | 0.59 | 2.02 | 3.36 | ||||
| LRP-1 | 0.73 | 2.01 | 3.45 | ||||
| HGF | 0.56 | 2.00 | 2.76 | ||||
| Endothelin | 0.56 | 2.00 | 3.58 | ||||
| TIMP-1 | 0.21 | 2.00 | 9.66 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, H.A.; Kwak, J.; Kim, B.J.; Jin, H.J.; Park, W.S.; Choi, S.J.; Oh, W.; Um, S. Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth. Cells 2020, 9, 1344. https://doi.org/10.3390/cells9061344
Oh HA, Kwak J, Kim BJ, Jin HJ, Park WS, Choi SJ, Oh W, Um S. Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth. Cells. 2020; 9(6):1344. https://doi.org/10.3390/cells9061344
Chicago/Turabian StyleOh, Hyun Ah, Jihye Kwak, Beom Joon Kim, Hye Jin Jin, Won Seok Park, Soo Jin Choi, Wonil Oh, and Soyoun Um. 2020. "Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth" Cells 9, no. 6: 1344. https://doi.org/10.3390/cells9061344