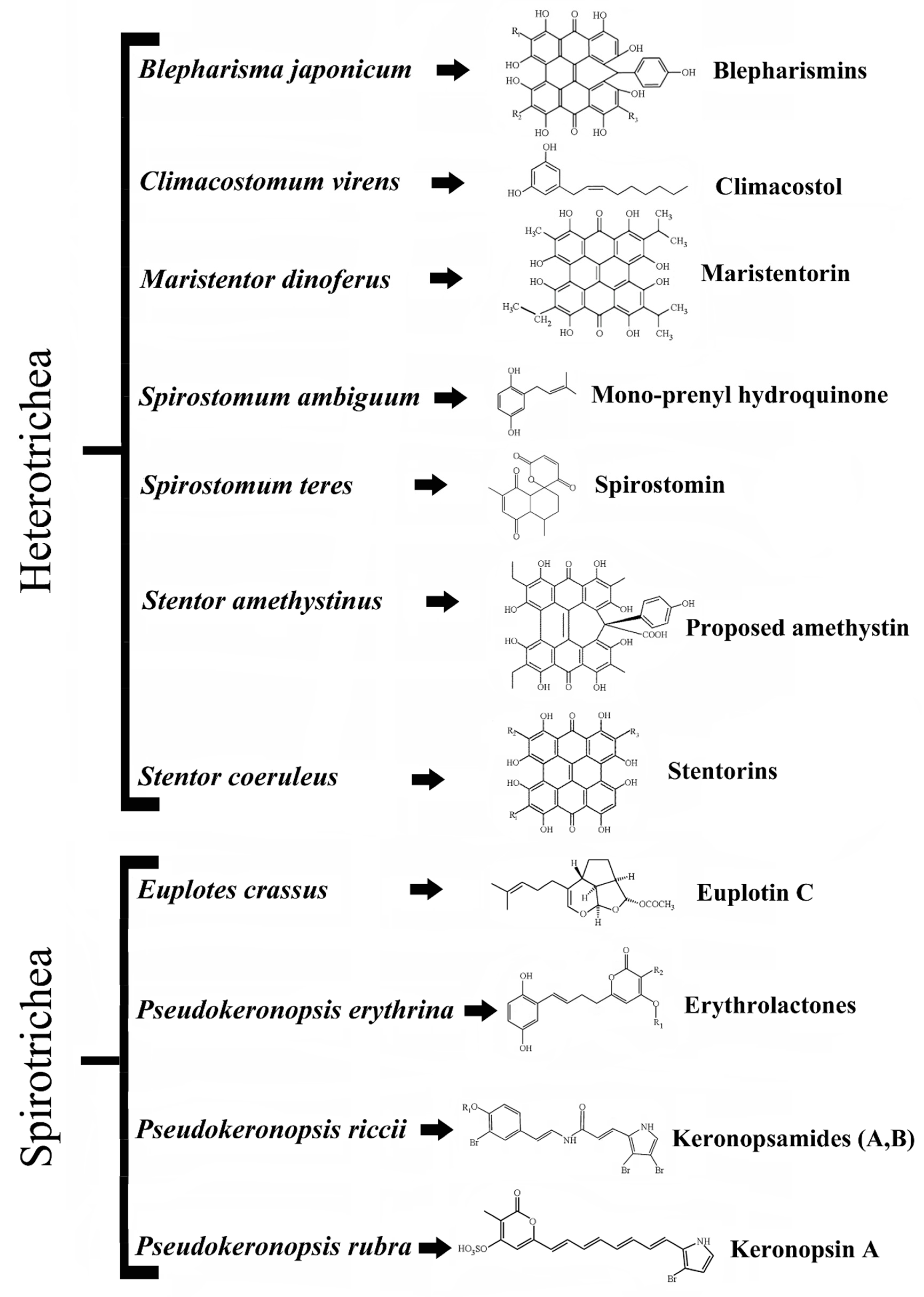
1. Introduction
Aquatic organisms live in environments which are exceptionally full of competitive biodiversity and for this reason have evolved a great number of adaptive strategies to survive and reproduce, which in eukaryotic unicellular microbes (protists) appear particularly well represented. At the molecular level, ciliated protists show adaptive strategies based on at least two classes of compounds, pheromones and secondary metabolites. Pheromones are diffusible proteins produced and secreted by different ciliate species to mediate self–nonself recognition mechanisms, responsible for cell shift between the vegetative and sexual stages of the cell biological cycle (see [
1] for a review). Secondary metabolites produced by ciliates include toxic substances either stored in the cell cytoplasm or inside ejectable organelles (known as extrusomes) that are anchored to the cell cortex before being used in predator-prey and cell-environment interactions [
2,
3]. These secondary metabolites are synthesized through different biogenetic routes and have been named from the taxonomic names of their source organisms. Some metabolites are biosynthesized via the polyketide pathway (or mixed biogenetic routes), whilst several others derive from the classical terpenoid biogenesis (see [
3,
4] for a review). Representative examples are the terpenoid euplotins from
Euplotes species [
5] and some polycyclic aromatic compounds built on hypericin structure such as blepharismins from
Blepharisma species [
6,
7], stentorins from
Stentor coeruleus [
8], amethystin from
Stentor amethystinus [
9], and maristentorin from
Maristentor dinoferus [
10]. Other toxic secondary metabolites are spirostomin from
Spirostomum teres [
11] and mono-prenyl hydroquinone from
Spirostomum ambiguum [
12]. Keronopsins, keronopsamides and erythrolactones are, instead, produced by three different species of the genus
Pseudokeronopsis:
P. rubra,
P. riccii and
P. erythrina [
7,
13,
14,
15,
16], respectively (
Figure 1).
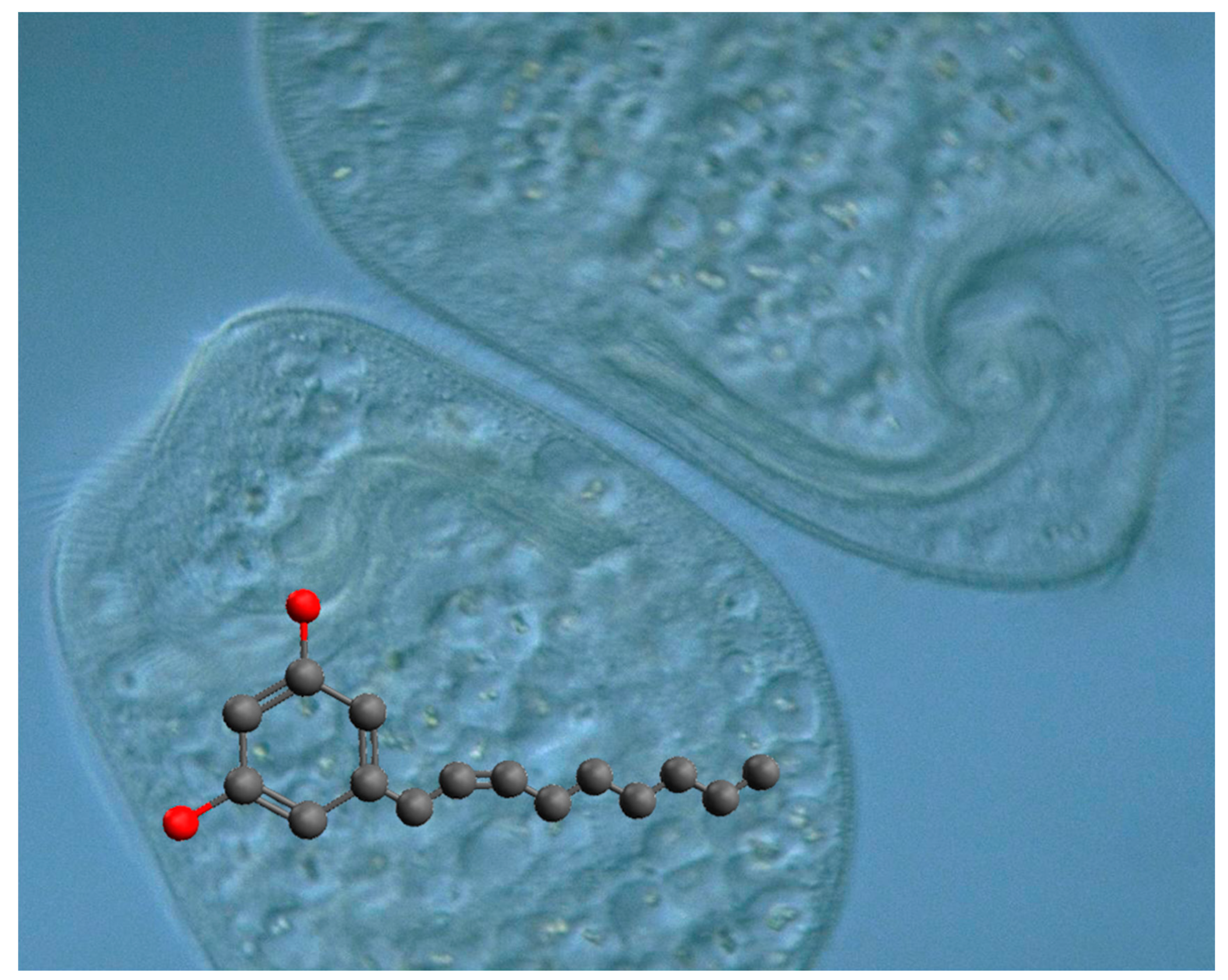
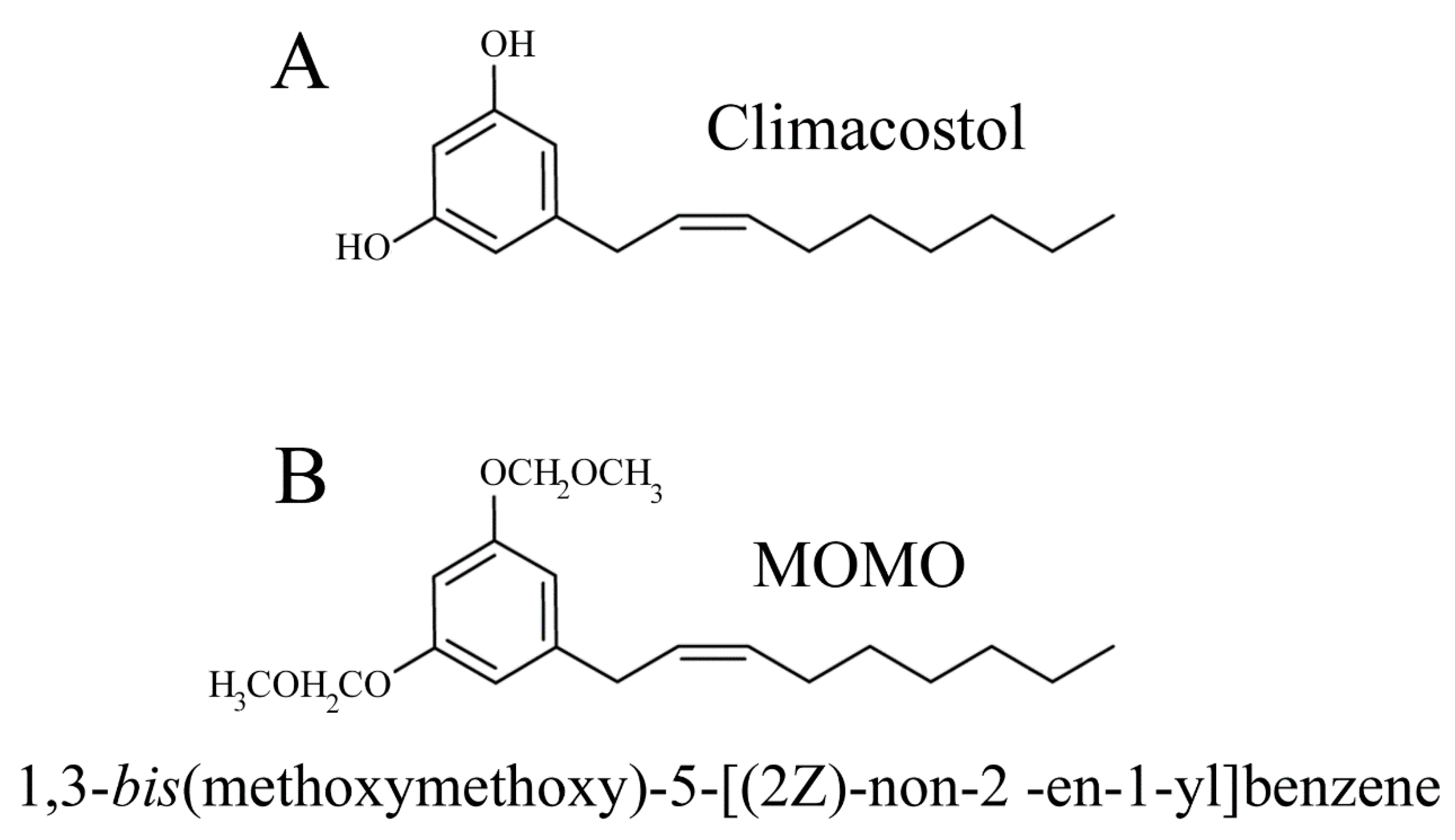
Climacostol (5-[(2
Z)-non-2-en-1-yl]benzene-1,3-diol) is a toxic secondary metabolite physiologically produced by the freshwater ciliate
Climacostomum virens (
Figure 2) for chemical defense against unicellular and multicellular predators, or for chemical offence to assist its carnivorous feeding [
3,
17]. This molecule is formed by a phenolic skeleton and a long unsaturated aliphatic hydrocarbon chain attached to the ring structure, and belongs to resorcinolic lipids (or alkenylresorcinols), a group of natural amphiphilic compounds detected in both prokaryotes and eukaryotes. The 5-alkenylresorcinols usually have an isolated double bond in the chain portion of the molecule, while examples with a conjugated double bond with aromatic ring are unusual [
18]. A possible pathway for the biosynthesis of climacostol was proposed from the C16-polyketide, with a cyclization and a decarboxylation [
19], and the toxin was also obtained by chemical synthesis [
19,
20]. More recently, climacostol has been synthetized as a pure compound in the natural and most bioactive
Z-configuration by a novel and straightforward synthesis [
21], which has allowed researchers to better study its effects on different biological systems.
2. Climacostol Mediates Predator–Prey Interactions
Climacostomum virens is a noncontractile freshwater heterotrich ciliate, which often looks green due to the presence of endosymbiotic algae (
Chlorella sp.). The green cells can be cultivated in the dark to reduce or eliminate the symbionts, in order to obtain colorless strains [
17,
22].
C. virens possess numerous extrusomes reported as cortical granules (or granulocysts) which are able to discharge their content to the outside of the cell in response to external stimuli. The colorless extruded substances are mainly represented by climacostol with the presence of some related analogues [
16,
18]. The defensive function of climacostol was first demonstrated against the raptorial ciliate
Dileptus margaritifer. Indeed, when this predator touches a cell of
C. virens with its toxicyst-bearing proboscis,
D. margaritifer swims backward and a condensed substance, raised from the site of contact, becomes visible under a dark-field microscope. After other attacks by the same individual, the proboscis of
D. margaritifer appears shorter and visibly damaged, whereas subsequent attacks can lead to the death of the predator due to the cytotoxic effect of climacostol [
17]. More recently, the toxic action of climacostol against metazoan predators was also demonstrated [
23], and it was furthermore assumed that
C. virens is able to use climacostol for chemical offense, to paralyze and kill prey [
3].
3. Climacostol Exerts Cytotoxic and Antimicrobial Effect
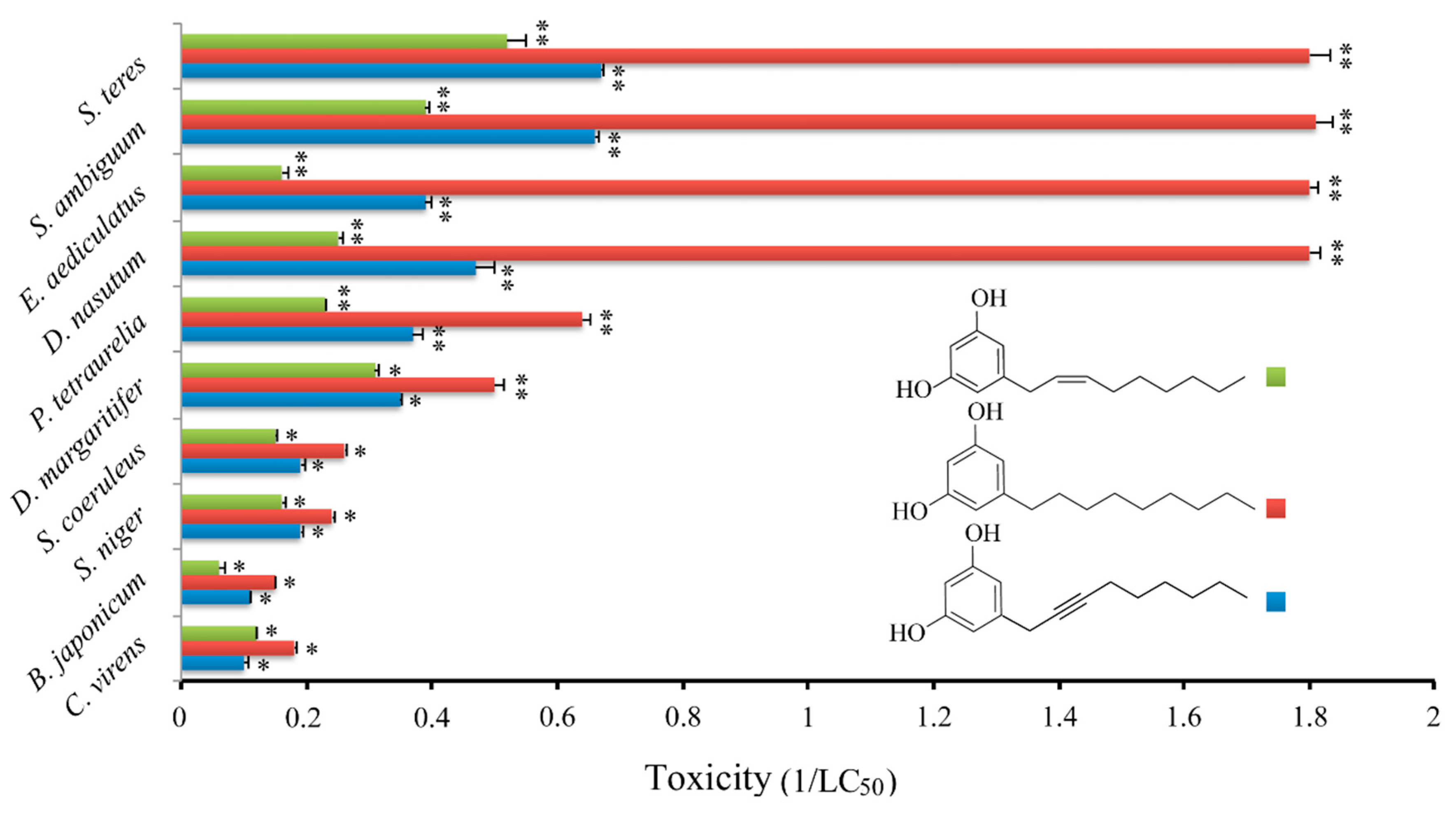
Initially, the cytotoxic effect of climacostol was analyzed against a panel of nine free-living ciliate species which share the same microhabitat with
C. virens:
Blepharisma japonicum,
Didinium nasutum,
Dileptus margaritifer,
Euplotes aediculatus,
Paramecium tetraurelia,
Spirostomum ambiguum,
Spirostomum teres,
Stentor coeruleus, and
Stentor niger. It was demonstrated that the cytotoxic potency of climacostol is related to the molecular structure of the unsaturated side chain, and that it could be modulated by the substitution of the double bond with a single or a triple one at the C
8 position [
24]. In fact, while the saturated alkyl derivative showed the highest cytotoxicity against the tested species, the alkynyl derivative that carries a triple bond in the side chain showed the lowest cytotoxic potency. An intermediate cytotoxicity was revealed by the native climacostol (with a double bond in the side chain) (
Figure 3).
In a further study, the effect of climacostol and its alkyl and alkynyl derivatives was also tested on some Gram-positive and Gram-negative pathogen bacteria and on the fungus
Candida albicans [
25]. The results showed an appreciable and comparable antimicrobial activity of the three compounds, which were effective against Gram-positive bacteria and
C. albicans with Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentration (MBC) ranging from 8 to 32 mg L
−1. By contrast, no significant toxicity against Gram-negative species (
Escherichia coli and
Pseudomonas aeruginosa) has been observed [
25]. The authors speculated that this limited effect observed on the Gram-negative species can be explained by the peculiar structure of the bacterial cell wall. The outer membrane is indeed a selective barrier to the penetration of several compounds, due to the hydrophilic nature of the surface exposed to the environment and to the selectivity of the outer membrane proteins. To summarize, the overall results suggest that the saturation rate of the side chain of climacostol does not couple to its antimicrobial activity, whereas it is closely related to the cytotoxic action against ciliated protists. Therefore, it is likely that the general structure of the two moieties of climacostol, i.e., the di-hydroxy-phenyl group and the alkenyl chains, contributes to the antibiotic action as a whole. In general, the presence of a double bond in the hydrocarbon chain is associated with an increase in cell viability as compared to the saturated compounds, and an increase in the chain length does not interfere with the effect of aromatic ring substituents, which is clearly predominant [
26].
4. Climacostol Reduces Tumor Progression via p53-Dependent Apoptosis
On the basis of anticancer activity displayed by a number of other resorcinolic lipids (see [
27] for a review), the effects of climacostol were initially explored
in vitro on some human and rodent tumor cell lines [
28]. In particular, experiments performed on human tumor squamous carcinoma A431 cells, and human promyelocytic leukemia HL60 cells demonstrated that climacostol exerts its action by inhibiting cell growth and triggering a mitochondrion-dependent apoptotic program. Subsequent extensive
in vitro screenings and
in vivo experiments confirmed the previous observations [
4,
21,
29,
30,
31]. Although climacostol cytotoxicity appears more selective against tumors than certain immortalized nontumor cells [
21,
29], recent data suggest that climacostol effects are not necessarily correlated to the cancerous origin of cells [
31]. However, the possibility that climacostol preferentially affects cancerous vs. normal (nontransformed non-immortalized) cells requires further investigation.
With regard to the action mechanism of climacostol, it was demonstrated that the protozoan toxin can trigger apoptosis by binding to nuclear and mitochondrial DNA, and promoting their cleavage after the generation of reactive oxygen species (ROS) in the presence of Cu(II) [
32,
33]. It is known that the induction of DNA damage could be effective in treating cancer, and many currently employed antitumor drugs, for example, platinum agents, function by means of this mechanism [
34,
35,
36]. In this role, it was shown that cisplatin and climacostol do not display any additive effects on melanoma viability [
29].
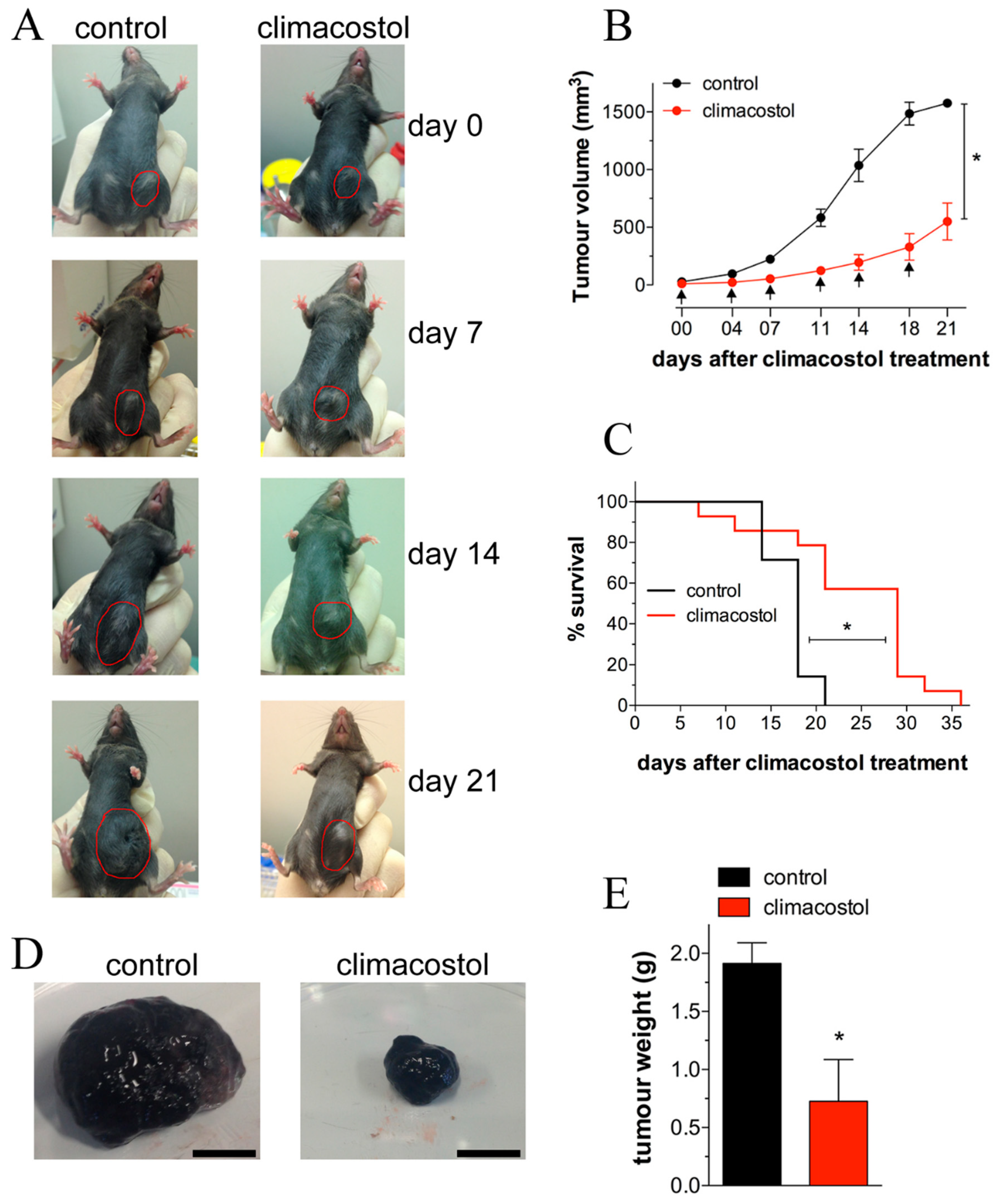
Recent investigations on the mechanisms of action of climacostol demonstrated that it reduces the viability/proliferation of melanoma cells, causing rapidly occurring DNA damage, also inducing the intrinsic apoptotic pathway characterized by the dissipation of the mitochondrial membrane potential, the translocation of Bax to the mitochondria, the release of cytochrome c from the mitochondria, and the activation of caspase 9-dependent cleavage of caspase 3 [
29,
30]. Additionally, persistent inhibition of the growth of melanoma allografts as well as a reduction in the number of viable and proliferating tumor cells was achieved after intratumoral injections of the toxin [
4,
29]. The notion that climacostol may induce a decrease in the microvessel sprouting that contributes to inhibition of melanoma growth was also suggested [
4]. As shown in
Figure 4, a significant improvement in survival of transplanted mice was reported, together with a decrease of tumor weight, and a reduction of viable cells inside the tumor. Of interest, the signaling events responsible for the climacostol-induced pro-apoptotic effects rely on the upregulation of the p53 network and its targets Noxa and Puma [
4].
5. Climacostol Induces Dysfunctional Autophagy in Tumor Cells
In all the eukaryotic cells, a pivotal role for the maintenance of homeostasis is played by autophagy, the highly conserved process which operates via the degradation of cytoplasmic organelles, proteins, and macromolecules, and the recycling of the breakdown products. As autophagy supports cell survival or activates death pathways, it may represent a potential target for the available library of anticancer substances and for the development of new molecules.
Some of us recently reported on how climacostol regulates autophagy and the involvement of p53-dependent mechanisms [
30]. Essentially, our data indicated that the protozoan toxin potently and selectively impairs autophagy in multiple tumor cells that are committed to dying by apoptosis. Climacostol exerts a marked and sustained accumulation of autophagosomes as the result of dysfunctional autophagic degradation. Mechanistic insights showed that climacostol affects autophagosome turnover via p53-AMPK axis, although the mTOR pathway unrelated to p53 levels plays a role. Of note are indications suggesting that climacostol effects on autophagy and apoptosis are actually two separate events, which may act independently on life/death decisions of the cell.
In this picture, the upregulation of the p53 system appears at the molecular crossroads regulating both the anti-autophagic action of climacostol and its role in the induction of apoptosis.
6. The Synthetic Analogues of Climacostol for Biotechnological Applications
The results collected in the last 20 years regarding the activity of climacostol together with the structural properties of other resorcinolic lipids allowed us to design and obtain new synthetic analogues of the protozoan toxin. These compounds shed light on the chemical bases of climacostol actions.
Two analogues, methyl-5 [(2
Z)-non-2-en-1-yl]benzene-1,3-diol (AN1) and 5-[(2
Z)-non-2-en-1-yl]benzene-1,2,3-triol (AN2), respectively carrying an additional methyl group and a hydroxyl group in the aromatic ring, were analyzed for their biological activity [
31] (
Figure 5).
The choice to introduce the aforementioned moieties into the aromatic ring of climacostol was supported by the observation that similar modification performed on some polyphenols and phenolic lipids resulted in a significant improvement of their cytotoxic and antimicrobial activity [
37,
38].
The effects of AN1 and AN2 were investigated on mammalian cells, pathogenic microbes and free-living ciliated protists, with the main purpose being to identify the structural traits of native climacostol primarily involved in its cytotoxic activities.
6.1. Antitumour Activity
The pro-apoptotic features of AN1 and AN2 on immortalized cell lines of both tumor (B16-F10, GL261, SK-N-BE, and CT26) and nontumor (C
2C
12) origin were analyzed and compared with that of climacostol. The results indicated that cell viability was negatively affected by the two analogues with a comparable strength (
Table 1), and both AN1 and AN2 displayed similar or even lower potencies when compared to climacostol [
31]. Furthermore, as in the case of various tumor cells exposed to climacostol [
4,
29,
30], immunostaining techniques revealed that B16-F10 melanoma cells expressed high levels of active caspase 3 after AN1 and AN2 treatment (
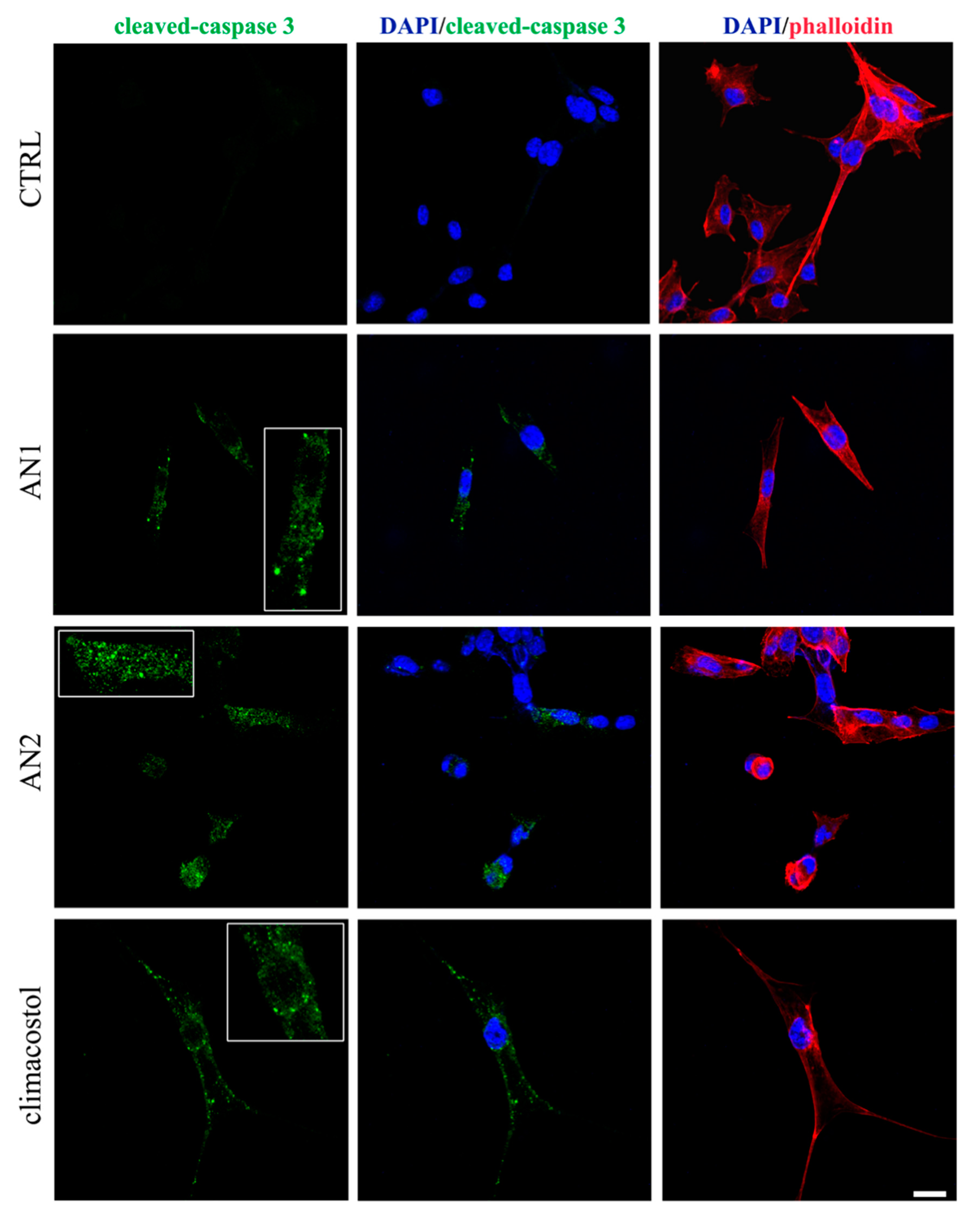
Figure 6), thus demonstrating the activation of an apoptotic pathway induced by both analogues.
6.2. Antimicrobial Activity
AN1 and AN2 were also investigated with dose–response experiments performed to compare their cytotoxic potential against a panel of microorganisms comprising bacterial and fungal pathogens, and freshwater ciliates.
The data collected confirmed that both AN1 and AN2 show an appreciable cytotoxicity on all the microorganisms exposed to different concentrations of the two analogues, with the exception of
Escherichia coli and
Pseudomonas aeruginosa that proved to be immune to the toxins. AN1 was the most toxic compound against pathogens and ciliates, whereas AN2 effects were comparable to or worse than climacostol. In particular, AN1 showed MIC and MBC values of 8 μg/mL against the Gram-positive
Staphylococcus aureus and
Enterococcus faecalis, respectively, and a value of 4 μg/mL against the fungus
Candida albicans. In experiments with ciliates, the highest toxicity of AN1 (0.64 μg/mL < LC50 < 2.15 μg/mL) was observed against
B. japonicum,
P. multimicronucleatum,
S. ambiguum, and
S. teres [
31].
With regard to the action mechanism of the synthetic toxins, the cytotoxicity of both AN1 and AN2 on some ciliate species appeared to be mediated by a necrotic process, as previously reported for the effect of climacostol on free-living ciliates [
17,
24]. An exception was represented by the ciliate
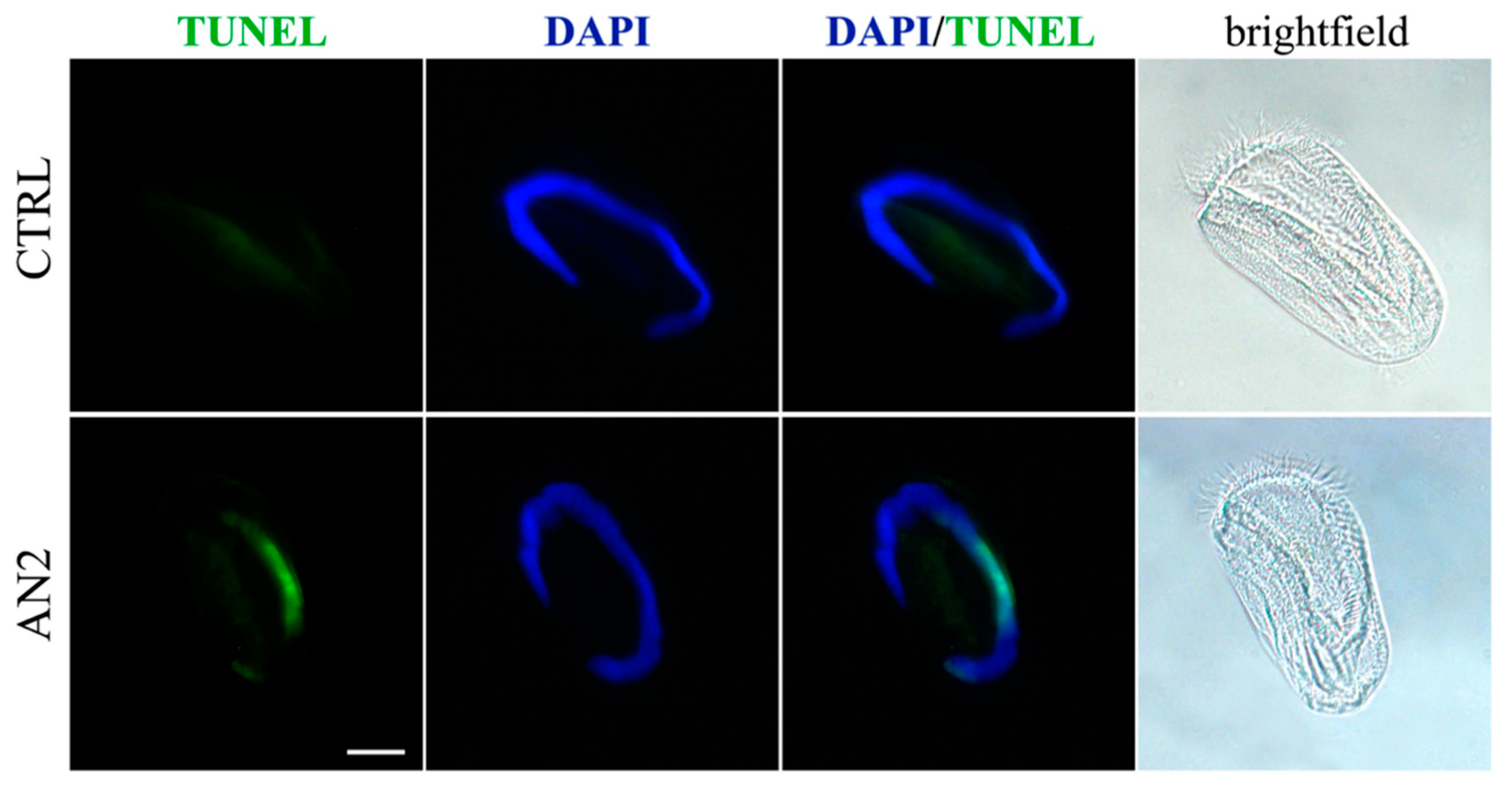
Euplotes aediculatus which displayed the activation of programmed cell death triggered by AN2. In fact, Terminal deoxyribonucleotidyl transferse (TdT)-mediated biotin-16-dUTP Nick-End Labelling (TUNEL) fluorescence assay and light microscopy observations on
Euplotes specimens exposed to AN2 revealed the typical sequence of events associated with canonical apoptosis, such as the progressive fragmentation of the macronucleus during the early stages in the absence of major changes in cell morphology, and the lack of severe necrotic damage, such as cell swelling and rupture [
31] (
Figure 7). To summarize, whereas similar effects were substantially observed for climacostol and its analogues on mammalian cells, AN1 was the most active compound against bacterial and fungal pathogens, and protists. The hydroxyl group added to climacostol to obtain AN2 appears to be the pivotal structural trait transforming the protozoan toxin into an apoptosis-inducing compound in unicellular eukaryotes.
The overall results obtained with AN1 and AN2 encourage the attempt to design and synthetize additional novel analogues of climacostol that can increase or optimize its pharmacological properties.
7. Climacostol as a Prodrug
During the synthesis of climacostol, the choice of the methoxymethyl ether (MOM) protecting group, which can be removed in a weakly acidic environment, allowed the researchers to efficiently obtain the toxin in the biologically active (
Z)-configuration climacostol [
21] (
Figure 8).
This procedure was also applied to design a prodrug strategy based on pH activation, potentially useful to deliver climacostol to acidic pathological multicellular (tumors) or unicellular (parasitophorous of
Leishmania and digestive vacuole of
Plasmodium) targets [
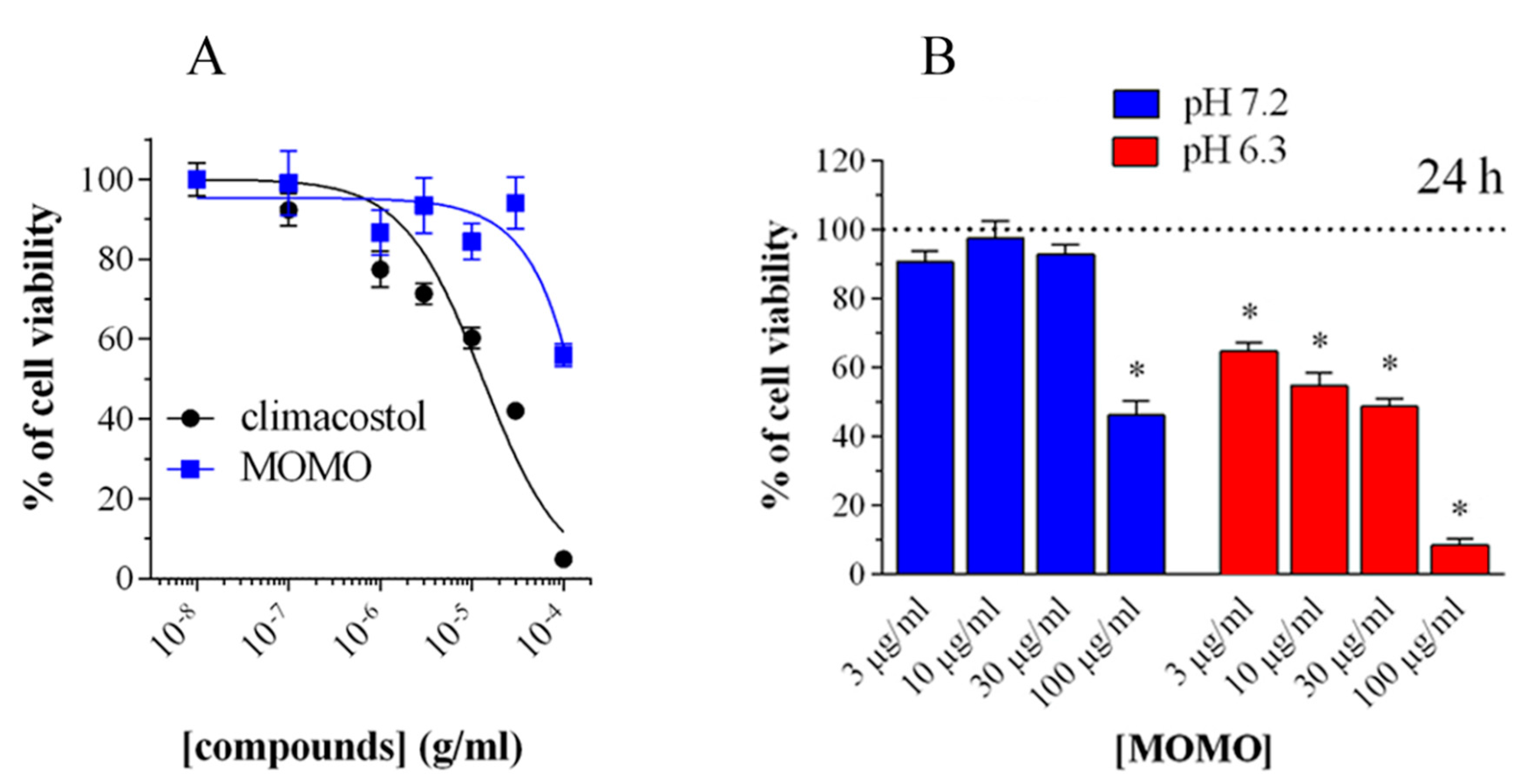
39]. In fact, the MOM-protected climacostol, which the authors called MOMO, progressively shifts to active climacostol if exposed to pH values lower than 7. This shifting was initially verified in a native biological system (against ciliates other than
C. virens) where ciliates exposed to MOMO in a physiological culture medium (SMB) at pH = 6.8 did not reveal any sign of toxicity. On the contrary, MOMO resuspended in SMB at pH = 6.3 (not harmful itself for ciliates) induced the same necrotic effect as climacostol [
39].
In addition, it was demonstrated that the activity of MOMO could be modulated in a dose-dependent manner to induce cytotoxic or cytostatic effects. The biological activity of MOMO on some tumor cell lines confirmed that this molecule proved toxic in acidic conditions (pH < 6.8), by yielding active climacostol. In these conditions, MOMO induces apoptosis in melanoma cells confirming that it behaves with the same action mechanism as climacostol (
Figure 9).
Finally, the safety of MOMO was also assessed in vivo using the fruit fly Drosophila melanogaster which is considered a very potent tool to detect the potential damaging effect of new compounds. Drosophila flies fed directly with acidic nutrients containing increasing concentrations of MOMO showed a significant reduction in oviposition and survival of larvae, which also exhibited apoptotic cells in the gut, as revealed by TUNEL labeling and cleaved-caspase 3 immunostaining. In addition, mitosis was also reduced in brain tissue of larvae, thus suggesting that active MOMO exhibited prolonged toxic effects after oral intake and gut absorption.
Overall, our data revealed that MOMO efficiently targets different tissues of the developing fly with high metabolic/proliferating activity, such as midgut and brain.
8. Conclusions
Natural products possess enormous structural and chemical diversity that is unsurpassed by any synthetic libraries. Notably, small organic molecules from nature have shown great translational potential [
40] since living organisms have a marked ability to synthetize complex molecular structures with defined biological properties, and climacostol is a typical example as a small molecule. However, the possibility to directly purify climacostol as a natural compound from cell cultures is actually limited to very small amounts. Therefore, to address this problem, many efforts have been directed towards the preparation of large amounts of synthetic climacostol and some analogues, through strategies capable of making a cheaper and faster product.
The studies carried out in recent years on climacostol and its analogues have shown how the alkenylresorcinol structure can exert both appreciable antimicrobial and antitumor activities. The resorcinol moiety is in fact a widespread phenolic unit, which is found in several biologically active natural products [
41], as well as a synthetic derivative with biological activities that show considerable therapeutic potential.
Structure–activity relationship investigations in distinct alkenylresorcinols have shown that an increase in the number of the ring substituents leads to higher cytotoxic activities in many cell lines compared to climacostol. Therefore, it is interesting to note that an increase in the number of hydroxyl groups in climacostol does not lead to an increase in antitumor activity, which can be comparable to or worse than that of the native toxin. In this picture, further synthetic studies to better understand the relationship between the biological activity of climacostol and the position of the substituents in its aromatic ring are currently under development. The biological results will be reported as soon as they are available.
In conclusion, it appears that small molecule derivatives exhibiting optimum potency/selectivity may have a pivotal role to play in understanding the biochemical machinery underlying cell function/dysfunction as well as in exploiting new therapeutic indications. In this respect, climacostol appears to be a promising lead compound for chemical organic approaches and biotechnological investigations which merit exploitation by drug pharmacology.
Author Contributions
Conceptualization: F.B., E.C., D.C., C.C., E.M., C.O.; Writing—Original Draft Preparation: F.B., C.O.; Writing—Review and Editing: F.B., E.C., D.C., C.C., E.M., C.O.; Visualization; F.B., E.C., D.C., C.C., E.M., C.O.; Supervision, C.O.; Funding Acquisition, F.B., D.C., E.M., C.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by University of Macerata.
Acknowledgments
The authors wish to thank Gill Philip (University of Macerata) for the linguistic revision of the text and Akira Kihara (Laboratory of Biology, Science Research Center, Hosei University, Japan) for permitting the open use of the picture from Protist Information Server (
http://protist.i.hosei.ac.jp), an important source for protistological dissemination. Elisabetta Catalani is a recipient of “Departments of Excellence-2018” (DIBAF Project “Landscape 4.0—food, wellbeing and environment”) postdoctoral fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Luporini, P.; Alimenti, C.; Vallesi, A. Ciliate pheromones structures and activity: A review. Ital. J. Zool. 2015, 82, 3–14. [Google Scholar] [CrossRef]
- Rosati, G.; Modeo, L. Extrusomes in ciliates: Diversification, distribution, and phylogenetic implications. J. Eukaryot. Microbiol. 2003, 50, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, F.; Ortenzi, C. Predator-prey interactions in ciliated protists. In Extremophilic Microbes and Metabolites—Diversity, Bioprespecting and Biotechnological Applications, 1st ed.; InTechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Catalani, E.; Proietti Serafini, F.; Zecchini, S.; Picchietti, S.; Fausto, A.M.; Marcantoni, E.; Buonanno, F.; Ortenzi, C.; Perrotta, C.; Cervia, D. Natural products from aquatic eukaryotic microorganisms for cancer therapy: Perspectives on anti-tumour properties of ciliate bioactive molecules. Pharmacol. Res. 2016, 113, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Guella, G.; Skropeta, D.; Di Giuseppe, G.; Dini, F. Structures, biological activities and phylogenetic relationship of terpenoids from marine ciliates of the genus Euplotes. Mar. Drugs 2010, 8, 2080–2116. [Google Scholar] [CrossRef] [Green Version]
- Harumoto, T.; Miyake, A.; Ishikawa, N.; Sugibayashi, R.; Zenfuku, K.; Iio, H. Chemical defense by means of pigmented extrusomes in the ciliate Blepharisma japonicum. Eur. J. Protistol. 1998, 34, 458–470. [Google Scholar] [CrossRef]
- Buonanno, F.; Anesi, A.; Guella, G.; Ortenzi, C. Blepharismins used for chemical defense in two ciliate species of the genus Blepharisma, B. stoltei and B. undulans (Ciliophora: Heterotrichida). Europ. Zool. J. 2017, 84, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Miyake, A.; Harumoto, T.; Iio, H. Defensive function of pigment granules in Stentor coeruleus. Eur. J. Protistol. 2001, 37, 77–88. [Google Scholar] [CrossRef]
- Höfle, G.; Reinecke, S.; Laude, U.; Kabbe, K.; Dietrich, S. Amethystin, the coloring principle of Stentor amethystinus. J. Nat. Prod. 2014, 77, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Fulton, D.B.; Halder, M.; Han, X.; Armstrong, D.W.; Petrich, J.W.; Lobban, C.S. Maristentorin, a novel pigment from the positively phototactic marine ciliate Maristentor dinoferus, is structurally related to hypericin and stentorin. J. Phys. Chem. 2006, 110, 6359–6364. [Google Scholar] [CrossRef]
- Sera, Y.; Masaki, M.E.; Doe, M.; Buonanno, F.; Miyake, A.; Usuki, Y.; Iio, H. Spirostomin, defense toxin of the ciliate Spirostomum teres: Isolation, structure elucidation, and synthesis. Chem. Lett. 2015, 44, 633–635. [Google Scholar] [CrossRef] [Green Version]
- Buonanno, F.; Guella, G.; Strim, C.; Ortenzi, C. Chemical defence by mono-prenyl hydroquinone in a freshwater ciliate, Spirostomum ambiguum. Hydrobiologia 2012, 684, 97–107. [Google Scholar] [CrossRef]
- Höfle, G.; Pohlan, S.; Uhlig, G.; Kabbe, K.; Schumacher, D. Keronopsins A and B, chemical defence substances of the marine ciliate Pseudokeronopsis rubra (Protozoa): Identification by ex vivo HPLC. Angew. Chem. Int. Ed Engl. 1994, 33, 1495–1497. [Google Scholar] [CrossRef]
- Guella, G.; Frassanito, R.; Mancini, I.; Sandron, T.; Modeo, L.; Verni, F.; Dini, F.; Petroni, G. Keronopsamides, a new class of pigments from marine ciliates. Europ. J. Org. Chem. 2010, 3, 427–434. [Google Scholar] [CrossRef]
- Anesi, A.; Buonanno, F.; Di Giuseppe, G.; Ortenzi, C.; Guella, G. Metabolites from the Euryhaline Ciliate Pseudokeronopsis erythrina. Eur. J. Org. Chem. 2016, 1330–1336. [Google Scholar] [CrossRef]
- Buonanno, F.; Anesi, A.; Di Giuseppe, G.; Guella, G.; Ortenzi, C. Chemical defense by erythrolactones in the euryhaline ciliated protist, Pseudokeronopsis erythrina. Zool. Sci. 2017, 34, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Buonanno, F.; Saltalamacchia, P.; Masaki, M.E.; Iio, H. Chemical defence by means of extrusive cortical granules in the heterotrich ciliate Climacostomum virens. Eur. J. Protistol. 2003, 39, 25–36. [Google Scholar] [CrossRef]
- Yang, S.-W.; Chan, T.-M.; Terracciano, J.; Loebenberg, D.; Patel, M.; Gullo, V.; Chu, M. A new 5-alkenylresorcinol Sch 725681 from Aspergillus sp. J. Antibiot. 2006, 59, 190–192. [Google Scholar] [CrossRef]
- Masaki, M.E.; Hiro, S.; Usuki, Y.; Harumoto, T.; Terazima, M.N.; Buonanno, F.; Miyake, A.; Iio, H. Climacostol, a defense toxin of Climacostomum virens (protozoa, ciliata), and its congeners. Tetrahedron 2004, 60, 7041–7048. [Google Scholar] [CrossRef]
- Masaki, M.E.; Harumoto, T.; Terazima, M.N.; Miyake, A.; Usuki, Y.; Iio, H. Climacostol, a defense toxin of the heterotrich ciliate Climacostomum virens against predators. Tetrahedron Lett. 1999, 40, 8227–8229. [Google Scholar] [CrossRef]
- Fiorini, D.; Giuli, S.; Marcantoni, E.; Quassinti, L.; Bramucci, M.; Amantini, C.; Santoni, G.; Buonanno, F.; Ortenzi, C. A Straightforward diastereoselective synthesis and evaluation of climacostol, a natural product with anticancer activities. Synthesis 2010, 9, 1550–1556. [Google Scholar] [CrossRef]
- Karajan, B.; Vishnjakov, A.; Tavrovskaya, M.; Vasjanin, S. Infection of algae-free Climacostomum virens with symbiotic Chlorella sp. isolated from algae-containing C. virens. Europ. J. Protistol. 2007, 43, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, F.; Harumoto, T.; Ortenzi, C. The defensive function of trichocysts in Paramecium tetraurelia against metazoan predators compared with the chemical defense of two species of toxin-containing ciliates. Zool. Sci. 2013, 30, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, F.; Ortenzi, C. The protozoan toxin climacostol and its derivatives: Cytotoxicity studies on 10 species of free-living ciliates. Biologia 2010, 65, 675–680. [Google Scholar] [CrossRef] [Green Version]
- Petrelli, D.; Buonanno, F.; Vitali, L.A.; Ortenzi, C. Antimicrobial activity of the protozoan toxin climacostol and its derivatives. Biologia 2012, 67, 525–529. [Google Scholar] [CrossRef]
- Kruk, J.; Aboul-Enein, B.; Bernstein, J.; Marchlewicz, M. Dietary alkylresorcinols and cancer prevetion: A systematic review. Eur. Food Res. Technol. 2017, 243, 1693–1710. [Google Scholar] [CrossRef] [Green Version]
- Stasiuk, M.; Kozubek, A. Biological activity of phenolic lipids. Cell. Mol. Life Sci. 2010, 67, 841–860. [Google Scholar] [CrossRef]
- Buonanno, F.; Quassinti, L.; Bramucci, M.; Amantini, C.; Lucciarini, R.; Santoni, G.; Iio, H.; Ortenzi, C. The protozoan toxin climacostol inhibits growth and induces apoptosis of human tumor cell lines. Chem. Biol. Interact. 2008, 176, 151–164. [Google Scholar] [CrossRef]
- Perrotta, C.; Buonanno, F.; Zecchini, S.; Giavazzi, A.; Proietti Serafini, F.; Catalani, E.; Guerra, L.; Belardinelli, M.C.; Picchietti, S.; Fausto, A.M.; et al. Climacostol reduces tumour progression in a mouse model of melanoma via the p53-dependent intrinsic apoptotic programme. Sci. Rep. 2016, 6, 27281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zecchini, S.; Proietti Serafini, F.; Catalani, E.; Giovarelli, M.; Coazzoli, M.; Di Renzo, I.; De Palma, C.; Perrotta, C.; Clementi, E.; Buonanno, F.; et al. Dysfunctional autophagy induced by the pro-apoptotic natural compound climacostol in tumour cells. Cell Death Dis. 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Buonanno, F.; Catalani, E.; Cervia, D.; Proietti Serafini, F.; Picchietti, S.; Fausto, A.M.; Giorgi, S.; Lupidi, G.; Rossi, F.V.; Marcantoni, E.; et al. Bioactivity and structural properties of novel synthetic analogues of the protozoan toxin climacostol. Toxins 2019, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Muto, Y.; Tanabe, Y.; Kawai, K.; Okano, Y.; Iio, H. Climacostol inhibits Tetrahymena motility and mitochondrial respiration. Cent. Eur. J. Biol. 2011, 6, 99–104. [Google Scholar] [CrossRef]
- Quassinti, L.; Ortenzi, F.; Marcantoni, E.; Ricciutelli, M.; Lupidi, G.; Ortenzi, C.; Buonanno, F.; Bramucci, M. DNA binding and oxidative DNA damage induced by climacostol–copper(II) complexes: Implications for anticancer properties. Chem. Biol. Interact. 2013, 206, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Krishnamurthy, S. Cellular responses to cisplatin-induced DNA damage. J. Nucl. Acids 2010, 201367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolaou, V.A.; Stratigos, A.J.; Flaherty, K.T.; Tsao, H. Melanoma: New insights and new therapies. J. Investig. Dermatol. 2012, 132, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gembeh, S.V.; Brown, R.L.; Grimm, C.; Cleveland, T.E. Identification of chemical components of corn kernel pericarp wax associated with resistance to Aspergillus flavus infection and aflatoxin production. J. Agric. Food Chem. 2001, 49, 4635–4641. [Google Scholar] [CrossRef]
- Zheng, L.-F.; Wei, Q.-Y.; Cai, Y.-J.; Fang, J.-G.; Zhou, B.; Yang, L.; Liu, Z.-L. DNA damage induced by resveratrol and its synthetic analogues in the presence of Cu(II) ions: Mechanism and structure-activity relationship. Free Radic. Biol. Med. 2006, 41, 1807–1816. [Google Scholar] [CrossRef]
- Catalani, E.; Buonanno, F.; Lupidi, G.; Bongiorni, S.; Belardi, R.; Zecchini, S.; Giovarelli, M.; Coazzoli, M.; De Palma, C.; Perrotta, C.; et al. The natural compound climacostol as a prodrug strategy based on pH activation for efficient delivery of cytotoxic small agents. Front. Chem. 2019, 7, 463–484. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [Green Version]
- Luìs, A.; Cruz, C.; Duarte, A.P.; Domingues, F. An alkenylresorcinol derivative from Hakea Sericea fruits and their antimicrobial activity. Nat. Prod. Commun. 2013, 8, 1459–1462. [Google Scholar] [CrossRef] [Green Version]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}