Characterization of Scardovia wiggsiae Biofilm by Original Scanning Electron Microscopy Protocol


 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Static Biofilm Assay of S. mutans and S. wiggsiae
2.3. SEM Protocols
3. Results
3.1. S. mutans Prepared by Conventional SEM Procedure
3.2. S. mutans Prepared by Conventional VP-SEM Procedure
3.3. S. mutans Prepared by OsO4-RR-TA-IL Procedure
3.4. Evaluation of Best Suitable Protocol for Biofilm Imaging on S. mutans Samples
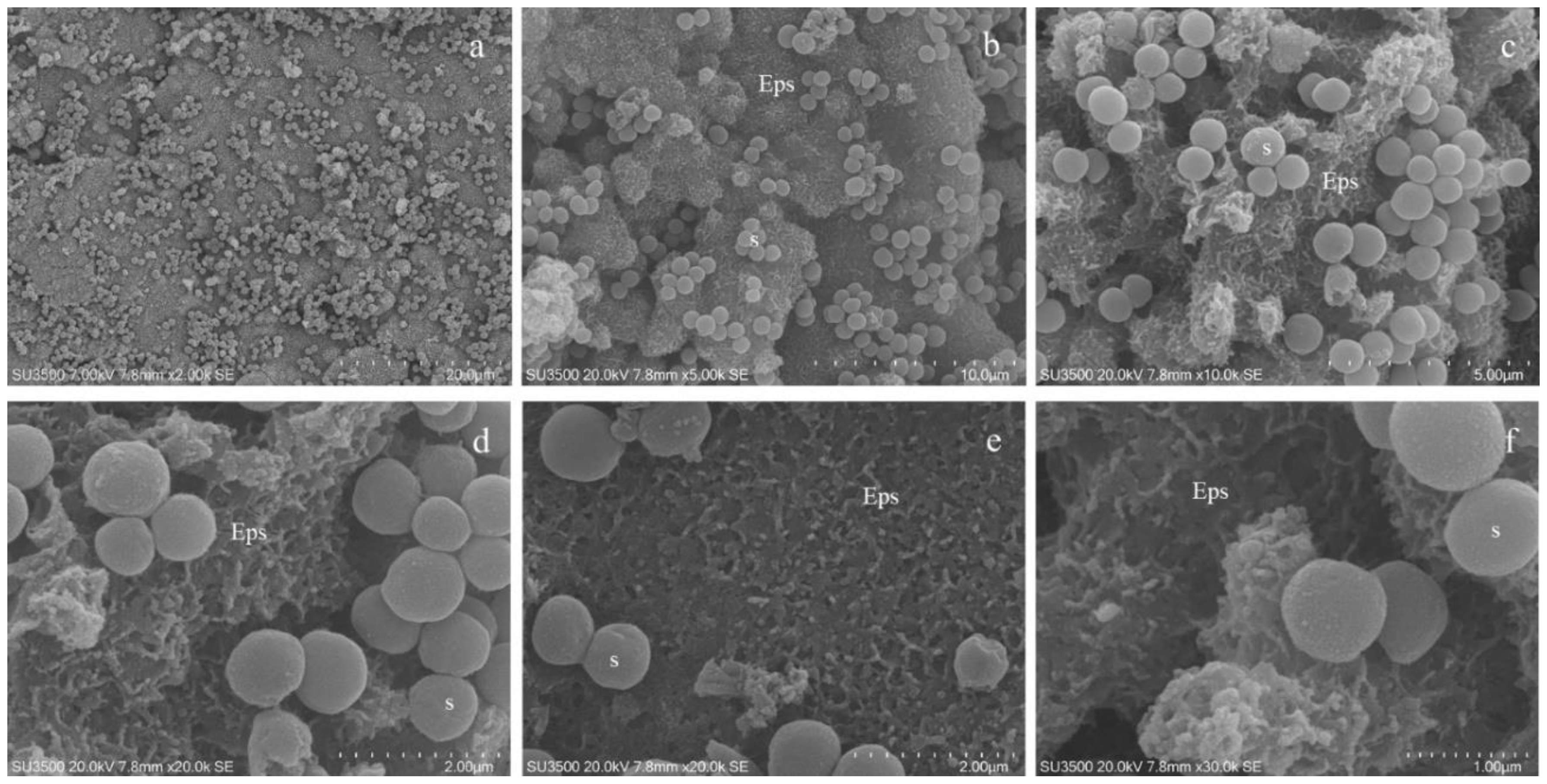
3.5. Characterization of Biofilm Formation by S. wiggsiae DSM 22547
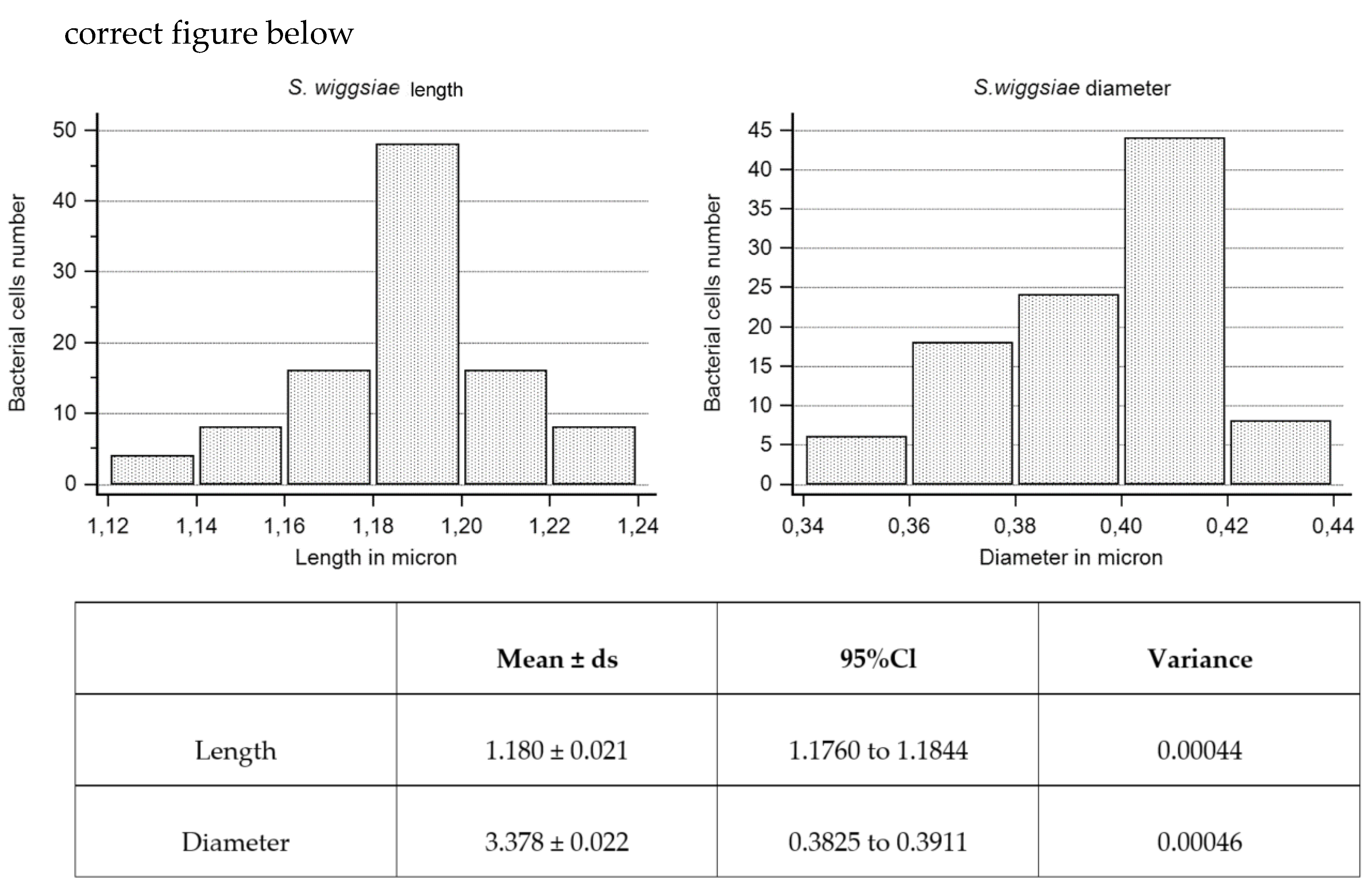
3.6. Ultrastructural Morphological Characterization of S. wiggsiae and Its Biofilm
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berg, J.H.; Slayton, R.L. Early Childhood Oral Health, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Douglass, J.M.; Douglass, A.B.; Silk, H.J. A practical guide to infant oral health. Am. Fam. Physician 2004, 70, 2113–2120. [Google Scholar] [PubMed]
- Martins-Junior, P.A.; Vieira-Andrade, R.G.; Correa-Faria, P.; Oliveira-Ferreira, F.; Marques, L.S.; Ramos-Jorge, M.L. Impact of early childhood caries on the oral health-related quality of life of preschool children and their parents. Caries Res. 2013, 47, 211–218. [Google Scholar] [PubMed]
- Meyer, F.; Enax, J. Early childhood caries: Epidemiology, aetiology, and prevention. Int. J. Dent. 2018, 2018, 1415873. [Google Scholar] [CrossRef] [Green Version]
- Van Houte, J.; Gibbs, G.; Butera, C. Oral flora of children with “nursing bottle caries”. J. Dent. Res. 1982, 61, 382–385. [Google Scholar] [CrossRef]
- Berkowitz, R.J.; Turner, J.; Hughes, C. Microbial characteristics of the human dental caries associated with prolonged bottle-feeding. Arch. Oral Biol. 1984, 29, 949–951. [Google Scholar]
- Parisotto, T.M.; Steiner-Oliveira, C.; Silva, C.M.; Rodrigues, L.K.; Nobre-dos-Santos, M. Early childhood caries and mutans streptococci: A systematic review. Oral Health Prev. Dent. 2010, 8, 59–70. [Google Scholar]
- Peterson, S.N.; Meissner, T.; Su, A.I.; Snesrud, E.; Ong, A.C.; Schork, N.J.; Bretz, W.A. Functional expression of dental plaque microbiota. Front. Cell. Infect. Microbiol. 2014, 4, 108. [Google Scholar]
- Li, Y.; Tanner, A. Effect of antimicrobial interventions on the oral microbiota associated with early childhood caries. Pediatr. Dent. 2015, 37, 226–244. [Google Scholar]
- Becker, M.R.; Paster, B.J.; Leys, E.J.; Moeschberger, M.L.; Kenyon, S.G.; Galvin, J.L.; Boches, S.K.; Dewhirst, F.E.; Griffen, A.L. Molecular analysis of bacterial species associated with childhood caries. J. Clin. Microbiol. 2002, 40, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Van Houte, J.; Lopman, J.; Kent, R. The final pH of bacteria comprising the predominant flora on sound and carious human root and enamel surfaces. J. Dent. Res. 1996, 75, 1008–1014. [Google Scholar] [CrossRef]
- Downes, J.; Mantzourani, M.; Beighton, D.; Hooper, S.; Wilson, M.J.; Nicholson, A.; Wade, W.G. Scardovia wiggsiae sp. nov., isolated from the human oral cavity and clinical material, and emended descriptions of the genus Scardovia and Scardovia inopinata. Int. J. Syst. Evol. Microbiol. 2011, 61, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Tanner, A.C.; Mathney, J.M.; Kent, R.L.; Chalmers, N.I.; Hughes, C.V.; Loo, C.Y.; Pradhan, N.; Kanasi, E.; Hwang, J.; Dahlan, M.A.; et al. Cultivable anaerobic microbiota of severe early childhood caries. J. Clin. Microbiol. 2011, 49, 1464–1474. [Google Scholar] [CrossRef] [Green Version]
- Ledezma-Rasillo, G.; Flores-Reyes, H.; Gonzalez-Amaro, A.M.; Garrocho-Rangel, A.; del Socorro Ruiz-Rodriguez, M.; Pozos-Guillen, A.J. Identification of cultivable microrganisms from primary teeth with necrotic pulps. J. Clin. Pediatr. Dent. 2010, 34, 329–333. [Google Scholar] [CrossRef]
- Tanner, A.C.; Sonis, A.L.; Lif Holgerson, P.; Starr, J.R.; Nunez, Y.; Kressirer, C.A.; Paster, B.J.; Johansson, I. White-spot lesions and gingivitis microbiotas in orthodontic patients. J. Dent. Res. 2012, 91, 853–858. [Google Scholar] [PubMed] [Green Version]
- Streiff, B.J.; Seneviratne, M.; Kingsley, K. Screening and prevalence of the novel cariogenic pathogen scardovia wiggsiae among adult orthodontic and non-orthodontic patient saliva samples. Int. J. Dent. Oral Health 2015, 1, 1–6. [Google Scholar]
- Reyes, N.; Pollock, A.; Whiteley, A.; Kingsley, K.; Howard, K. Prevalence of Scardovia wiggsiae among a pediatric Orthodontic patient population. EC Dent. Sci. 2017, 2, 203–210. [Google Scholar]
- Kressirer, C.A.; Smith, D.J.; King, W.F.; Dobeck, J.M.; Starr, J.R.; Tanner, A.C.R. Scardovia wiggsiae and its potential role as a caries pathogen. J. Oral Biosci. 2017, 59, 135–141. [Google Scholar] [CrossRef]
- Larsen, T.; Fiehn, N.E. Dental biofilm infections—An update. APMIS 2017, 125, 376–384. [Google Scholar] [CrossRef]
- Sutherland, I. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar]
- Asahi, Y.; Miura, J.; Tsuda, T.; Kuwabata, S.; Tsunashima, K.; Noiri, Y.; Sakata, T.; Ebisu, S.; Hayashi, M. Simple observation of Streptococcus mutans biofilm by scanning electron microscopy using ionic liquids. AMB Express 2015, 5, 6. [Google Scholar]
- Weber, K.; Delben, J.; Bromage, T.G.; Duarte, S. Comparison of SEM and VP-SEM imaging techniques with respect to Streptococcus mutans biofilm topography. FEMS Microbiol. Lett. 2014, 350, 175–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priester, J.H.; Horst, A.M.; Van de Werfhorst, L.C.; Saleta, J.L.; Mertes, L.A.; Holden, P.A. Enhanced visualization of microbial biofilms by staining and environmental scanning electron microscopy. J. Microbiol. Methods 2007, 68, 577–587. [Google Scholar] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar]
- Kachlany, S.C.; Levery, S.B.; Kim, J.S.; Reuhs, B.L.; Lion, L.W.; Ghiorse, W.C. Structure and carbohydrate analysis of the exopolysaccharide capsule of Pseudomonas putida G7. Environ. Microbiol. 2001, 3, 774–784. [Google Scholar] [CrossRef]
- Fassel, T.A.; Edmiston, C.E. Bacterial biofilms: Strategies for preparing glycocalyx for electron microscopy. Methods Enzym. 1999, 310, 194–203. [Google Scholar]
- Little, B.; Wagner, P.; Ray, R.; Pope, R.; Scheetz, R. Biofilms: An ESEM evaluation of artifacts introduced during SEM preparation. J. Ind. Microbiol. 1991, 8, 213–221. [Google Scholar] [CrossRef]
- Reese, S.; Guggenheim, B. A novel TEM contrasting technique for extracellular polysaccharides in in vitro biofilms. Microsc. Res. Tech. 2007, 70, 816–822. [Google Scholar] [CrossRef]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-biofilm activity of a long-chain fatty aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis biofilm. Front. Cell. Infect. Microbiol. 2017, 23, 46. [Google Scholar]
- Chang, H.T.; Rittmann, B.E. Biofilm loss during sample preparation for scanning electron microscopy. Water Res. 1986, 20, 1451–1456. [Google Scholar] [CrossRef]
- Vitkov, L.; Hermann, A.; Krautgartner, W.D.; Herrmann, M.; Fuchs, K.; Klappacher, M.; Hannig, M. Chlorhexidine-induced ultrastructural alterations in oral biofilm. Microsc. Res. Tech. 2005, 68, 85–89. [Google Scholar] [CrossRef]
- Vitkov, L.; Krautgartner, W.D.; Hannig, M. Bacterial internalization in periodontitis. Oral Microbiol. Immunol. 2005, 20, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Vitkov, L.; Hannig, M.; Krautgartner, W.D.; Herrmann, M.; Fuchs, K.; Klappacher, M.; Hermann, A. Ex vivo gingival-biofilm consortia. Lett. Appl. Microbiol. 2005, 41, 404–411. [Google Scholar] [PubMed]
- Erlandsen, S.L.; Kristich, C.J.; Dunny, G.M.; Wells, C.L. High-resolution visualization of the microbial glycocalyx with low-voltage scanning electron microscopy: Dependence on cationic dyes. J. Histochem. Cytochem. 2004, 52, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, T.P.; Dart, J.K.; McLaughlin-Borlace, L.; Howes, R.; Matheson, M.; Cree, I. Demonstration of biofilm in infectious crystalline keratopathy using ruthenium red and electron microscopy. Ophthalmology 2001, 108, 1088–1092. [Google Scholar] [CrossRef]
- Luft, J.H. Electron microscopy of cell extraneous coats as revealed by ruthenium red staining. J. Cell Biol. 1964, 23, 54A–55A. [Google Scholar]
- Fassel, T.A.; Schaller, M.J.; Remsen, C.C. Comparison of alcian blue and ruthenium red effects on preservation of outer envelope ultrastructure in methanotrophic bacteria. Microsc. Res. Tech. 1992, 20, 87–94. [Google Scholar] [CrossRef]
- Fassel, T.A.; Edmiston, C.E., Jr. Ruthenium red and the bacterial glycocalyx. Biotech. Histochem. 1999, 74, 194–212. [Google Scholar]
- Carr, K.E.; Toner, P.G. Cell Structure: An Introduction to Biomedical Electron Microscopy; Churchill Livingstone: Edinburgh, UK, 1982. [Google Scholar]
- Murakami, T.; Iida, N.; Taguchi, T.; Ohtani, O.; Kikuta, A.; Ohtsuka, A.; Itoshima, T. Conductive staining of biological specimens for scanning electron microscopy with special reference to ligand-mediated osmium impregnation. Scan. Electron Microsc. 1983, 1, 235–246. [Google Scholar]
- Murakami, T. A revised tannin-osmium method for non-coated SEM specimens. Arch. Histol. Jap. 1974, 36, 189–193. [Google Scholar]
- Murakami, T. A metal impregnation method of biological specimens for SEM. Arch. Histol. Jap. 1973, 35, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T. Puncture perfusion of small tissue pieces for SEM. Arch. Histol. Jap. 1976, 39, 99–103. [Google Scholar] [PubMed] [Green Version]
- Murakami, T.; Yamamoto, K.; Itoshima, T.; Irino, S. Modified tannin osmium conductive staining method for non-coated SEM specimens. Its application to microdissection SEM of the spleen. Arch. Histol. Jap. 1977, 40, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunji, T.; Wakita, M.; Kobayashi, S. Conductive staining in SEM with especial reference to tissue transparency. Scanning 1980, 3, 227–232. [Google Scholar]
- Arimoto, S.; Sugimura, M.; Kageyama, H.; Torimoto, T.; Kuwabata, S. Development of new techniques for scanning electron microscope observation using ionic liquid. Electrochim. Acta 2008, 53, 6228–6234. [Google Scholar]
- Tsuda, T.; Nemoto, N.; Kawakami, K.; Mochizuki, E.; Kishida, S.; Tajiri, T.; Kushibiki, T.; Kuwabata, S. SEM observation of wet biological specimens pretreated with room-temperature ionic liquid. ChemBioChem 2011, 12, 2547–2550. [Google Scholar]
- Brodusch, N.; Waters, K.; Demers, H.; Gauvin, R. Ionic liquid-based observation technique for nonconductive materials in the scanning electron microscope: Application to the characterization of a rare earth ore. Microsc. Res. Tech. 2014, 77, 225–235. [Google Scholar] [CrossRef]
- Imashuku, S.; Kawakami, T.; Ze, L.; Kawai, J. Possibility of scanning electron microscope observation and energy dispersive X-ray analysis in microscale region of insulating samples using diluted ionic liquid. Microsc. Microanal. 2012, 18, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Lo Torto, F.; Relucenti, M.; Familiari, G.; Vaia, N.; Casella, D.; Matassa, R.; Miglietta, S.; Marinozzi, F.; Bini, F.; Fratoddi, I.; et al. The effect of postmastectomy radiation therapy on breast implants: material analysis on silicone and polyurethane prosthesis. Ann. Plast. Surg. 2018, 81, 228–234. [Google Scholar]
- Relucenti, M.; Miglietta, S.; Bove, G.; Donfrancesco, O.; Battaglione, E.; Familiari, P.; Barbaranelli, C.; Covelli, E.; Barbara, M.; Familiari, G. SEM BSE 3D image analysis of human incus bone affected by cholesteatoma ascribes to osteoclasts the bone erosion and VP-SEM dEDX analysis reveals new bone formation. Scanning 2020, 2020, 9371516. [Google Scholar] [CrossRef] [Green Version]
- Relucenti, M.; Miglietta, S.; Covelli, E.; Familiari, P.; Battaglione, E.; Familiari, G.; Barbara, M. Ciliated cell observation by SEM on the surface of human incudo-malleolar-joint articular cartilage: Are they a new chondrocyte phenotype? Acta Otolaryngol. 2019, 139, 439–443. [Google Scholar]
- Timp, W.; Matsudaira, P. Electron microscopy of hydrated samples. Methods Cell Biol. 2008, 89, 391–407. [Google Scholar] [PubMed]
- Alhede, M.; Qvortrup, K.; Liebrechts, R.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Combination of microscopic techniques reveals a comprehensive visual impression of biofilm structure and composition. FEMS Immunol. Med. Microbiol. 2012, 65, 335–342. [Google Scholar] [PubMed] [Green Version]
- Araujo, J.C.; Teran, F.C.; Oliveira, R.A.; Nour, E.A.; Montenegro, M.A.; Campos, J.R.; Vazoller, R.F. Comparison of hexamethyldisilazane and critical point drying treatments for SEM analysis of anaerobic biofilms and granular sludge. J. Electron Microsc. 2003, 52, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, G.; Piccolomini, R.; Paludi, D.; D’Orio, V.; Vergara, A.; Conter, M.; Ianieri, A. Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 2008, 104, 1552–1561. [Google Scholar]
- Karcz, J.; Bernas, T.; Nowak, A.; Talik, E.; Woznica, A. Application of lyophilization to prepare the nitrifying bacterial biofilm for imaging with scanning electron microscopy. Scanning 2012, 34, 26–36. [Google Scholar]
- Bossù, M.; Matassa, R.; Relucenti, M.; Iaculli, F.; Salucci, A.; Di Giorgio, G.; Familiari, G.; Polimeni, A.; Di Carlo, S. Morpho-Chemical Observations of Human Deciduous Teeth Enamel in Response to Biomimetic Toothpastes Treatment. Materials (Basel) 2020, 13, 1803. [Google Scholar] [CrossRef] [Green Version]
- Bossù, M.; Saccucci, M.; Salucci, A.; Di Giorgio, G.; Bruni, E.; Uccelletti, D.; Sarto, M.S.; Familiari, G.; Relucenti, M.; Polimeni, A. Enamel remineralization and repair results of Biomimetic Hydroxyapatite toothpaste on deciduous teeth: An effective option to fluoride toothpaste. J. Nanobiotechnol. 2019, 17, 17. [Google Scholar]
- Redler, A.; Miglietta, S.; Monaco, E.; Matassa, R.; Relucenti, M.; Daggett, M.; Ferretti, A.; Familiari, G. Ultrastructural Assessment of the Anterolateral Ligament. Orthop. J. Sports Med. 2019, 7, 2325967119887920. [Google Scholar] [CrossRef] [Green Version]
- Ishii, S.; Koki, J.; Unno, H.; Hori, K. Two morphological types of cell appendages on a strongly adhesive bacterium, Acinetobacter sp. strain Tol 5. Appl. Environ. Microbiol. 2004, 70, 5026–5029. [Google Scholar]
- Weimer, P.J.; Price, N.P.; Kroukamp, O.; Joubert, L.M.; Wolfaardt, G.M.; Van Zyl, W.H. Studies of the extracellular glycocalyx of the anaerobic cellulolytic bacterium Ruminococcus albus 7. Appl. Environ. Microbiol. 2006, 72, 7559–7566. [Google Scholar] [CrossRef] [Green Version]
- Li, W.W.; Yu, H.Q. Insight into the roles of microbial extracellular polymer substances in metal biosorption. Bioresour. Technol. 2014, 160, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Dawes, C.J. Biological Techniques in Electron Microscopy; Barnes and Noble: New York, NY, USA, 1971. [Google Scholar]
- Gerhardt, P.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994. [Google Scholar]
- Torimoto, T.; Tsuda, T.; Okazaki, K.; Kuwabata, S. New frontiers in materials science opened by ionic liquids. Adv. Mater. 2010, 22, 1196–1221. [Google Scholar] [CrossRef] [PubMed]
- Kuwabata, S.; Kongkanand, A.; Oyamatsu, D.; Torimoto, T. Observation of ionic liquid by scanning electron microscope. Chem. Lett. 2006, 35, 600–601. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Familiari, G.; Heyn, R.; Petruzziello, L.; Relucenti, M. A method to visualize the micro architecture of glycoprotein matrices with scanning electron microscopy. In Scanning Electron Microscopy for the Life Sciences; Schatten, H., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 165–178. [Google Scholar]
- Relucenti, M.; Heyn, R.; Correr, S.; Familiari, G. Cumulus oophorus extracellular matrix in the human oocyte: A role for adhesive proteins. Ital. J. Anat. Embryol. 2005, 110 (Suppl. 1), 219–224. [Google Scholar]
- Familiari, G.; Relucenti, M.; Ermini, M.; Verlengia, C.; Nottola, S.A.; Motta, P.M. The human zona pellucida and scanning electron microscopy. Reality or artifacts? Ital. J. Anat. Embryol. 2001, 106 (Suppl. 2), 33–41. [Google Scholar]
- Familiari, G.; Nottola, S.A.; Macchiarelli, G.; Familiari, A.; Motta, P.M. A technique for exposure of the glycoproteic matrix (zona pellucida and mucus) for scanning electron microscopy. Microsc. Res. Tech. 1992, 23, 225–229. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protocols | ||||
|---|---|---|---|---|
| Steps | Conventional SEM | VP-SEM | OsO4-RR-TA-IL | |
| 1 | Fixation | Glutharaldehyde 2,5% in PB 0.1 M pH 7.4 at least 48 h | ||
| 2 | Washing | 10 min ×2 times in PB 0.1 M pH 7.4 | ||
| 3 | Postfixation | OsO4 2% 1 h | OsO4 2% in 1 h | OsO4 2% + RR 0.2% 1:1 solution, 1 h |
| 4 | Washing | 10 min ×2 times in dH20 10 min ×2 times in dH20 10 min ×2 times in dH20 | ||
| 5 | Impregnation | None | None | Tannic Acid 1% in d H20 30 min |
| 6 | Washing | continues from 2 | continues from 2 | 10 min ×2 times in dH20 |
| 7 | Dehydration | Ascending ethanol series | None | None |
| 8 | Drying | Ascending HMDS 1 series | None | None |
| 9 | Pt Sputter coating | 15 mA, 2 min | None | Replaced by IL |
| Operating conditions | 1520– kV, high vacuum | 510–kV 30 Pa | 1520– kV, high vacuum | |
| Parameters | Protocols | ||
|---|---|---|---|
| Conventional SEM | VP-SEM | OsO4-RR-TA-IL | |
| Procedure time | 2 days | 1 h and 30 min | 2 h and 10 min |
| Sample loss | Steps produce sample loss of about 60% | about 20% | about 20% |
| Dehydration and drying | yes | None | None |
| Pt sputter coating | yes | None | Replaced by IL |
| Resistance in vacuum | Excellent, it is possible to observe for hours | Good for 1 hour | Excellent, it is possible to observe for hours |
| Operating conditions | 1520 kV, high vacuum | 510 kV 30 Pa | 1520 kV, high vacuum |
| Image magnification | Good up to 40k | Good up to 10k | Good up to 30k |
| Image quality | Excellent up to 30k | Good up to 8k | Excellent up to 30k |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bossù, M.; Selan, L.; Artini, M.; Relucenti, M.; Familiari, G.; Papa, R.; Vrenna, G.; Spigaglia, P.; Barbanti, F.; Salucci, A.; et al. Characterization of Scardovia wiggsiae Biofilm by Original Scanning Electron Microscopy Protocol. Microorganisms 2020, 8, 807. https://doi.org/10.3390/microorganisms8060807
Bossù M, Selan L, Artini M, Relucenti M, Familiari G, Papa R, Vrenna G, Spigaglia P, Barbanti F, Salucci A, et al. Characterization of Scardovia wiggsiae Biofilm by Original Scanning Electron Microscopy Protocol. Microorganisms. 2020; 8(6):807. https://doi.org/10.3390/microorganisms8060807
Chicago/Turabian StyleBossù, Maurizio, Laura Selan, Marco Artini, Michela Relucenti, Giuseppe Familiari, Rosanna Papa, Gianluca Vrenna, Patrizia Spigaglia, Fabrizio Barbanti, Alessandro Salucci, and et al. 2020. "Characterization of Scardovia wiggsiae Biofilm by Original Scanning Electron Microscopy Protocol" Microorganisms 8, no. 6: 807. https://doi.org/10.3390/microorganisms8060807