Reduce, Reuse and Recycle in Protein Chromatography: Development of an Affinity Adsorbent from Waste Paper and Its Application for the Purification of Proteases from Fish By-Products


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Extraction and Characterization of Cellulose Microfibers from Waste Paper
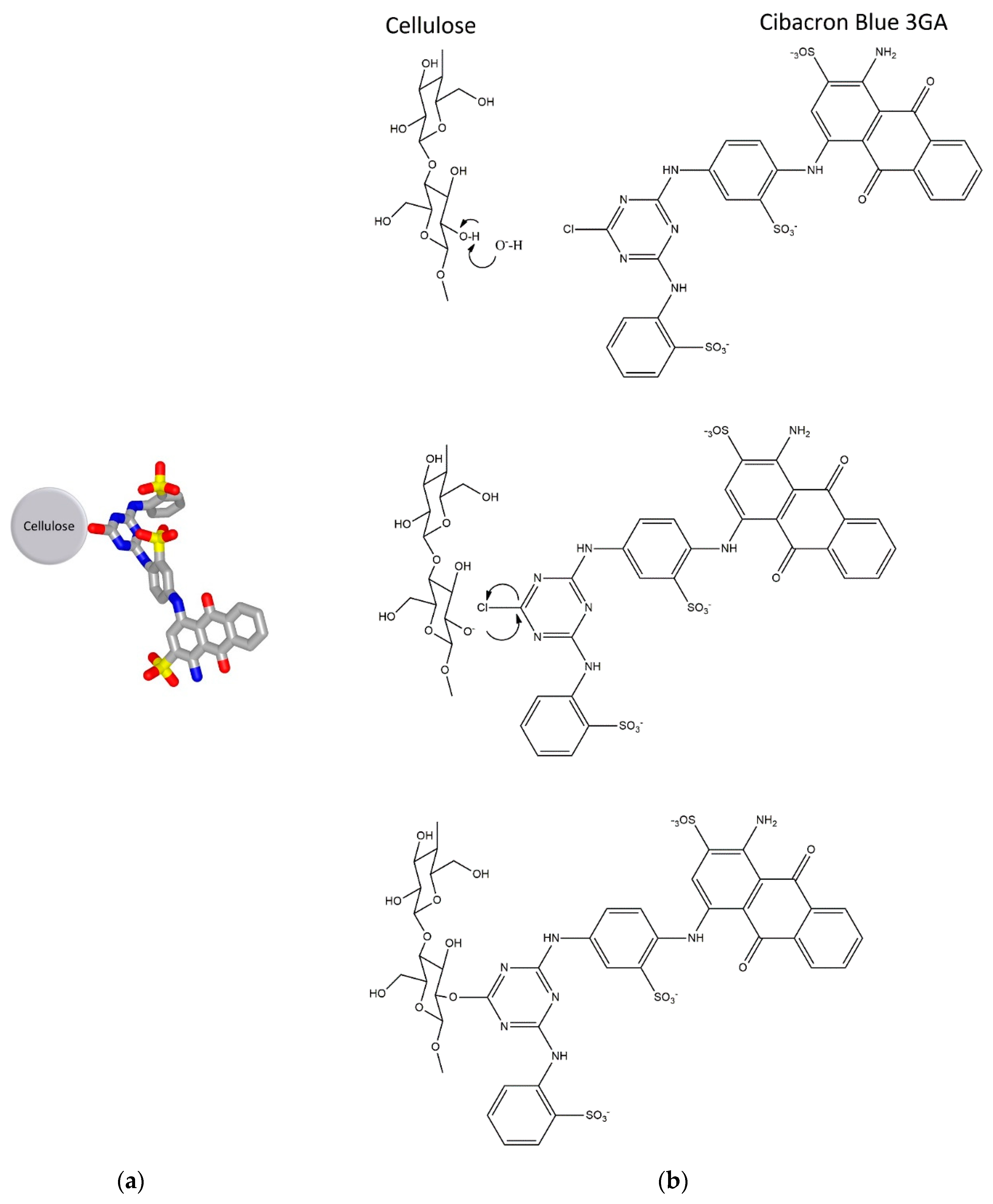
2.2.2. Immobilization of Cibacron Blue 3GA on Cellulose Microfibers
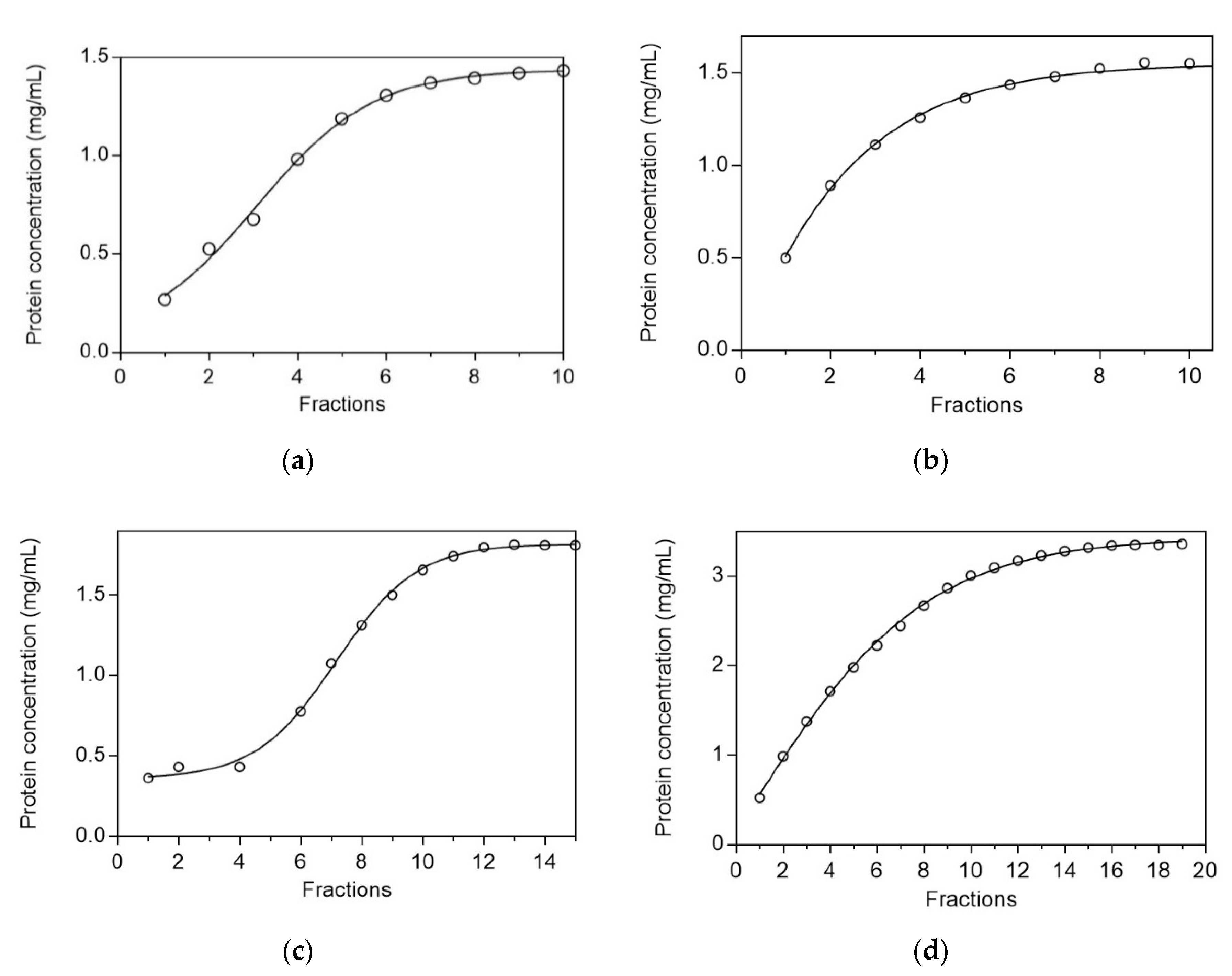
2.2.3. Determination of Protein Dynamic Capacity for the Affinity Adsorbents
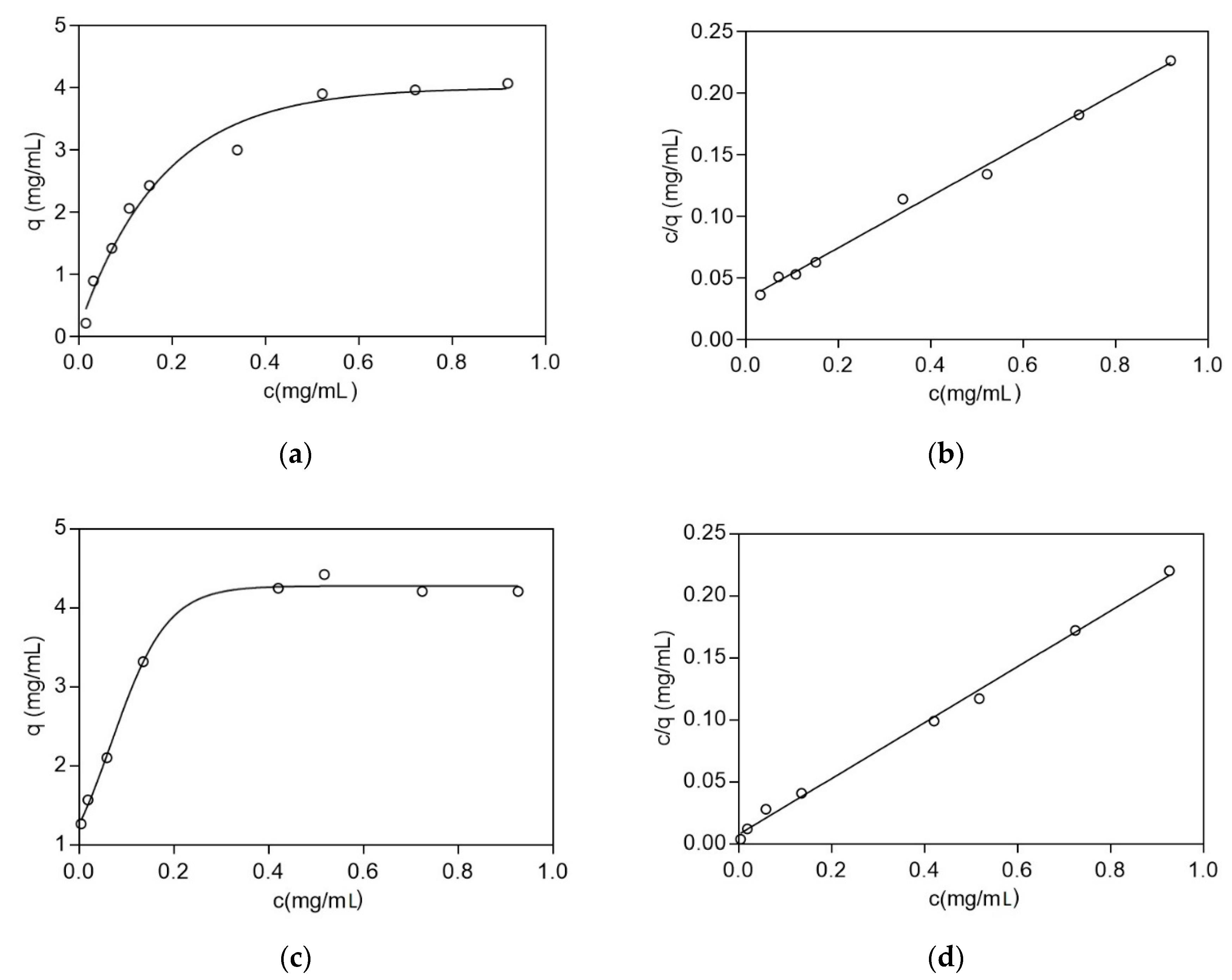
2.2.4. Absorption Equilibrium Studies
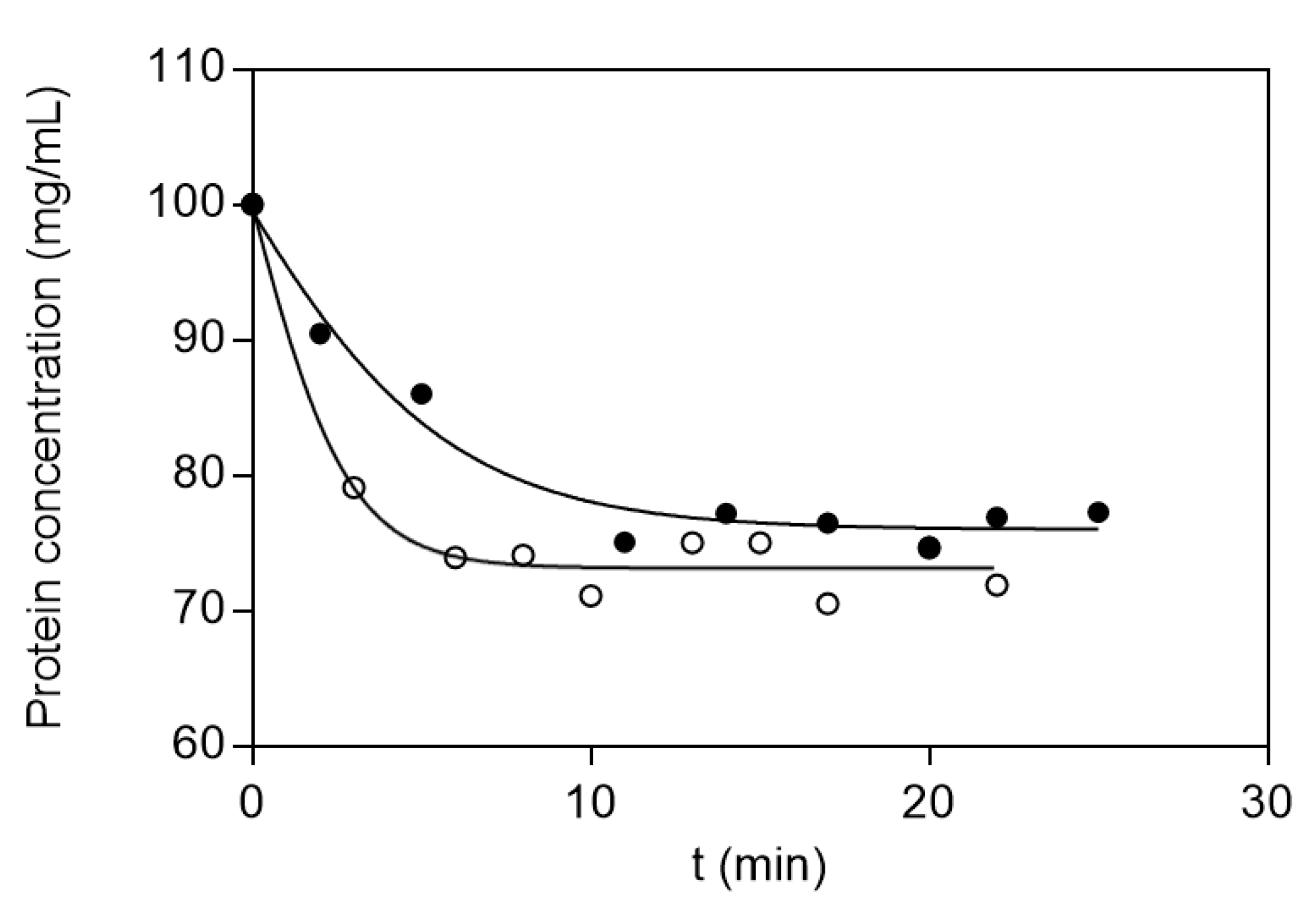
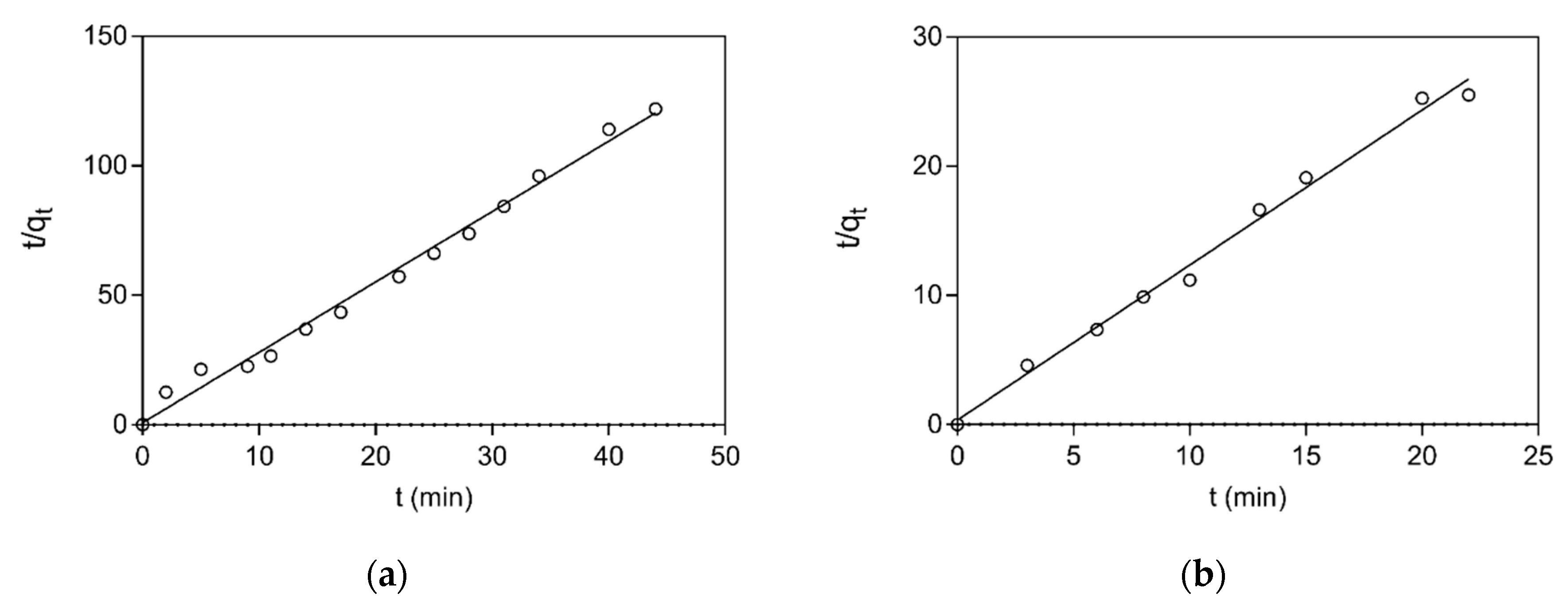
2.2.5. Absorption Kinetic Studies
2.2.6. Enzyme Assay
2.2.7. Extraction of Proteases from Sparus aurata Stomach and Pancreas
2.2.8. Affinity Chromatography of Proteases from Sparus aurata Stomach and Pancreas
3. Results and Discussion
3.1. Extraction and Characterization of Cellulose Microfibers from Waste Paper (Newspaper)
3.2. Synthesis of the Affinity Adsorbent
3.3. Adsorption Equilibrium and Kinetics Studies
3.4. Development and Optimization of the Purification Protocol of Proteases from Sparus aurata on the Adsorbent CB3GA-Cellulose-2
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Boto, R.E.F.; Anyanwu, U.; Sousa, F.; Almeida, P.; Queiroz, J.A. Thiacarbocyanine as ligand in dye-affinity chromatography for protein purification. II. Dynamic binding capacity using lysozyme as a model. Biomed. Chromatogr. 2009, 23, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Labrou, N.E. Design and selection of ligands for affinity chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 790, 67–78. [Google Scholar] [CrossRef]
- Labrou, N.E. Protein purifcation: An overview. In Methods in Molecular Biology; Clifon, N.J., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1129, pp. 3–10. [Google Scholar]
- Scopes, R.K. Overview of protein purification and characterization. Curr. Protoc. Protein Sci. 2001. [Google Scholar] [CrossRef]
- Halan, V.; Maity, S.; Bhambure, R.; Rathore, A.S. Multimodal Chromatography for Purification of Biotherapeutics—A Review. Curr. Protein. Pept. Sci. 2019, 20, 4–13. [Google Scholar] [CrossRef]
- Hanke, A.T.; Ottens, M. Purifying biopharmaceuticals: Knowledge-based chromatographic process development. Trends Biotechnol. 2014, 32, 210–220. [Google Scholar] [CrossRef]
- Insaidoo, F.K.; Rauscher, M.A.; Smithline, S.J.; Kaarsholm, N.C.; Feuston, B.P.; Ortigosa, A.D.; Linden, T.O.; Roush, D.J. Targeted purification development enabled by computational biophysical modeling. Biotechnol. Prog. 2015, 31, 154–164. [Google Scholar] [CrossRef]
- Clonis, Y.D. Affinity chromatography matures as bioinformatic and combinatorial tools develop. J. Chromatogr. A 2006, 1101, 1–24. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, G.; Lowe, C.R. A biomimetic Protein G affinity adsorbent: An Ugi ligand for immunoglobulins and Fab fragments based on the third IgG-binding domain of Protein, G.J. Mol. Recognit. 2013, 26, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Bhagavat, R.; Kim, H.B.; Kim, C.Y.; Terwilliger, T.C.; Mehta, D.; Srinivasan, N.; Chandra, N. A genome-wide structure-based survey of nucleotide binding proteins in M. tuberculosis. Sci. Rep. 2017, 7, 12489. [Google Scholar] [CrossRef] [PubMed]
- Amiri, S.; Mehrnia, M.R.; Roudsari, F.P. Enhancing purification efficiency of affinity functionalized composite agarose micro beads using Fe3O4 nanoparticles. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1041–1042, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Lee, S.Y.; Show, P.L.; Hong, S.C.; Chang, Y.K. Direct recovery of malate dehydrogenase from highly turbid yeast cell homogenate using dye-ligand affinity chromatography in stirred fluidized bed. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1100–1101, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ng, I.S.; Song, C.P.; Ooi, C.W.; Tey, B.T.; Lee, Y.H.; Chang, Y.K. Purification of lysozyme from chicken egg white using nanofiber membrane immobilized with Reactive Orange 4 dye. Int. J. Biol. Macromol. 2019, 34, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Başar, N.; Uzun, L.; Güner, A.; Denizli, A. Lysozyme purification with dye-affinity beads under magnetic field. Int. J. Biol. Macromol. 2007, 41, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Labrou, N.E.; Clonis, Y.D. Biomimetic dye affinity chromatography for the purification of bovine heart lactate dehydrogenase. J. Chromatogr. A 1995, 718, 35–44. [Google Scholar] [CrossRef]
- Chronopoulou, E.; Labrou, N.E. Synthesis and Application of Dye-Ligand Affinity Adsorbents. In Protein Downstream Processing. Methods in Molecular Biology (Methods and Protocols); Labrou, N., Ed.; Humana Press: Totowa, NJ, USA, 2014; pp. 263–276. [Google Scholar]
- Manoukian, O.S.; Sardashti, N.; Stedman, T.; Gailiunas, K.; Ojha, A.; Penalosa, A.; Kumbar, S.G. Biomaterials for Tissue Engineering and Regenerative Medicine. Encycl. Biomed. Eng. 2019, 462–482. [Google Scholar]
- Clonis, Y.D.; Labrou, N.E.; Kotsira, V.P.; Mazitsos, C.; Melissis, S.; Gogolas, G. Biomimetic dyes as affinity chromatography tools in enzyme purification. J. Chromatogr. A 2000, 891, 33–44. [Google Scholar] [CrossRef]
- Denizli, A.; Pişkin, E. Dye-ligand affinity systems. J. Biochem. Bioph. Meth. 2001, 49, 391–416. [Google Scholar] [CrossRef]
- Li, Z.; Cao, M.; Zhang, W.; Liu, L.; Wang, J.; Ge, W.; Yuan, Y.; Yue, T.; Li, R.; Yu, W.W. Affinity adsorption of lysozyme with Reactive Red 120 modified magnetic chitosan microspheres. Food Chem. 2014, 145, 749–755. [Google Scholar] [CrossRef]
- Andaç, M. Cibacron blue immobilized poly(glycidyl-methacrylate) nanobeads for albumin removal in proteome studies. Artif. Cells Nanomed. Biotechnol. 2015, 43, 133–139. [Google Scholar] [CrossRef]
- Graça, V.C.; Sousa, F.; Santos, P.F.; Almeida, P.S. Protein purification by aminosquarylium cyanine dye-affinity chromatography. Methods Mol. Biol. 2015, 1286, 23–33. [Google Scholar]
- Kim, S.K.; Dewapriya, P. Enzymes from Fish Processing Waste Materials and Their Commercial Applications. In Seafood Processing By-Products; Springer: New York, NY, USA, 2013; pp. 183–196. [Google Scholar]
- Patel, S. A critical review on serine protease: Key immune manipulator and pathology mediator. Allergol. Immunopathol. (Madr) 2017, 45, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Haard, N.F.; Simpson, B.K. Proteases from Aquatic Organisms and Their Uses in the Seafood Industry. Fisheries Processing; Springer: Boston, MA, USA, 1994; pp. 132–154. [Google Scholar]
- Pant, D.; Sharma, V.; Singh, P.; Kumar, M.; Giri, A.; Singh, M.P. Perturbations and 3R in carbon management. Env. Sci. Pollut. Res. Int. 2017, 24, 4413–4432. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Nakagaito, A.N.; Bistamam, M.S.A. Extraction of cellulose nanofiber from waste papers and application to reinforcement in biodegradable composites. J. Reinf. Plast. Comp. 2013, 32, 1542–1546. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Livingston, A.; Chase, H. Preparation and Characterization of Adsorbents for Use in High Performance Liquid Affinity Chromatography. J. Chromatogr. 1989, 481, 159–174. [Google Scholar] [CrossRef]
- Dalakouras, T.; Smith, B.J.; Platis, D.; Cox, M.M.J.; Labrou, N.E. Development of recombinant protein-based influenza vaccine. Expression and affinity purification of H1N1 influenza virus neuraminidase. J. Chromatogr. A 2006, 1136, 48–56. [Google Scholar] [CrossRef]
- Platis, D.; Labrou, N.E. Affinity chromatography for the purification of therapeutic proteins from transgenic maize using immobilized histamine. J. Sep. Sci. 2008, 31, 636–645. [Google Scholar] [CrossRef]
- Marinou, M.; Platis, D.; Ataya, F.S.; Chronopoulou, E.; Vlachakis, D.; Labrou, N.E. Structure-based design and application of a nucleotide coenzyme mimetic ligand: Application to the affinity purification of nucleotide dependent enzymes. J. Chromatogr. A 2018, 1535, 88–100. [Google Scholar] [CrossRef]
- Gerente, C.; McKay, G.; Andres, Y.; Le Cloirec, P. Interactions of Natural Aminated Polymers with Different Species of Arsenic at Low Concentrations: Application in Water Treatment. Adsorption 2005, 11, 859–863. [Google Scholar] [CrossRef]
- Ho, Y.S.; Mckay, G. Pseudo-second order model for sorption process. Process. Biochem. 1999, 34, 451–465. [Google Scholar] [CrossRef]
- Azizian, S. Kinetic models of sorption: A theoretical analysis. J. Colloid. Interface Sci. 2004, 276, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.J. Biosorption isotherms, kinetics and thermodynamics. Sep. Purif. 2008, 61, 229–242. [Google Scholar] [CrossRef]
- Côlho, D.F.; Saturnino, T.P.; Fernandes, F.F.; Mazzola, P.G.; Silveira, E.; Tambourgi, E.B. Azocasein Substrate for Determination of Proteolytic Activity: Reexamining a Traditional Method Using Bromelain Samples. Biomed. Res. Int. 2016, 8, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bergmeyer, H.U. Methods of Enzymatic Analysis; Elsevier: New York, NY, USA, 1974; Volume 2. [Google Scholar]
- Heath, L.; Thielemans, W. Cellulose nanowhisker aerogels. Green Chem. 2010, 12, 1448–1453. [Google Scholar] [CrossRef]
- Souza, A.G.; de Kano, F.S.; Bonvent, J.J.; Rosa, D.d.S. Cellulose Nanostructures Obtained from Waste Paper Industry: A Comparison of Acid and Mechanical Isolation Methods. Mater. Res. 2017, 20, 209–214. [Google Scholar] [CrossRef]
- Danial, W.H.; Abdul Majid, Z.; Mohd Muhid, M.N.; Triwahyono, S.; Bakar, M.B.; Ramli, Z. The reuse of wastepaper for the extraction of cellulose nanocrystals. Carbohyd. Polym. 2015, 118, 165–169. [Google Scholar] [CrossRef]
- Vanderghem, C.; Jacquet, N.; Danthine, S.; Blecker, C.; Paquot, M. Effect of Physicochemical Characteristics of Cellulosic Substrates on Enzymatic Hydrolysis by Means of a Multi-Stage Process for Cellobiose Production. Appl. Biochem. Biotechnol. 2012, 166, 1423–1432. [Google Scholar] [CrossRef]
- Labrou, N.E.; Eliopoulos, E.; Clonis, Y.D. Dye-affinity labelling of bovine heart mitochondrial malate dehydrogenase and study of the NADH-binding site. Biochem. J. 1996, 315, 687–693. [Google Scholar] [CrossRef]
- Labrou, N.E.; Eliopoulos, E.; Clonis, Y.D. Molecular modelling for the design of chimaeric biomimetic dye-ligands and their interaction with bovine heart mitochondrial malate dehydrogenase. Biochem. J. 1996, 315, 695–703. [Google Scholar] [CrossRef]
- Boyer, P.M.; Hsu, J.T. Effects of ligand concentration on protein adsorption in dye-ligand adsorbents. Chem. Eng. Sci. 1992, 47, 241–251. [Google Scholar] [CrossRef]
- Lindner, N.M.; Jeffcoat, R.; Lowe, C.R. Design and applications of biomimetic anthraquinone dyes. J. Chromatogr. A 1989, 473, 227–240. [Google Scholar] [CrossRef]
- Burton, J.; Division, O.; Street, D.; Vivian, C.; Lowe, R.; Street, D. Design and application of biomimetic dyes II: The interaction of C.I. Reactive Blue 2 analogues bearing terminal ring modifications with horse liver alcohol dehydrogenase. J. Chromatogr. A 1988, 455, 201–216. [Google Scholar] [CrossRef]
- Labrou, N.E.; Clonis, Y.D. Biomimetic-dye affinity chromatography for the purification of mitochondrial L-malate dehydrogenase from bovine heart. J. Biotechnol. 1996, 45, 185–194. [Google Scholar] [CrossRef]
- Pabst, T.M.; Thai, J.; Hunter, A.K. Evaluation of Recent Protein A Stationary Phase Innovations for Capture of Biotherapeutics. J. Chromatogr. A 2018, 1554, 45–60. [Google Scholar] [CrossRef]
- Jiwalak, N.; Rattanaphani, S.; Bremner, J.B.; Rattanaphani, V. Equilibrium and kinetic modeling of the adsorption of indigo carmine onto silk. Fiber Polym. 2010, 11, 572–579. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, G.C.; Garcia Rojas, E.E.; Coelho Junior, M.S.; Coimbra, J.S.R. Adsorption kinetics and thermodynamic parameters of egg white proteins. Eur. Food Res. Technol. 2011, 232, 985–993. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Dissociation Constants (KD) (mg/mL) | Maximum Adsorption Capacity (qmax) (mg/mL) |
|---|---|---|
| BSA | 0.16 | 4.78 |
| LYS | 0.03 | 4.43 |
| Protein | k2 (mg Absorbent/mg Protein min) | qe (mg Protein/mg) | h (mg Protein/mg min) | R2 |
|---|---|---|---|---|
| BSA | 0.76 | 0.39 | 5.64 | 0.990 |
| LYS | 0.35 | 0.90 | 0.51 | 0.992 |
| pH of the Equilibration Buffer (Binding) | pH of the Equilibration Buffer (Elution) | Purifying Ability (Purification Fold) | Capacity (%) * | Yield (%) |
|---|---|---|---|---|
| 7.0 | 7.0 | 9 | 25 | 8.5 |
| 6.5 | 6.5 | 11 | 25 | 46.7 |
| 5.5 | 6.5 | 47.8 | 100 | 167.8 |
| Step | Activity (Units) | Protein (mg) | Specific Activity (U/mg) | Purification (fold) | Yield (%) * |
|---|---|---|---|---|---|
| Protease from stomach extract | |||||
| Crude extract | 7.07 | 10.51 | 0.67 | 1 | 100 |
| Affinity chromatography | 11.87 | 0.37 | 32.08 | 47.88 | 167.89 |
| Protease from pancreatic extract | |||||
| Crude extract | 7.87 | 1.81 | 4.35 | 1 | 100 |
| Affinity chromatography | 8.92 | 0.06 | 148.67 | 34.18 | 113.34 |
| Trypsin | |||||
| Crude extract | 960.32 | 1.81 | 530.56 | 1 | 100 |
| Affinity chromatography | 471.06 | 0.06 | 7851.00 | 14.80 | 49.05 |
| Chymotrypsin | |||||
| Crude extract | 4.56 | 1.81 | 2.52 | 1 | 100 |
| Affinity chromatography | 0.86 | 0.06 | 14.33 | 5.69 | 18.86 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Premetis, G.E.; Labrou, N.E. Reduce, Reuse and Recycle in Protein Chromatography: Development of an Affinity Adsorbent from Waste Paper and Its Application for the Purification of Proteases from Fish By-Products. Biomolecules 2020, 10, 822. https://doi.org/10.3390/biom10060822
Premetis GE, Labrou NE. Reduce, Reuse and Recycle in Protein Chromatography: Development of an Affinity Adsorbent from Waste Paper and Its Application for the Purification of Proteases from Fish By-Products. Biomolecules. 2020; 10(6):822. https://doi.org/10.3390/biom10060822
Chicago/Turabian StylePremetis, Georgios E., and Nikolaos E. Labrou. 2020. "Reduce, Reuse and Recycle in Protein Chromatography: Development of an Affinity Adsorbent from Waste Paper and Its Application for the Purification of Proteases from Fish By-Products" Biomolecules 10, no. 6: 822. https://doi.org/10.3390/biom10060822