Activation of 3-Mercaptopyruvate Sulfurtransferase by Glutaredoxin Reducing System

Isotope Research Laboratory, Nippon Medical School, 1-1-5 Sendagi Bunkyo-Ku, Tokyo 113-8602, Japan
Biomolecules 2020, 10(6), 826; https://doi.org/10.3390/biom10060826
Submission received: 26 March 2020
/
Revised: 22 May 2020
/
Accepted: 27 May 2020
/
Published: 28 May 2020
(This article belongs to the Special Issue H2S, Polysulfides, and Enzymes: Physiological and Pathological Aspects)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Glutaredoxin (EC 1.15–1.21) is known as an oxidoreductase that protects cysteine residues within proteins against oxidative stress. Glutaredoxin catalyzes an electron transfer reaction that donates an electron to substrate proteins in the reducing system composed of glutaredoxin, glutathione, glutathione reductase, and nicotinamide-adenine dinucleotide phosphate (reduced form). 3-mercaptopyruvate sulfurtransferase (EC 2.8.1.2) is a cysteine enzyme that catalyzes transsulfuration, and glutaredoxin activates 3-mercaptopyruvate sulfurtransferase in the reducing system. Interestingly, even when glutathione or glutathione reductase was absent, 3-mercaptopyruvate sulfurtransferase activity increased, probably because reduced glutaredoxin was partly present and able to activate 3-mercaptopyruvate sulfurtransferase until depletion. A study using mutant Escherichia coli glutaredoxin1 (Cys14 is the binding site of glutathione and was replaced with a Ser residue) confirmed these results. Some inconsistency was noted, and glutaredoxin with higher redox potential than either 3-mercaptopyruvate sulfurtransferase or glutathione reduced 3-mercaptopyruvate sulfurtransferase. However, electron-transfer enzymatically proceeded from glutaredoxin to 3-mercaptopyruvate sulfurtransferase.
1. Introduction
Glutaredoxin (Grx) was first discovered by Holmgren [1,2,3]. Grx is an enzyme and is categorized as an oxidoreductase (EC 1.15–1.21) that catalyzes not only the reduction process of a disulfide bond on the substrate protein but also cleavage of glutathione (GSH) from the GSH-disulfide complex, meaning that Grx enzymatically donates electrons to substrate proteins; the full reaction mechanism has been clarified [4,5,6,7]. Protein substrates are reduced by Grx, and then oxidized Grx is non-enzymatically reduced by GSH, a redox molecule. Oxidized GSH is enzymatically reduced by glutathione reductase (GRD) together with nicotinamide-adenine dinucleotide phosphate in its reduced form (NADPH) as a coenzyme. Many eukaryotic and prokaryotic subtypes of Grx have been reported; among these subtypes, a ternary structure for Escherichia coli Grx1 has been reported [8]. This study clarified that Cys14 was not only a binding site for GSH but also the site of a disulfide bond with Cys11. These facts were confirmed by a study using mutant E. coli Grx1 (Cys14 was replaced with Ser) [9]. As a redox protein like Grx, thioredoxin (Trx) is well studied and is a smaller molecule with lower redox potential. Trx also reduces substrate proteins associated with Trx reductase (TRD).
3-mercaptopyruvate sulfurtransferase (MST, EC 2.8.1.2) catalyzes a transsulfuration. Recently, it has been revealed that MST produces hydrogen sulfide and polysulfides [10,11,12,13,14,15,16,17,18] and plays important roles in living organisms [19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43]. MST is activated by the Trx-TRD, but not the GSH-GRD, reducing system due to differences in redox potential [44,45,46,47]. There are two target points for the reduction in MST; a sulfenic acid formed at the catalytic site cysteine and a disulfide bond in dimeric MST [44,45,46,47]. MST activity is regulated via redox-dependent changes at these molecular points at the post-translational level [44,45,46,47]. In this experiment, we proved that Grx reduced oxidized-MST to activate MST, although the redox potentials of human Grx1 and Grx2 were −232 mV and −222 mV, respectively [48], and that of E. coli Grx1 was −233 mV [49], which was higher than those of GSH (~−240 mV) [48] and Trx (~−270 mV) [38].
2. Materials and Methods
2.1. Overexpression and Preparation of Rat MST
A pET28a vector containing rat MST was introduced in E. coli, BL21(DE3) cells transformed with a pSTV vector containing GroEL and GroES cDNAs. MST was purified according to a procedure described previously [44,45]. The resulting MST was 131 μM in 20 mM in potassium phosphate buffer, pH 7.2, and was frozen until use.
2.2. Preparation of Other Proteins and Chemical Agents
Human Grx1 (IMCO Corporation Ltd AB, Stockholm, Sweden) and human Grx2 (IMCO Corporation Ltd AB, Stockholm, Sweden) were adjusted to 1 mM each using 20 mM potassium phosphate buffer, pH 7.2, as a stock solution. E. coli Grx1 (IMCO Corporation Ltd AB, Stockholm, Sweden) and E. coli mutant Grx (C14S, Cys14 of E. coli Grx1 was replaced with Ser) (IMCO Corporation Ltd AB, Stockholm, Sweden) were adjusted to 3 mM each with 20 mM potassium phosphate buffer, pH 7.2. Human GRD (Sigma-Aldrich, Inc. St. Louis, MO, USA) and E. coli GRD (Novus Biologicals, Centennial, CO, USA) were 304 μM and 19.531 μM, respectively, in their original solutions. NADPH (Sigma-Aldrich, Inc. MO, USA) and GSH (Sigma-Aldrich, Inc.) were adjusted to 10 mM each with potassium phosphate buffer, pH 7.2. Sodium thiosulfate, sodium cyanide, and other chemicals were purchased from Wako Pure Chemicals Industries, Ltd. (Osaka, Japan).
2.3. Grx-GSH-GRD-NADPH Reducing Systems
In the Grx, GSH, GRD, and NADPH systems, all reagents were further diluted with 20 mM potassium phosphate buffer, pH 7.2. MST was diluted to 1/50-fold molar concentration (final concentration; 2.62 μM), human Grx1 to 1/100-fold molar concentration (final 10 μM), human Grx2 to 100-fold molar concentration (final 10 μM), E. coli Grx1 to 1/100-fold molar concentration (final 30 μM), E. coli mutant Grx to 1/100-fold molar concentration (final 30 μM), human GRD to 1/100-fold molar concentration (final 3.04 μM), E. coli GRD to 1/100-fold molar concentration (final 0.2 μM), and NADPH to 1/100-fold molar concentration (final 100 μM). GSH (10 mM) was used without dilution.
The incubation mixtures for the study of activation contained 1 μL of diluted MST solution, 1.6 μL of diluted NADPH solution, 1.2 μL of diluted human Grx1 (or human Grx2) solution, 0.9 μL of human GRD solution, and 1 μL of diluted GSH solution. Other incubation mixtures contained 1 μL of diluted MST solution, 1.6 μL of diluted NADPH solution, 1 μL of diluted E. coli Grx1 (or E. coli Grx1 mutant) solution, 1.3 μL of E. coli GRD solution, and 1 μL of diluted GSH solution. As control experiments, after the incubation of MST with 1.2 mM dithiothreitol (DTT) for 20 min on ice, DTT was removed on a PD 10 (GE Healthcare, Chicago, IL, USA). Then, MST activity was measured. After treatment of human Grx 1 with 1.2 mM DTT for 20 min on ice, DTT was removed on a PD 10, and Grxs were concentrated with VIVASPIN 500, MWCO 5000, PES (Sartorius AG, Goettingen, Germany). Then, MST was mixed with DTT-pretreated Grxs, and MST activity was measured. The total volume of each incubation mixture was adjusted to 20 μL using 20 mM potassium phosphate buffer, pH 7.2.
2.4. Time Dependency of MST Activation
The contents of the incubation mixture were as described above. After incubation at 20 °C for 0, 5, 10, and 15 min, individual 5 μL aliquots were used to measure enzyme activity (rhodanese activity).
2.5. MST Activation Studies Using the Reducing System
2.5.1. Enzyme Activation in Human Grx1 System with or without GSH
- (a)
- Measurements of MST activity under various human Grx1 concentrations without GSH: The incubation mixture contained 0, 0.3, 0.6, 0.9, 1.2, or 1.5 μL of human Grx1 solution in 20 μL of incubation mixture (final concentrations: 0, 0.15. 0.3, 0.45, 0.6, and 0.75 μM, respectively) without GSH solution. After incubation at 20 °C for 10 min, 5 μL aliquots were used to measure enzyme activity (rhodanese activity).
- (b)
- Measurements of MST activity under various NADPH concentrations without GSH: The incubation mixture contained 0, 0.4, 0.8, 1.2, 1.6, or 2.0 μL of NADPH solution in 20 μL of incubation mixture (final concentrations: 0, 2, 4, 6, 8, and 10 μM, respectively) without GSH solution.
- (c)
- Measurements of MST activity under various human GRD concentrations without GSH: The incubation mixture contained 0, 0.3, 0.6, 0.9, 1.2, or 1.5 μL of human GRD solution in 20 μL of incubation mixture (final concentrations: 0, 0.0456, 0.0912, 0.1368, 0.1824, and 0.228 μM, respectively) without GSH solution.
- (d)
- Measurements of MST activity under various GSH concentrations: The incubation mixture contained 0, 0.2, 0.5, 1, 1.5 or 2 μL of GSH solution in 20 μL of incubation mixture (final concentrations: 0, 0.1, 0.25, 0.5, 0.75, and 1 mM, respectively).
2.5.2. Enzyme Activation in a Human Grx2 System with or without GSH
The incubation mixture contained 0, 0.3, 0.6, 0.9, 1.2, or 1.5 μL of human Grx2 solution in 20 μL of incubation mixture (final concentrations: 0, 0.15. 0.3, 0.45, 0.6, and 0.75 μM, respectively) with or without 1 μL of GSH solution.
2.5.3. Enzyme Activation in an E. coli Grx1 System with or without GSH
The incubation mixture contained 0, 0.5, 1, 1.5, 2, or 2.5 μL in 20 μL of E. coli Grx1 solution and 20 μL of incubation mixture (final concentrations: 0, 0.75, 1.5, 2.25, 3, and 3.75 μM, respectively) with or without 1 μL of GSH solution.
2.5.4. Enzyme Activation in an E. coli Grx1 Mutant System with or without GSH
The incubation mixture contained 0, 0.5, 1, 1.5, 2, or 2.5 μL in 20 μL of E. coli Grx1 mutant solution and 20 μL of incubation mixture (final concentrations: 0, 0.75, 1.5, 2.25, 3 and 3.75 μM, respectively) with or without 1 μL of GSH solution.
2.6. Rhodanese Activity of MST
In usual MST activity when catalyzing the trans-sulfuration from mercaptopyruvate to 3-mercaptoethanol, 3-mercaptoethanol can reduce agents including MST during incubation in the assay mixture. As MST possesses rhodanese activity [50,51] via catalyzing the trans-sulfuration from thiosulfate to cyanide, the rhodanese activity of MST was measured in this experiment. All activities were measured in triplicate.
2.7. Protein Concentration
The protein concentration was determined with a Coomassie protein assay kit (Pierce Biotechnology, Rockford, IL, USA) using crystalline bovine serum albumin (ICN Biochemicals, Irvine, CA, USA) as the standard.
2.8. Statistical Analysis
All values are expressed as the mean ± S.D. Significance of difference between values was estimated by a Student’s t-test. A p-value of less than 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Time-Dependent MST Activation
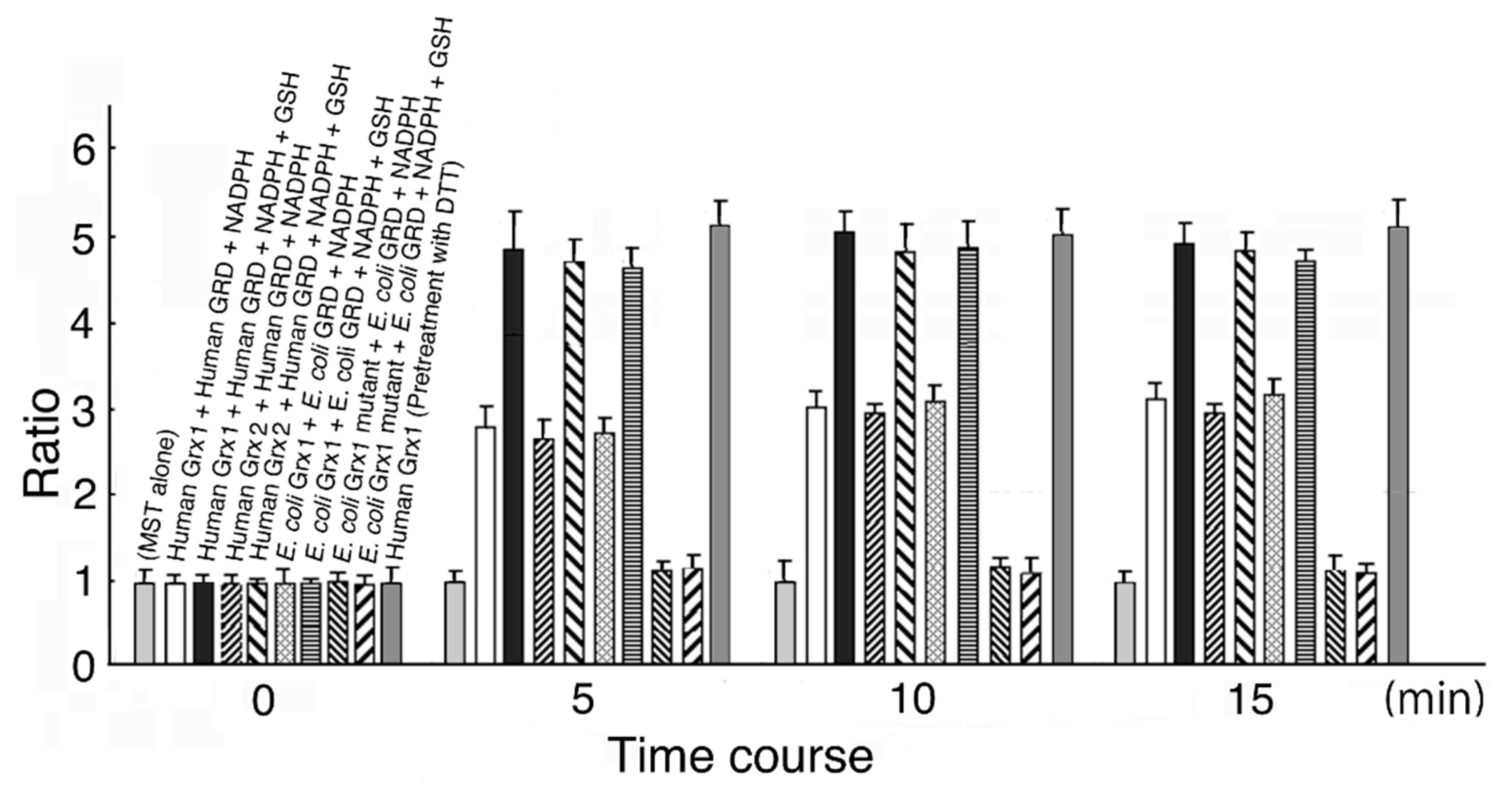
MST activation was completed at around 5 min, and therefore, we measured activity at 10 min in this experiment. As shown in Figure 1, MST activity was increased by 3.01-fold in the reducing system containing human Grx1 (final concentration 0.6 μM), human GRD (final concentration 90 μM), and NADPH (final concentration 8 μM); by 4.13-fold in the system containing human Grx1, human GRD, NADPH, and GSH (final concentration 0.5 mM); by 2.95-fold in the system containing human Grx2 (final concentration 0.6 μM), human GRD, and NADPH; by 3.92-fold in the system containing Grx2, human GRD, NADPH, and GSH; by 3.02-fold in the system containing E. coli Grx1 (final concentration 1.5 μM) (p = 1.69 × 10−1), E. coli GRD, and NADPH; and by 1.10-fold in the system containing E. coli Grx1 mutant, E. coli GRD, NADPH, and GSH (p = 3.52 × 10−1). When MST was treated with 1.2 M DTT, MST activation increased to about 1.5-fold of control, consistent with the previous results [44] (data not shown). Further, when human Grx 1 was pretreated with 1.2 mM DTT, MST activity increased to about 5-fold of control, also consistent with the previous results [44]. Even when the reducing system did not contain GSH, MST activity was increased. This is probably because residual Grx in a reduced form donates electrons to MST until depleted. When GSH was present in the system, activity increased to between 1.58- and 1.70-fold of the values without GSH.
In our previous studies [44,45,46,47], MST was reversely inhibited by a stoichiometric concentration of hydrogen peroxide or oxygen. MST is oxidized to form a sulfenate at the catalytic site cysteine. Furthermore, MST was oxidized to form sulfinate and/or sulfonate to be inactivated. Conversely, MST contained a small amount of these pieces under the present experimental conditions. On the other hand, dimer–monomer equilibrium is also involved in MST activity regulation (in overexpressed MST under air-saturated conditions; monomeric MST to dimeric MST = ~10:1 [44]). The observed MST activation may be due to not only the reduction of a sulfenic acid formed at the catalytic site cysteine but also the reduction of a disulfide bond in dimeric MST [45,46,47]. Grxs may contribute to the dimer–monomer equilibrium of MST. Previous studies confirmed that thioredoxin (Trx) also activates MST via not only the reduction of a sulfenic acid formed at the catalytic site cysteine but also the reduction of a disulfide bond in dimeric MST [45,46,47]. Interestingly, eukaryotic MST is much more effectively activated by prokaryotic Trx than by the eukaryotic one [44]. Conversely, eukaryotic MST is more effectively activated by eukaryotic Grx than by the prokaryotic one.
The E. coli Grx1 mutant had little effect on MST activity with GSH. MST activity with or without GSH after a 10 min incubation increased to 1.24- or 1.16-fold, respectively from before incubation; however, the difference was not significant (p = 9.21 × 10−2 and p = 8.52 × 10−2, respectively). Bushweller et al. [9] reported that Cys14 on E. coli Grx1 was a binding site for GSH and replacement of the Cys with Ser decreased activity to 38% of full.
Considering the redox potential [45,46,47] of the inactive form, oxidized MST (a low redox potential sulfenate formed at the catalytic site cysteine and/or disulfide bridge in dimeric MST) can be reduced and reactivated through accepting electrons from the Trx-TRD system, but not from the GHS-GRD system, because the redox potential of MST is lower than that of GSH (~−240 mV) [48] and higher than that of Trx (~−270 mV) [48]. The findings of this experiment show that the Grx-GSH-GRD system can also reduce MST, despite Grx possessing a higher redox potential (~−200~−233 mV) [48,49]. This is probably because MST and Grx are cross-compatible in their substrate–enzyme interaction.
3.2. Human Grx1 and GSH Dose-Dependent MST Activation
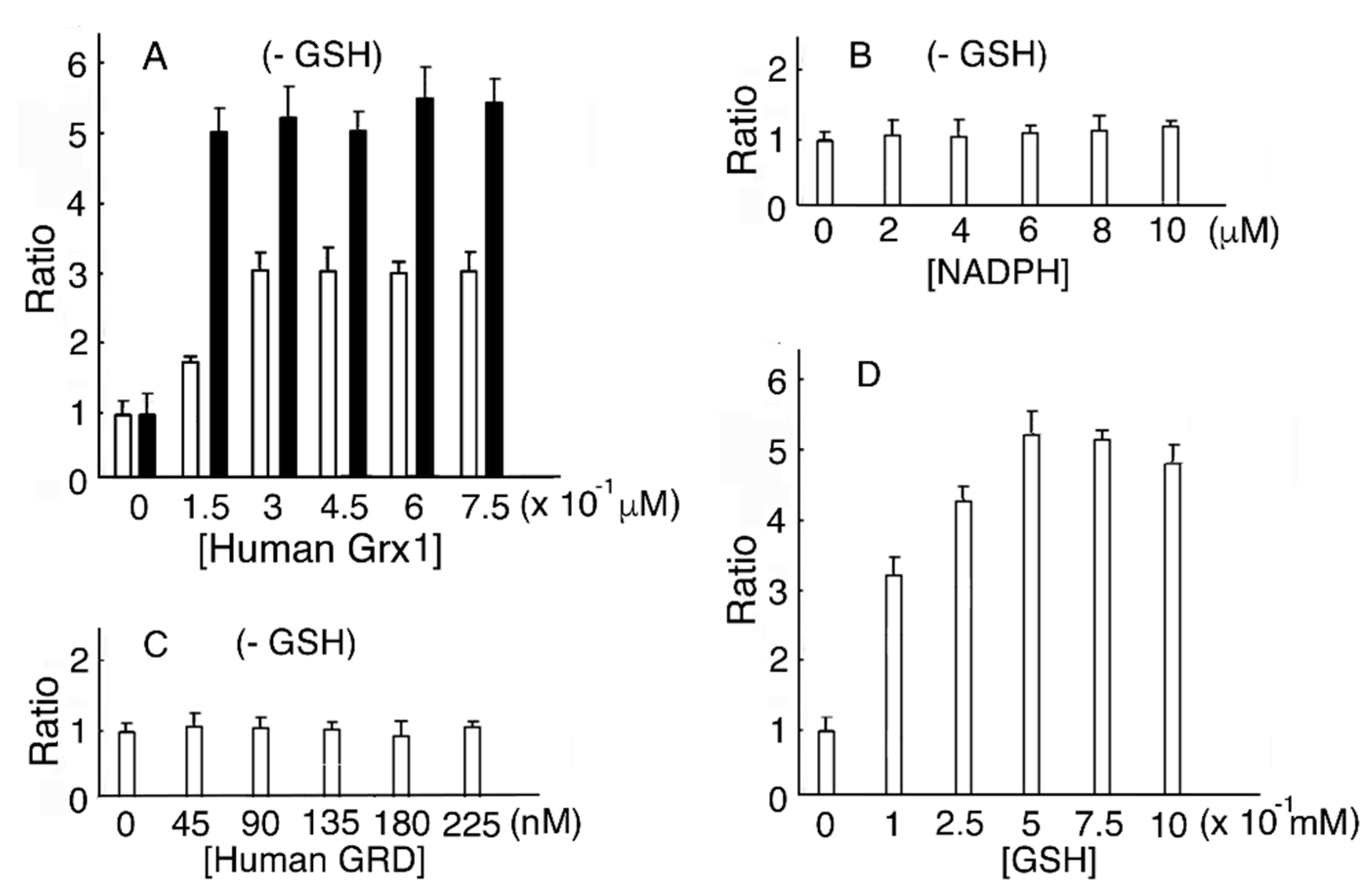
In a reducing system containing human Grx1, human GRD, and NADPH without GSH, MST activity reached a maximum value (2.91-fold of that without human Grx1) at 0.45 μM human Grx1. As a control, MST was incubated with DTT-pretreated human Grx1. The ratio of MST activity showed a maximum value (5.2) (Figure 2A). Residual reduced human Grx1 donates electrons to MST until depleted. When the reducing mixture contained human Grx1, human GRD, and NADPH without GSH, various concentrations of NADPH or human GRD induced no peak in MST activity (Figure 2B,C). It is reasonable to conclude that the absence of Grx does not change MST activity. When various concentrations of GSH were present in the mixture, MST activity reached a maximum (5.32-fold of without GSH) at 0.5 mM GSH (Figure 2D). Our results showed that MST activity at 0.45 μM human Grx1 and 0.5 mM GSH increased to 1.83-fold of 0.45 μM human Grx1 without GSH (p = 6.15 × 10−3).
3.3. Human Grx2 and GSH Dose-Dependent MST Activation
In the reducing system containing human Grx2, human GRD, and NADPH with or without GSH, MST activity reached a maximum value (4.93- and 3.06-fold of that without human Grx2, respectively) at 0.45 μM human Grx2. As a control, MST was incubated with DTT-pretreated human Grx2. The ratio of MST activity showed a maximum value (5.5) (Figure 3). Our results show that MST activity at 0.45 μM human Grx2 and 0.5 mM GSH increased to 1.61-fold of 0.45 μM human Grx2 without GSH. (p = 9.47 × 10−3). When the reducing mixture did not contain GSH, residual reduced human Grx2 donated electrons to MST until depletion. Compared to human Grx1, Grx2 exhibited no difference in function from its counterpart.
3.4. E. coli Grx1 and GSH Dose-Dependent MST Activation
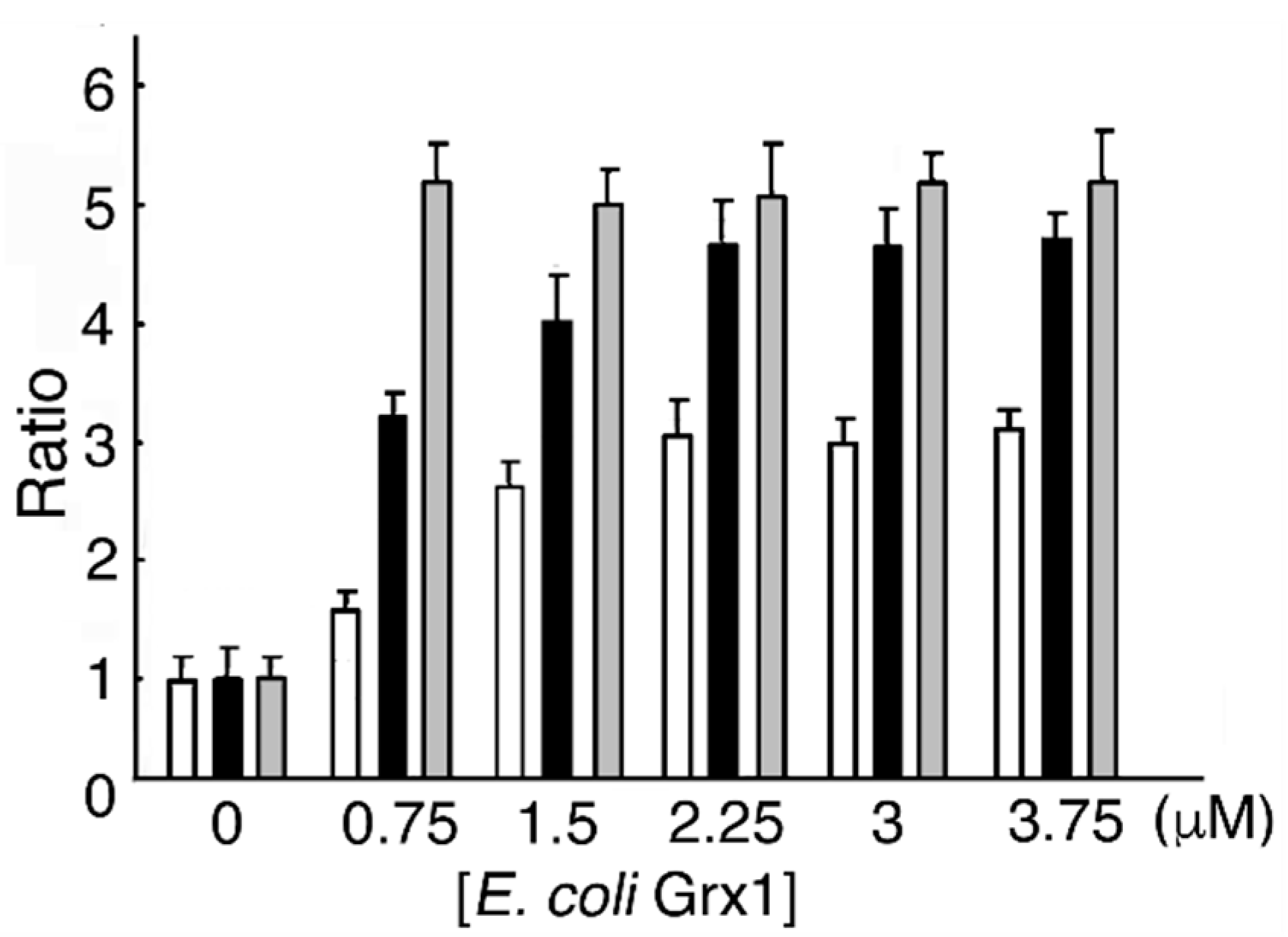
In a reducing system containing E. coli Grx1, E. coli GRD, and NADPH with or without GSH, MST activity reached a maximum value (4.78- and 3.03-fold of that without E. coli Grx1, respectively) at 2.25 μM E. coli Grx1. As a control, MST was incubated DTT-pretreated E. coli Grx1. The ratio of MST activity showed a maximum value (5.4) (Figure 4). Our results show that MST activity at 2.25 μM E. coli Grx1 and 0.5 mM GSH increased to 1.56-fold of that at 2.25 μM E. coli Grx1 without GSH (p = 7.47 × 10−2). When the reducing mixture did not contain GSH, residual reduced E. coli Grx1 donated electrons to MST until depletion. Compared to eukaryotic Grxs, about 5 times the amount of E. coli Grx1 is required to serve as an equivalent electron donor for eukaryotic MST. This experiment suggests that prokaryotic Grx is less effective on eukaryotic proteins than the eukaryotic form. This finding is opposite to the results for prokaryotic Trx, which more effectively reduces eukaryotic proteins than does eukaryotic Trx; this difference is probably due to structural properties [44,45,46,47].
3.5. E. coli Grx1 Mutant and GSH Dose-Dependent MST Activation
In the reducing system containing E. coli Grx1 mutant, E. coli GRD, and NADPH with or without GSH, MST activity was not affected. As a control, MST was incubated with DTT-pretreated E. coli Grx1 mutant. The ratio of MST activity showed a maximum value (1.5) (Figure 5). As discussed above, mutant Grx C14S can maintain 38% of enzyme activity to donate electrons to MST [9], but it hardly works as an electron donor.
4. Conclusions
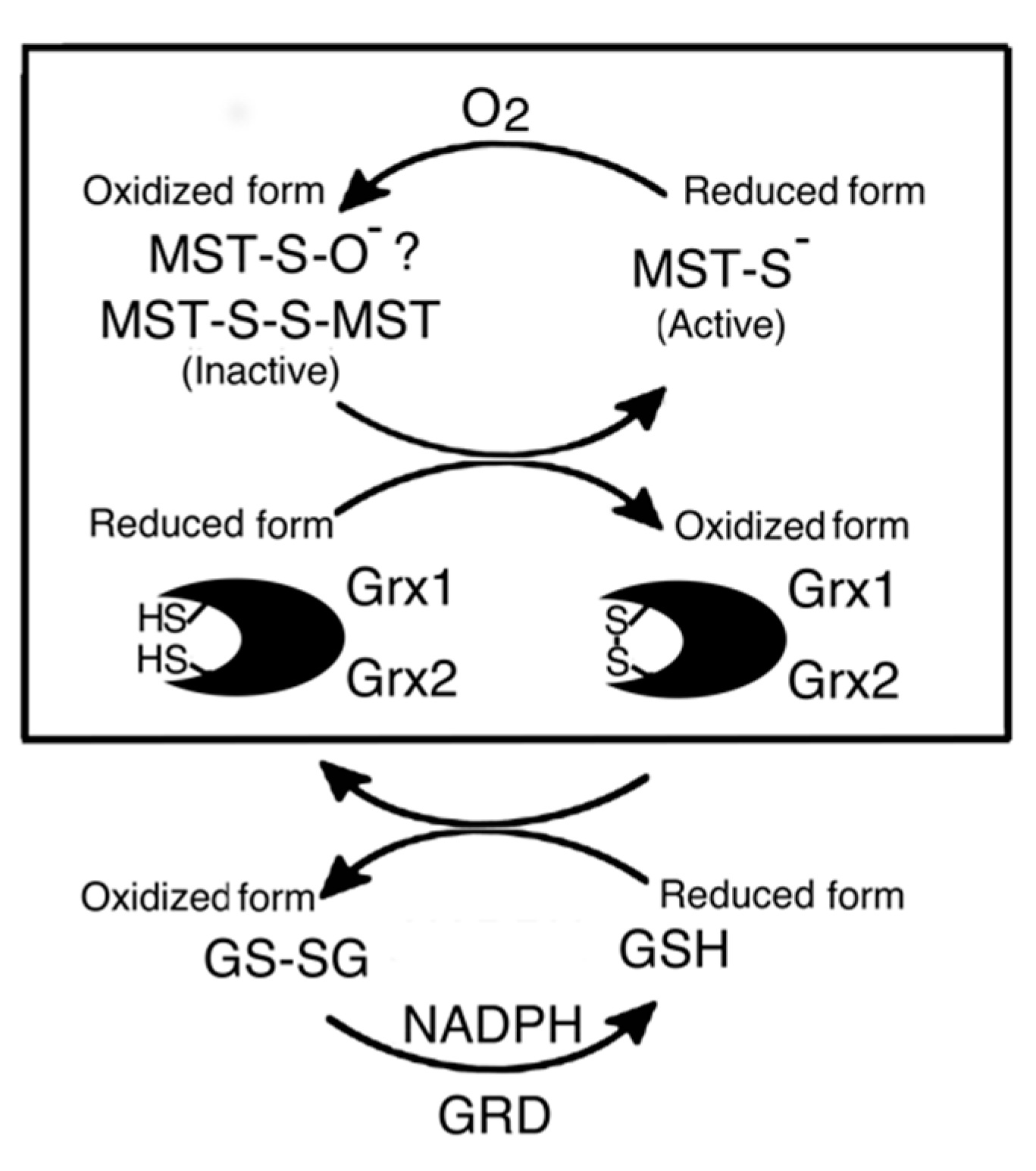
MST is activated in the reducing system containing Grx, GRD, GSH, and NADPH by the reduction of a disulfide bond of dimer MST (Figure 6). The result of a preliminary study on the activation mechanism of MST by human Grx1 mixed with the reducing system (Figure 7) also supported the conclusion. Although Grx possesses higher redox potential than does MST or GSH, Grx enzymatically reduces the oxidized form of MST. This is like the case of thioredoxin 1 that can be reduced by Grx with the reducing mixture [52].
Funding
This research received no external funding.
Conflicts of Interest
The author has no conflicts of interest to declare.
References
- Gleason, F.K.; Holmgre, A. Thioredoxin and related proteins in procaryotes. FEMS Microbiol. Rev. 1988, 4, 271–297. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin and Glutaredoxin: Small multi-functional redox proteins with active-site disulphide bonds. Biochem. Soc. Trans. 1988, 16, 95–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar]
- Yang, Y.; Jao, S.; Nanduri, S.; Starke, D.W.; Mieyal, J.J.; Qin, J. Reactivity of the human thioltransferase (glutaredoxin) C7S, C25S, C78S, C82S mutant and NMR solution structure of its glutathionyl mixed disulfide intermediate reflect catalytic specificity. Biochemistry 1998, 37, 17145–17156. [Google Scholar] [CrossRef]
- Berardi, M.J.; Bushweller, J.H. Binding specificity and mechanistic insight into glutaredoxin-catalyzed protein disulfide reduction. J. Mol. Biol. 1999, 292, 151–161. [Google Scholar] [CrossRef]
- Shelton, M.D.; Chock, P.B.; Mieyal, J.J. Glutaredoxin: Role in reversible protein s-glutathionylation and regulation of redox signal transduction and protein translocation. Antioxid. Redox. Signal. 2005, 7, 348–366. [Google Scholar] [CrossRef]
- Xiao, R.; Lundström-Ljung, J.; Holmgren, A.; Gilbert, H.F. Catalysis of thiol/disulfide exchange. Glutaredoxin 1 and protein-disulfide isomerase use different mechanisms to enhance oxidase and reductase activities. J. Biol. Chem. 2005, 280, 21099–21106. [Google Scholar] [CrossRef] [Green Version]
- Foloppe, F.; Nilsson, L. The Glutaredoxin -C-P-Y-C-motif: Influence of peripheral residues. Structure 2004, 12, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Bushweller, J.H.; Aaslund, F.; Wuethrich, K.; Holmgren, A. Structural and functional characterization of the mutant Escherichia coli glutaredoxin (C14→S) and its mixed disulfide with glutathione. Biochemistry 1992, 31, 9288–9293. [Google Scholar] [CrossRef]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid. Redox. Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef]
- Shibuya, N.; Mikami, Y.; Kimura, Y.; Nagahara, N.; Kimura, H. Vascular endothelium expresses 3-mercaptopyruvate sulfurtransferase and produces hydrogensulfide. J. Biochem. 2009, 146, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, N.; Koike, S.; Tanaka, M.; Ishigami-Yuasa, M.; Kimura, Y.; Ogasawara, Y.; Fukui, K.; Nagahara, N.; Kimura, H. A novel pathway for the production of hydrogen sulfide from D-cysteine in mammalian cells. Nat. Commun. 2013, 4, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, Y.; Shibuya, N.; Kimura, Y.; Nagahara, N.; Ogasawara, Y.; Kimura, H. Thioredoxin and dihydrolipoic acid are required for 3-mercaptopyruvate sulfurtransferase to produce hydrogen sulfide. Biochem. J. 2011, 439, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, Y.; Shibuya, N.; Kimura, Y.; Nagahara, N.; Yamada, M.; Kimura, H. Hydrogen sulfide protects the retina from light-induced degeneration by the modulation of Ca21 influx. J. Biol. Chem. 2011, 286, 39379–39386. [Google Scholar] [CrossRef] [Green Version]
- Modis, K.; Asimakopoulou, A.; Colettal, C.; Papapetropoulos, A.; Szabo, C. Oxidative stress suppresses the cellular bioenergetic effect of the 3-mercaptopyruvate sulfurtransferase/hydrogen sulfide pathway. Biochem. Biophys. Res. Commun. 2013, 433, 401–407. [Google Scholar] [CrossRef]
- Yadav, P.K.; Yamada, K.; Chiku, T.; Koutmos, M.; Banerjee, R. Structure and kinetic analysis of H2S production by human mercaptopyruvate sulfurtransferase. J. Biol. Chem. 2013, 288, 20002–20013. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Toyofuku, Y.; Koike, S.; Shibuya, N.; Nagahara, N.; Lefer, D.; Ogasawara, Y.; Kimura, H. Identification of H2S3 and H2S produced by 3-mercaptopyruvate sulfurtransferase in the brain. Sci. Rep. 2015, 5, 14774. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Koike, S.; Nirasawa, T.; Kimura, H.; Ogasawara, Y. Alternative pathway of H2S and polysulfides production from sulfurated catalytic-cysteine of reaction intermediates of 3-mercaptopyruvate sulfurtransferase. Biochem. Biophys. Res. Commun. 2018, 496, 648–653. [Google Scholar] [CrossRef]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Kimura, H. Physiological roles of hydrogen sulfide and polysulfides. Folia Pharm. Jpn. 2016, 147, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Kimura, H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004, 8, 1165–1167. [Google Scholar] [CrossRef]
- Yang, W.; Yang, G.; Jia, X.; Wu, L.; Wang, R. Activation of KATP channels by H2S in rat insulin-secreting cells and the underlying mechanisms. J. Physiol. 2005, 569, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Kimura, Y.; Kimura, H.; Niki, I. L-Cysteine inhibits insulin release from the pancreatic β-cell. Diabetes 2006, 55, 1391–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, K.R.; Dombkowski, R.A.; Russell, M.J.; Doellman, M.M.; Head, S.K.; Whitfield, N.L.; Madden, J.A. Hydrogen sulfide as an oxygen sensor/transducer in vertebrate hypoxic vasoconstriction and hypoxic vasodilation. J. Exp. Biol. 2006, 209, 4011–4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.J.; Nanduri, J.; Raghuraman, G.; Souvannakitti, D.; Gadalla, M.M.; Kumar, G.K.; Snyder, S.H.; Prabhakar, N.R. H2S mediates O2 sensing in the carotid body. Proc. Natl. Acad. Sci. USA 2010, 107, 10719–10724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanardo, R.C.O.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef]
- Elrod, J.W.; Calvert, J.W.; Morrison, J.; Doeller, J.E.; Kraus, D.W.; Tao, L.; Jiao, X.; Scalia, R.; Kiss, L.; Szabo, C.; et al. Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial function. Proc. Natl. Acad. Sci. USA 2007, 104, 15560–15565. [Google Scholar] [CrossRef] [Green Version]
- Tripatara, P.; Patel, N.; Collino, M.; Gallicchio, M.; Kieswich, J.; Castiglia, S.; Benetti, E.; Stewart, K., N.; Brown, P.A.; Yaqoob, M.M.; et al. Generation of endogenous hydrogen sulfide by cystathionine γ-lyase limits renal ischemia/reperfusion injury and dysfunction. Lab. Invest. 2008, 88, 1038–1048. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.J.; Wang, M.J.; Moore, P.K.; Jin, H.M.; Yao, T.; Zhu, Y.C. The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Pyriochou, A.; Altaany, Z.; Yang, G.; Marazioti, A.; Zhou, Z.; Jeschke, M.G.; Branski, L.K.; Herndon, D.N.; Wang, R.; et al. Hydrogen sulfide is an endogenous stimulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21972–21977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, N.; Fu, C.; Pappin, D.J.; Tonks, N.K. H2S-induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal 2011, 4, ra86. [Google Scholar] [CrossRef] [Green Version]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A universal defense against antibiotics in bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef]
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef]
- Kimura, H. Hydrogen sulfide and polysulfides as signaling molecules. Proc. Jpn. Acad. Ser. B 2015, 91, 131–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, Y.; Tsugane, M.; Oka, J.I.; Kimura, H. Hydrogen sulfide induces calcium waves in astrocytes. FASEB J. 2004, 18, 557–559. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Mikami, Y.; Osumi, K.; Tsugane, M.; Oka, J.; Kimura, H. Polysulfides are possible H2S-derived signaling molecules in rat brain. FASEB J. 2013, 27, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides link H2S to protein thiol oxidation. Antioxid. Redox. Signal 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [Green Version]
- Koike, S.; Ogasawara, Y.; Shibuya, N.; Kimura, H.; Ishii, K. Polysulfide exerts a protective effect against cytotoxicity caused by t-buthylhydroperoxide through Nrf2 signaling in neuroblastoma cells. FEBS Lett. 2013, 587, 3548–3555. [Google Scholar] [CrossRef] [Green Version]
- Koike, S.; Shibuya, N.; Kimura, H.; Ishii, K.; Ogasawara, Y. Polysulfide promotes neuroblastoma cell differentiation by accelerating calcium influx. Biochem. Biophys. Res. Commun. 2015, 459, 488–492. [Google Scholar] [CrossRef]
- Koike, S.; Kayama, T.; Yamamoto, S.; Komine, D.; Tanaka, R.; Nishimoto, S.; Suzuki, T.; Kishida, A.; Ogasawara, Y. Polysulfides protect SH-SY5Y cells from methylglyoxal-induced toxicity by suppressing protein carbonylation: A possible physiological scavenger for carbonyl stress in the brain. NeuroToxicology 2016, 55, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Peleli, M.; Bibli, S.I.; Li, Z.; Chatzianastasiou, A.; Varela, A.; Katsouda, A.; Zukunft, S.; Bucci, M.; Vellecco, V.; Davos, C.H.; et al. Cardiovascular phenotype of mice lacking 3-mercaptopyruvate sulfurtransferase. Biochem. Pharm. 2020. (In print) [Google Scholar] [CrossRef] [PubMed]
- Nasi, S.; Ehirchiou, D.; Chatzianastasiou, A.; Nagahara, N.; Papapetropoulos, A.; Bertrand, J.; Cirino, G.; So, A.; Busso, B. The protective role of the 3-mercaptopyruvate sulfurtransferase (3-MST)-hydrogen sulfide (H2S) pathway against experimental osteoarthritis. Arthritis Res. 2020. (In press) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahara, N.; Yoshii, T.; Abe, Y.; Matsumura, T. Thioredoxin-dependent enzymatic activation of mercaptopyruvate sulfurtransferase: An intersubunit disulfide bond serves as a redox switch for activation. J. Biol. Chem. 2007, 282, 1561–1569. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Katayama, A. Post-translational regulation of mercaptopyruvate sulfurtransferase via a low redox potential cysteine-sulfenate in the maintenance of redox homeostasis. J. Biol. Chem. 2005, 280, 34569–34576. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Nagano, M.; Ito, T.; Suzuki, H. Redox regulation of Mammalian 3-mercaptopyruvate sulfurtransferase. Method. Enzym. 2015, 554, 229–254. [Google Scholar]
- Nagahara, N. Multiple role of 3-mercaptopyruvate sulfurtransferase: Antioxidative function, H2S and polysulfide production, and possible SOx production. Br. J. Pharm. 2018, 175, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Sagemark, J.; Elgán, T.H.; Bürglin, T.R.; Johansson, C.; Holmgren, A.; Berndt, K.D. Redox properties and evolution of human glutaredoxins. Proteins 2007, 68, 879–892. [Google Scholar] [CrossRef]
- Aslund, F.; Berndt, K.D.; Holmgren, A. Redox potentials of glutaredoxins and other thiol-disulfide oxidoreductases of the thioredoxin superfamily determined by direct protein-protein redox equilibria. J. Biol. Chem. 1997, 272, 30780–30786. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Nishino, T. Role of amino acid residues in the active site of rat liver mercaptopyruvate sulfurtransferase. cDNA cloning overexpression, and site-directed mutagenesis. J. Biol. Chem. 1996, 271, 27395–27401. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Okazaki, T.; Nishino, T. Cytosolic mercaptopyruvate sulfurtransferase is evolutionarily related to mitochondrial rhodanese. Striking similarity in active site amino acid sequence and the increase in the mercaptopyruvate sulfurtransferase activity of rhodanese by site-directed mutagenesis. J. Biol. Chem. 1995, 270, 16230–16235. [Google Scholar] [PubMed] [Green Version]
- Du, Y.; Zhang, H.; Lu, J.; Holmgren, A. Glutathione and glutaredoxin act as a backup of human thioredoxin reductase 1 to reduce thioredoxin 1 preventing cell death by aurothioglucose. J. Biol. Chem. 2012, 287, 38210–38219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Time-dependent MST activation. The MST reducing system contains Grx, GSH, GRD, and NADPH. Each system was incubated at 20 °C for 0, 5, 10, and 15 min. Details are described in the text.
Figure 1.
Time-dependent MST activation. The MST reducing system contains Grx, GSH, GRD, and NADPH. Each system was incubated at 20 °C for 0, 5, 10, and 15 min. Details are described in the text.

Figure 2.
Human Grx1 and GSH dose-dependent MST activation. (A). The MST reducing system contains various concentrations of human Grx1, human GRD, and NADPH without GSH. (B). The MST reducing system contains human Grx1, human GRD, and various concentrations of NADPH without GSH. (C). The MST reducing system contains human Grx1, various concentrations of human GRD, and NADPH without GSH. (D). The MST reducing system contains human Grx1, human GRD, and NADPH with various concentrations of GSH. ![Biomolecules 10 00826 i001]() Incubation with DTT-pretreated human Grx1.
Incubation with DTT-pretreated human Grx1.
 Incubation with DTT-pretreated human Grx1.
Incubation with DTT-pretreated human Grx1.
Figure 2.
Human Grx1 and GSH dose-dependent MST activation. (A). The MST reducing system contains various concentrations of human Grx1, human GRD, and NADPH without GSH. (B). The MST reducing system contains human Grx1, human GRD, and various concentrations of NADPH without GSH. (C). The MST reducing system contains human Grx1, various concentrations of human GRD, and NADPH without GSH. (D). The MST reducing system contains human Grx1, human GRD, and NADPH with various concentrations of GSH. ![Biomolecules 10 00826 i001]() Incubation with DTT-pretreated human Grx1.
Incubation with DTT-pretreated human Grx1.
Incubation with DTT-pretreated human Grx1.
Figure 3.
Human Grx2 and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of human Grx2, human GRD, NADPH, and GSH, while the other reducing system contains human Grx2, human GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i002]() ) or without (
) or without ( ![Biomolecules 10 00826 i003]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i004]() ), Incubation with DTT-pretreated human Grx2.
), Incubation with DTT-pretreated human Grx2.
 ) or without (
) or without (  ) GSH. (
) GSH. (  ), Incubation with DTT-pretreated human Grx2.
), Incubation with DTT-pretreated human Grx2.
Figure 3.
Human Grx2 and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of human Grx2, human GRD, NADPH, and GSH, while the other reducing system contains human Grx2, human GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i002]() ) or without (
) or without ( ![Biomolecules 10 00826 i003]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i004]() ), Incubation with DTT-pretreated human Grx2.
), Incubation with DTT-pretreated human Grx2.
) or without ( ) GSH. ( ), Incubation with DTT-pretreated human Grx2.
Figure 4.
Escherichia coli Grx1 and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of E. coli Grx1, E. coli GRD, NADPH, and GSH, while the other reducing system contains E. coli Grx1, E. coli GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i005]() ) or without (
) or without ( ![Biomolecules 10 00826 i006]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i007]() ) , Incubation with DTT-pretreated E. coli Grx1.
) , Incubation with DTT-pretreated E. coli Grx1.
 ) or without (
) or without (  ) GSH. (
) GSH. (  ) , Incubation with DTT-pretreated E. coli Grx1.
) , Incubation with DTT-pretreated E. coli Grx1.
Figure 4.
Escherichia coli Grx1 and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of E. coli Grx1, E. coli GRD, NADPH, and GSH, while the other reducing system contains E. coli Grx1, E. coli GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i005]() ) or without (
) or without ( ![Biomolecules 10 00826 i006]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i007]() ) , Incubation with DTT-pretreated E. coli Grx1.
) , Incubation with DTT-pretreated E. coli Grx1.
) or without ( ) GSH. ( ) , Incubation with DTT-pretreated E. coli Grx1.
Figure 5.
E. coli Grx1 mutant and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of E. coli Grx1 mutant, E. coli GRD, NADPH, and GSH. The other reducing system contains E. coli Grx1 mutant, E. coli GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i008]() ) or without (
) or without ( ![Biomolecules 10 00826 i009]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i010]() ), Incubation with DTT-pretreated E. coli Grx1 mutant.
), Incubation with DTT-pretreated E. coli Grx1 mutant.
 ) or without (
) or without (  ) GSH. (
) GSH. (  ), Incubation with DTT-pretreated E. coli Grx1 mutant.
), Incubation with DTT-pretreated E. coli Grx1 mutant.
Figure 5.
E. coli Grx1 mutant and GSH dose-dependent MST activation. One MST reducing system contains various concentrations of E. coli Grx1 mutant, E. coli GRD, NADPH, and GSH. The other reducing system contains E. coli Grx1 mutant, E. coli GRD, NADPH, and various concentrations of GSH. Each mixture was incubated at 20 °C for 10 min. The mixture is with ( ![Biomolecules 10 00826 i008]() ) or without (
) or without ( ![Biomolecules 10 00826 i009]() ) GSH. (
) GSH. ( ![Biomolecules 10 00826 i010]() ), Incubation with DTT-pretreated E. coli Grx1 mutant.
), Incubation with DTT-pretreated E. coli Grx1 mutant.
) or without ( ) GSH. ( ), Incubation with DTT-pretreated E. coli Grx1 mutant.
Figure 6.
Probable activation mechanism of MST in the Grx-GSH-GRD-NADPH reducing systems. Box: The reducing system contains Grx without Grx, GRD, and NADPH. Details are described in the text.
Figure 6.
Probable activation mechanism of MST in the Grx-GSH-GRD-NADPH reducing systems. Box: The reducing system contains Grx without Grx, GRD, and NADPH. Details are described in the text.

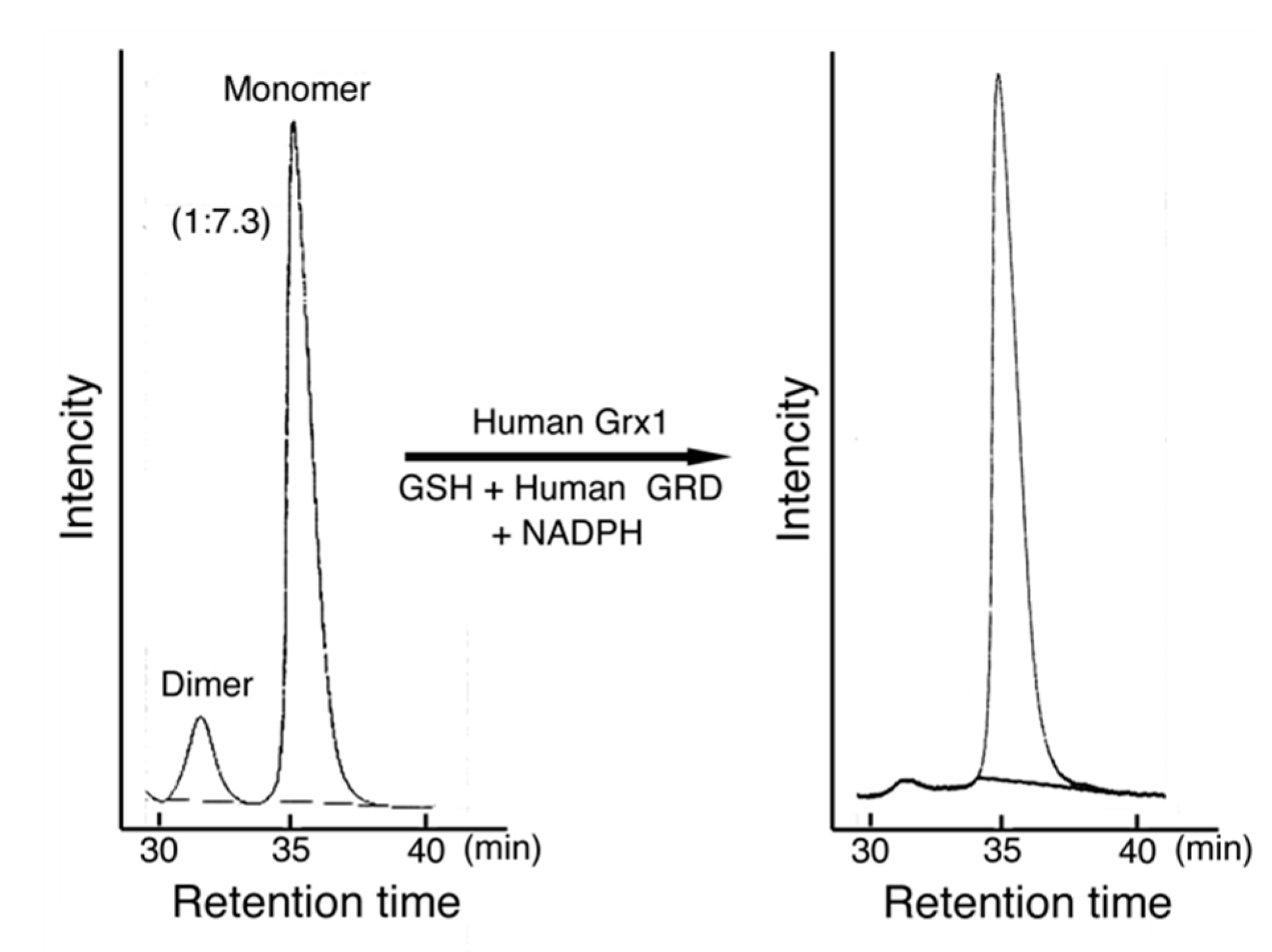
Figure 7.
Preliminary study on the activation mechanism of MST. MST was mixed with a human Grx1 mixture containing GSH, human GRD, and NADPH for 20 min on ice. The mixture and control MST were analyzed using an HPLC system (Hitachi Chromaster system, Tokyo, Japan) with two TSK gel filtration columns connected in series (G3000SWxL, 7.8 mm × 30 cm and G2000SWxL, 7.8 mm × 30 cm, TOSOH Corp., Tokyo, Japan). The mobile phase was 0.2 M potassium phosphate buffer, pH 7.2. The flow rate was 0.5 mL/min. The detection was made at 280 nm. In this case, MST contained 13.6% of the dimer. The dimer MST was reduced to monomer MST and MST activity was increased.
Figure 7.
Preliminary study on the activation mechanism of MST. MST was mixed with a human Grx1 mixture containing GSH, human GRD, and NADPH for 20 min on ice. The mixture and control MST were analyzed using an HPLC system (Hitachi Chromaster system, Tokyo, Japan) with two TSK gel filtration columns connected in series (G3000SWxL, 7.8 mm × 30 cm and G2000SWxL, 7.8 mm × 30 cm, TOSOH Corp., Tokyo, Japan). The mobile phase was 0.2 M potassium phosphate buffer, pH 7.2. The flow rate was 0.5 mL/min. The detection was made at 280 nm. In this case, MST contained 13.6% of the dimer. The dimer MST was reduced to monomer MST and MST activity was increased.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nagahara, N. Activation of 3-Mercaptopyruvate Sulfurtransferase by Glutaredoxin Reducing System. Biomolecules 2020, 10, 826. https://doi.org/10.3390/biom10060826
AMA Style
Nagahara N. Activation of 3-Mercaptopyruvate Sulfurtransferase by Glutaredoxin Reducing System. Biomolecules. 2020; 10(6):826. https://doi.org/10.3390/biom10060826
Chicago/Turabian StyleNagahara, Noriyuki. 2020. "Activation of 3-Mercaptopyruvate Sulfurtransferase by Glutaredoxin Reducing System" Biomolecules 10, no. 6: 826. https://doi.org/10.3390/biom10060826
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.