Effects of pH and Mineral Nutrition on Growth and Physiological Responses of Trembling Aspen (Populus tremuloides), Jack Pine (Pinus banksiana), and White Spruce (Picea glauca) Seedlings in Sand Culture


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
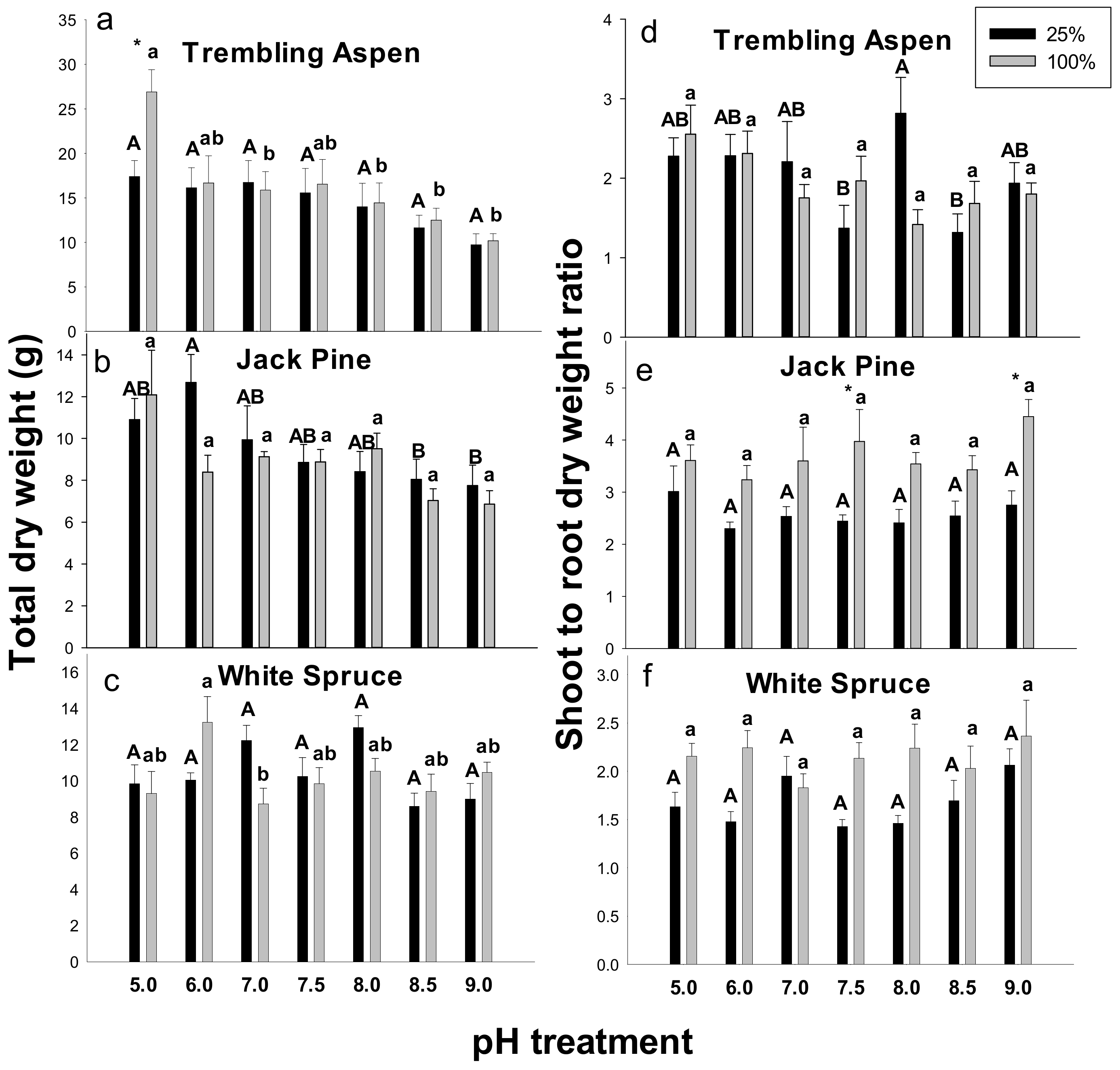
2.1. Total Dry Weights and Shoot to Root (s/r) Dry Weight Ratios
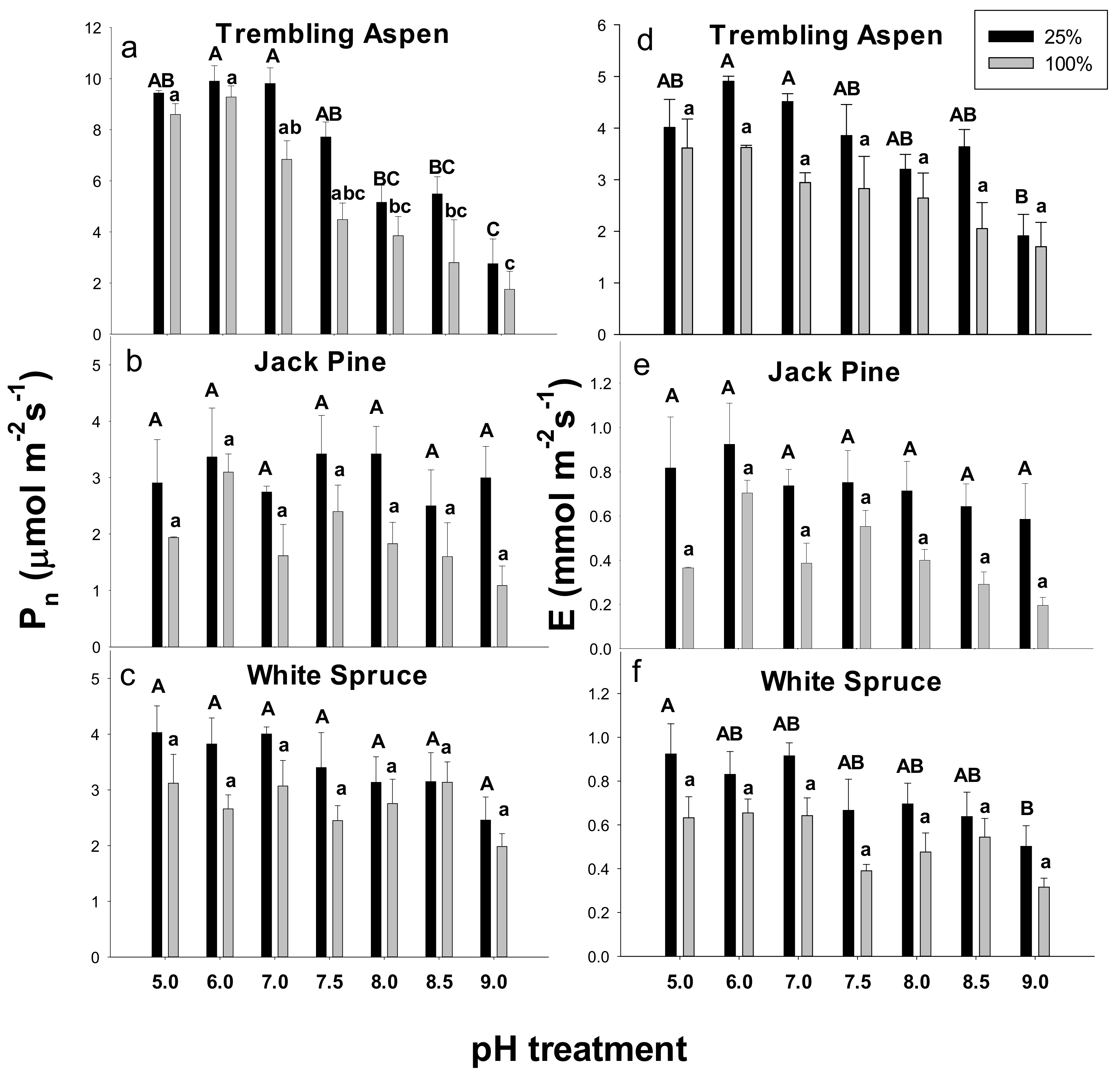
2.2. Gas Exchange
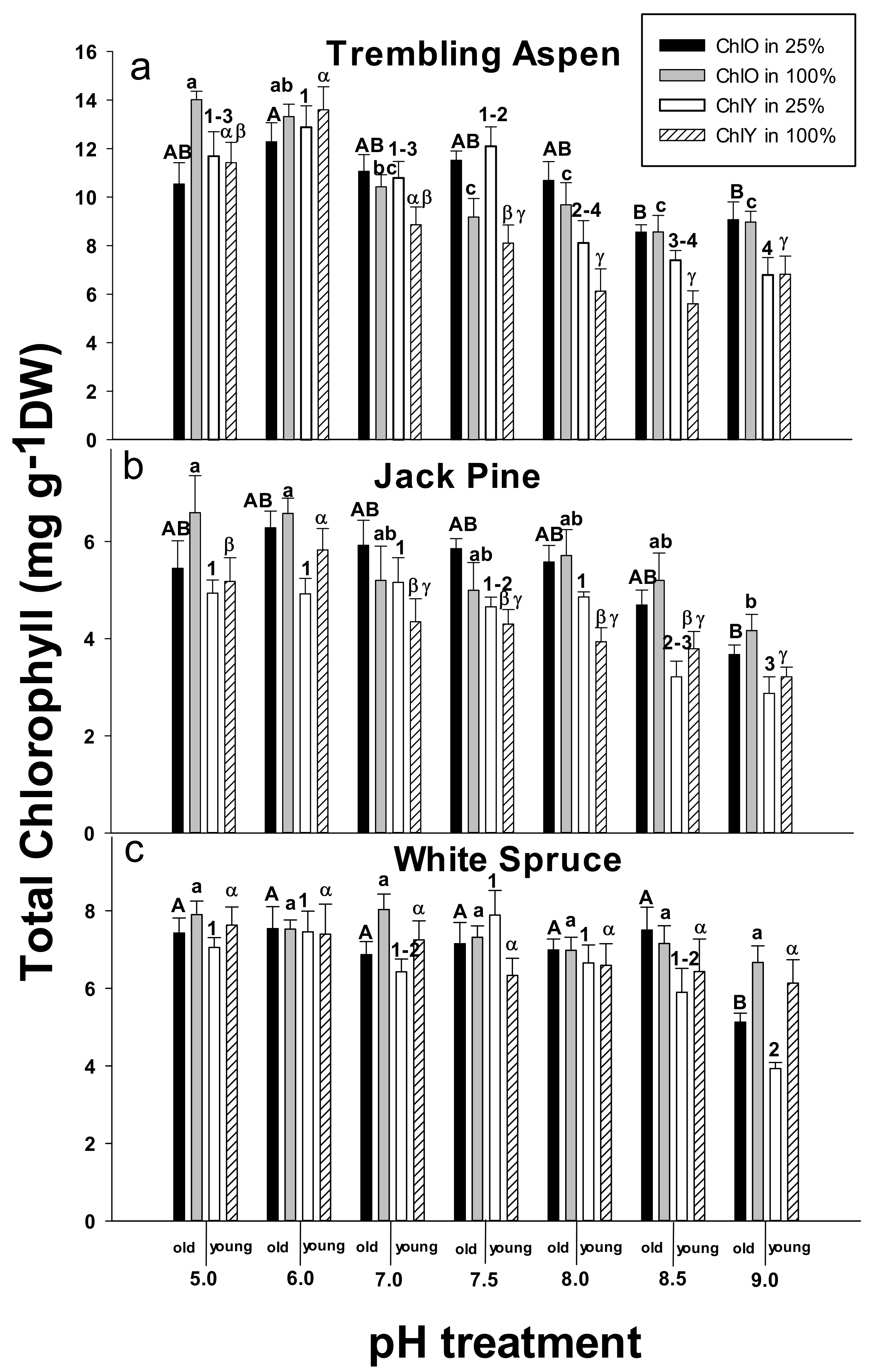
2.3. Chlorophyll Concentrations
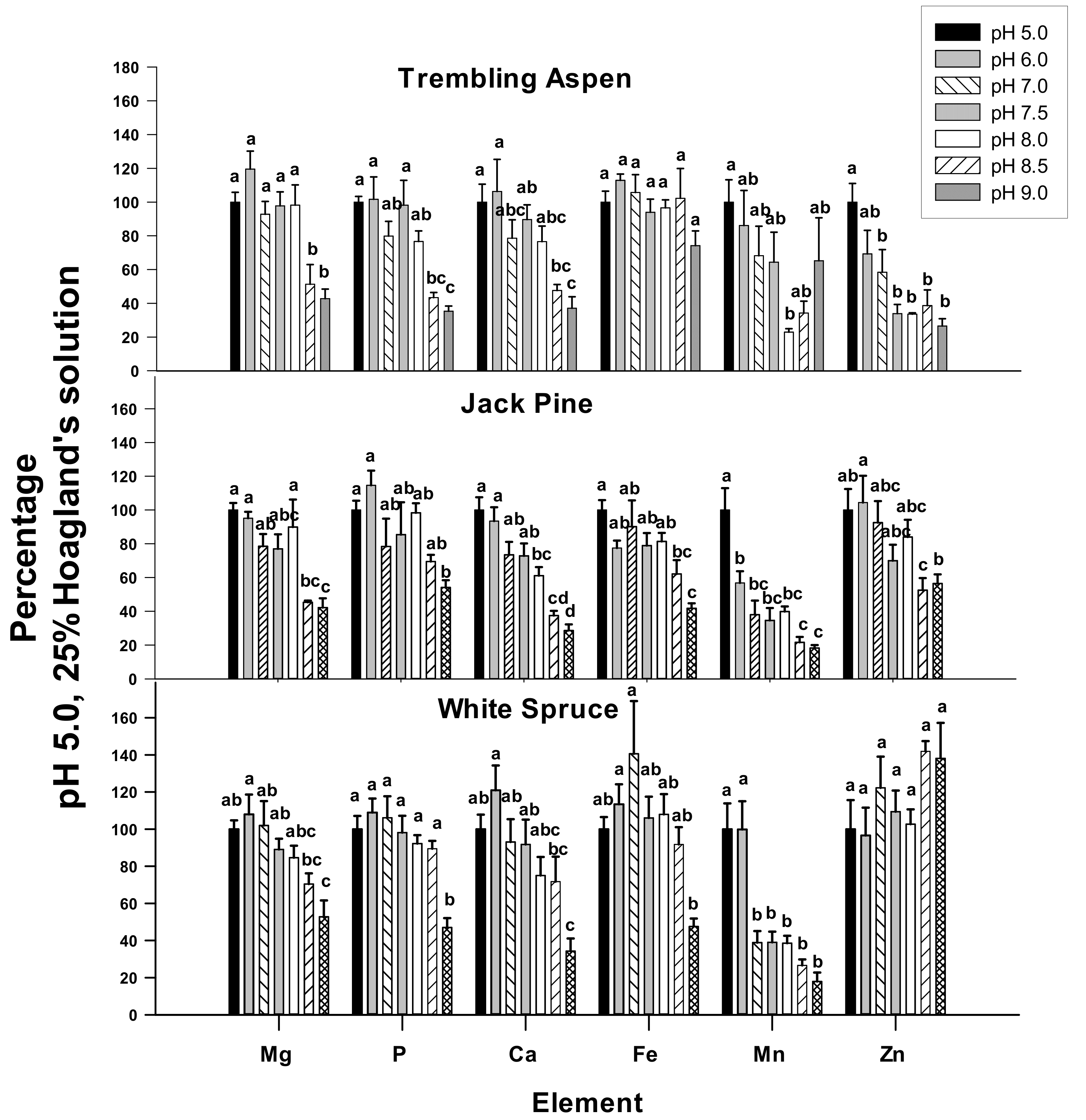
2.4. Elemental Concentrations of Young Leaves in 25% Hoagland’s Solution
3. Discussion
4. Materials and Methods
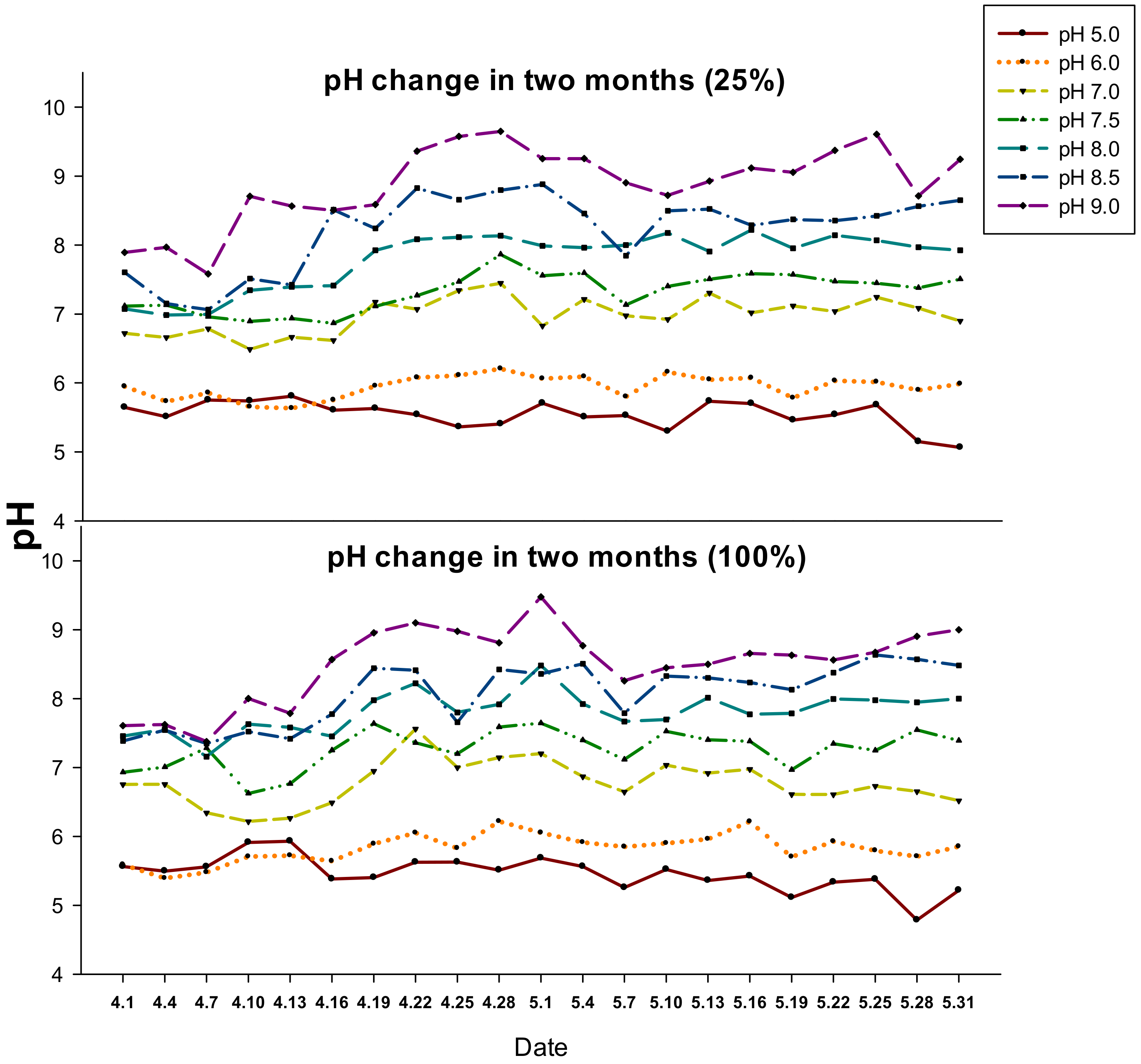
4.1. Plants and Experimental Setup
4.2. Experimental Treatments
4.3. Dry Weights and Foliar Chlorophyll Concentrations
4.4. Net Photosynthesis (Pn) and Transpiration (E) Rates
4.5. Elemental Analysis of Young Foliage
4.6. Experimental Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rengel, Z. Role of pH in availability of ions in soil. In Handbook of Plant Growth pH as the Master Variable; Rengel, Z., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 2002; pp. 323–350. [Google Scholar]
- Chen, Y.; Barak, P. Iron Nutrition of Plants in Calcareous Soils. Adv. Agron. 1982, 35, 217–240. [Google Scholar] [CrossRef]
- Government of Alberta. Oil Sands Overview. 2017. Available online: https://www.alberta.ca/oil-sands-overview.aspx (accessed on 21 November 2019).
- Terrestrial Subgroup Cumulative Environmental Management Association. Guidelines for Reclamation to Forest Vegetation in the Athabasca Oil Sands Region, 2nd ed.; Alberta Environment: Edmonton, AB, Canada, 2010. [Google Scholar]
- Howat, D. Acceptable Salinity, Sodicity and pH Values for Boreal Forest Reclamation; Report # ESD/LM/00-2; Alberta Environment: Edmonton, AB, Canada, 2000. [Google Scholar]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils; Prentice Hall Inc.: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- Zhang, W.; Zwiazek, J.J. Responses of reclamation plants to high root zone pH: Effects of phosphorus and calcium availability. J. Environ. Qual. 2016, 45, 1652–1662. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates Inc.: Sunderland, UK, 2005. [Google Scholar]
- Felle, H.H.; Hanstein, S. The apoplastic pH of the substomatal cavity of Vicia faba leaves and its regulation responding to different stress factors. J. Exp. Bot. 2002, 53, 73–82. [Google Scholar]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Cobley, B.T.; Mokhtara, S.; Wilson, C.E.; Greenway, H. High pH in the nutrient solution impairs water uptake in Lupinus angustifolius L. Plant Soil 1993, 155, 517–519. [Google Scholar] [CrossRef]
- Kamaluddin, M.; Zwiazek, J.J. Effects of root medium pH on water transport in paper birch (Betula papyrifera) seedlings in relation to root temperature and abscisic acid treatments. Tree Physiol. 2004, 24, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Kopittke, P.M.; Menzies, N. Effect of pH on Na induced Ca deficiency. Plant Soil 2005, 269, 119–129. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Zhang, W.-Q.; Macdonald, S.E.; Señorans, J.; Zwiazek, J.J. Boreal forest plant species responses to pH: ecological interpretation and application to reclamation. Plant Soil 2017, 420, 195–208. [Google Scholar] [CrossRef]
- Sheridan, C.; Depuydt, P.; De Ro, M.; Petit, C.; van Gysegem, E.; Delaere, P.; Dixon, M.; Stasiak, M.; Aciksöz, S.B.; Frossard, E.; et al. Microbial community dynamics and response to plant growth-promoting microorganisms in the rhizosphere of four common food crops cultivated in hydroponics. Microb. Ecol. 2016, 73, 378–393. [Google Scholar] [CrossRef]
- Hose, E.; Clarkson, D.; Steudle, E.; Schreiber, L.; Hartung, W. The exodermis: A variable apoplastic barrier. J. Exp. Bot. 2001, 52, 2245–2264. [Google Scholar] [CrossRef]
- Sankhalkar, S.; Komarpant, R.; Dessai, T.R.; Simoes, J.; Sharma, S. Effects of soil and soil-less culture on morphology, physiology and biochemical studies of vegetable plants. Curr. Agric. Res. J. 2019, 7, 181–188. [Google Scholar] [CrossRef]
- Tang, C.; Turner, N.C. The influence of alkalinity and water stress on the stomatal conductance, photosynthetic rate and growth of Lupinus angustifolius L. and Lupinus pilosus Murr. Aust. J. Exp. Agric. 1999, 39, 457. [Google Scholar] [CrossRef]
- Zhang, W.-Q.; Calvo-Polanco, M.; Chen, Z.C.; Zwiazek, J.J. Growth and physiological responses of trembling aspen (Populus tremuloides), white spruce (Picea glauca) and tamarack (Larix laricina) seedlings to root zone pH. Plant Soil 2013, 373, 775–786. [Google Scholar] [CrossRef]
- Zhang, W.-Q.; Xu, F.; Zwiazek, J.J. Responses of jack pine (Pinus banksiana) seedlings to root zone pH and calcium. Environ. Exp. Bot. 2015, 111, 32–41. [Google Scholar] [CrossRef]
- Tang, C.; Longnecker, N.; Thomson, C.J.; Greenway, H.; Robson, A. Lupin (Lupinus angustifolius L.) and pea (Pisum sativum L.) roots differ in their sensitivity to pH above 6.0. J. Plant Physiol. 1992, 140, 715–719. [Google Scholar] [CrossRef]
- Tang, C.; Longnecker, N.; Greenway, H.; Robson, A.D. Reduced root elongation of Lupinus angustifolius L. by high pH is not due to decreased membrane integrity of cortical cells or low proton production by the roots. Ann. Bot. 1996, 78, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-Q.; Zwiazek, J.J. Effects of root medium pH on root water transport and apoplastic pH in red-osier dogwood (Cornus sericea) and paper birch (Betula papyrifera) seedlings. Plant Biol. 2016, 18, 1001–1007. [Google Scholar] [CrossRef]
- Yousfi, S.; Wissal, M.; Mahmoudi, H.; Abdelly, C.; Gharsalli, M. Effect of salt on physiological responses of barley to iron deficiency. Plant Physiol. Biochem. 2007, 45, 309–314. [Google Scholar] [CrossRef]
- Renault, S.; Zwiazek, J.J.; Fung, M.; Tuttle, S. Germination, growth and gas exchange of selected boreal forest seedlings in soil containing oil sands tailings. Environ. Pollut. 2000, 107, 357–365. [Google Scholar] [CrossRef]
- Xu, F.; Tan, X.; Zhang, W.-Q.; Zwiazek, J.J. Effects of iron and root zone pH on growth and physiological responses of paper birch (Betula papyrifera), trembling aspen (Populus tremuloides) and red-osier dogwood (Cornus stolonifera) seedlings in a split-root hydroponic system. Acta Physiol. Plant. 2019, 41, 142. [Google Scholar] [CrossRef]
- Bowman, W.D.; Conant, R.T. Shoot growth dynamics and photosynthetic response to increased nitrogen availability in the alpine willow Salix glauca. Oecologia 1994, 97, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Hemming, J.D.C.; Lindroth, R.L. Effects of light and nutrient availability on aspen: growth, phytochemistry, and insect performance. J. Chem. Ecol. 1999, 25, 1687–1714. [Google Scholar] [CrossRef]
- Yu, Q.; Tang, C.; Kuo, J. Apoplastic pH in roots of Lupinus angustifolius L. in response to pH > 6. In Plant Nutrition; Springer Science and Business Media LLC: New York, NY, USA, 2001; pp. 242–243. [Google Scholar]
- Baszynski, T.; Warchoówa, M.; Krupa, Z.; Tukendorf, A.; Król, M.; Wolinska, D. The effect of magnesium deficiency on photochemical activities of rape and buckwheat chloroplasts. Z. Pflanzenphysiol. 1980, 99, 295–303. [Google Scholar]
- Larbi, A.; Abadía, A.; Morales, F.; Abadía, J. Fe resupply to Fe-deficient sugar beet plants leads to rapid changes in the violaxanthin cycle and other photosynthetic characteristics without significant de novo chlorophyll synthesis. Photosynth. Res. 2004, 79, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Shenker, M.; Plessner, O.E.; Tel-Or, E. Manganese nutrition effects on tomato growth, chlorophyll concentration, and superoxide dismutase activity. J. Plant Physiol. 2004, 161, 197–202. [Google Scholar] [CrossRef]
- Zhang, S.; Dang, Q.-L. Effects of carbon dioxide concentration and nutrition on photosynthetic functions of white birch seedlings. Tree Physiol. 2006, 26, 1457–1467. [Google Scholar] [CrossRef] [Green Version]
- Siemens, J.A.; Zwiazek, J.J. Hebeloma crustuliniforme modifies root hydraulic responses of trembling aspen (Populus tremuloides) seedlings to changes in external pH. Plant Soil 2011, 345, 247–256. [Google Scholar] [CrossRef]
- Tang, C.; Kuo, J.; Longnecker, N.E.; Thomson, C.J.; Robson, A.D. High pH causes disintegration of the root surface in Lupinus angustifolius L. Ann. Bot. 1993, 71, 201–207. [Google Scholar] [CrossRef]
- Barnes, J.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Valentine, D.W.; Kielland, K.; Chapin, F.S., III; McCuire, A.D.; Van Cleve, K. Patterns of biogeochemistry in Alaskan boreal forests. In Alaska’s Changing Boreal Forest; Chapin, F.S., Oswood, M.W., Van Cleve, K., Viereck, L.A., Verbyla, D.L., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 241–265. [Google Scholar]
- Zarcinas, B.; Cartwright, B.; Spouncer, L. Nitric acid digestion and multielement analysis of plant material by inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1987, 18, 131–146. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, F.; Vaziriyeganeh, M.; Zwiazek, J.J. Effects of pH and Mineral Nutrition on Growth and Physiological Responses of Trembling Aspen (Populus tremuloides), Jack Pine (Pinus banksiana), and White Spruce (Picea glauca) Seedlings in Sand Culture. Plants 2020, 9, 682. https://doi.org/10.3390/plants9060682
Xu F, Vaziriyeganeh M, Zwiazek JJ. Effects of pH and Mineral Nutrition on Growth and Physiological Responses of Trembling Aspen (Populus tremuloides), Jack Pine (Pinus banksiana), and White Spruce (Picea glauca) Seedlings in Sand Culture. Plants. 2020; 9(6):682. https://doi.org/10.3390/plants9060682
Chicago/Turabian StyleXu, Feng, Maryamsadat Vaziriyeganeh, and Janusz J. Zwiazek. 2020. "Effects of pH and Mineral Nutrition on Growth and Physiological Responses of Trembling Aspen (Populus tremuloides), Jack Pine (Pinus banksiana), and White Spruce (Picea glauca) Seedlings in Sand Culture" Plants 9, no. 6: 682. https://doi.org/10.3390/plants9060682