Discovery of a Novel Mutation in DNA Gyrase and Changes in the Fluoroquinolone Resistance of Helicobacter pylori over a 14-Year Period: A Single Center Study in Korea

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Patient Demographics and H. pylori Strains
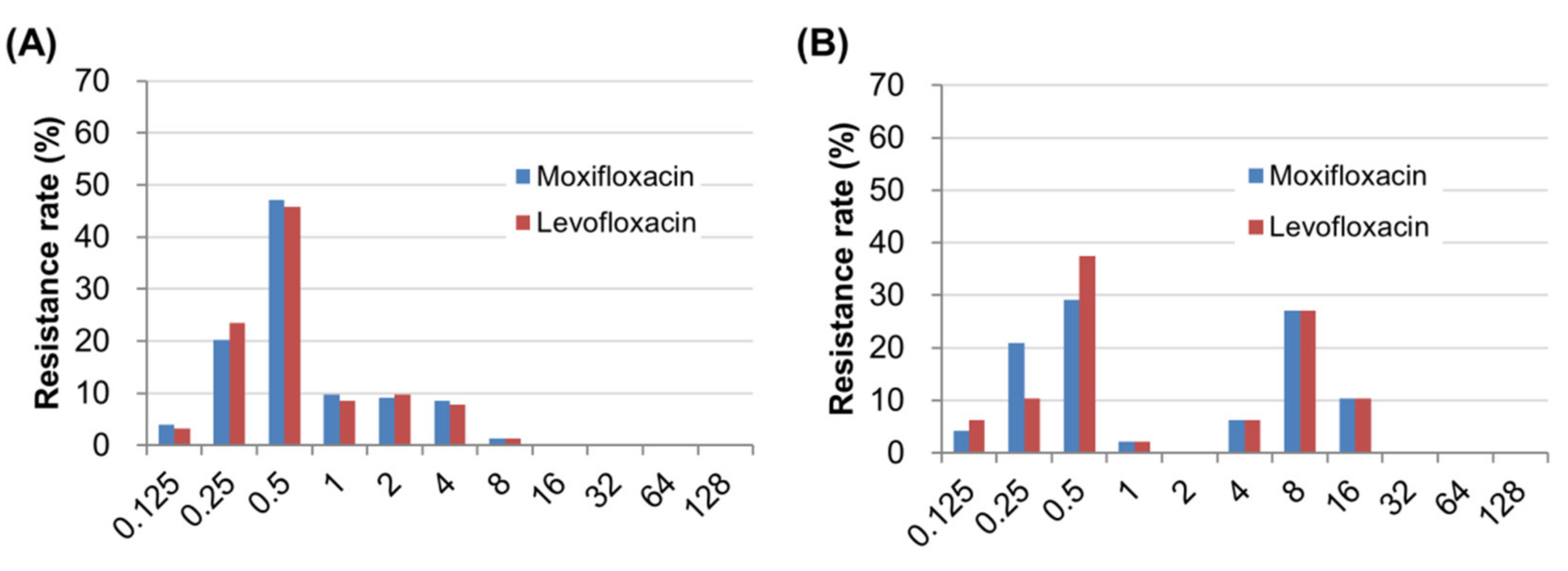
2.2. Prevalence of Fluoroquinolone Resistance and Distribution of Minimum Inhibitory Concentrations (MICs)
2.3. The Mutation Analyses of DNA Gyrase Genes and the Association with MIC Levels for Fluoroquinolones
2.4. Transformation of H. pylori and Confirmation of Gyrase Mutations Related to Fluoroquinolone Resistance
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. H. pylori Culture and Isolation
4.3. PCR-Based Randomly Amplified Polymorphic DNA (RAPD) Fingerprinting of H. pylori
4.4. Determination of MICs of Fluoroquinolones
4.5. PCR Amplification and Nucleotide Sequencing
4.6. Natural Transformation of H. pylori
4.7. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- NIH Consensus Conference. Helicobacter pylori in peptic ulcer disease. NIH Consensus Development Panel on Helicobacter pylori in Peptic Ulcer Disease. JAMA 1994, 272, 65–69. [Google Scholar] [CrossRef]
- Nahar, S.; Mukhopadhyay, A.K.; Khan, R.; Ahmad, M.M.; Datta, S.; Chattopadhyay, S.; Dhar, S.C.; Alam Sarker, S.; Engstrand, L.; Berg, U.E.; et al. Antimicrobial Susceptibility of Helicobacter pylori Strains Isolated in Bangladesh. J. Clin. Microbiol. 2004, 42, 4856–4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selgrad, M.; Meile, J.; Bornschein, J.; Kandulski, A.; Langner, C.; Varbanova, M.; Wex, T.; Tammer, I.; Schlüter, D.; Malfertheiner, P. Antibiotic susceptibility of Helicobacter pylori in central Germany and its relationship with the number of eradication therapies. Eur. J. Gastroenterol. Hepatol. 2013, 25, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Fischbach, L. Helicobacter pylori treatment in the era of increasing antibiotic resistance. Gut 2010, 59, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Mégraud, F.; Coenen, S.; Versporten, A.; Kist, M.; Lopez-Brea, M.; Hirschl, A.M.; Andersen, L.P.; Goossens, H.; Glupczynski, Y. On behalf of the Study Group participants Helicobacter pylori resistance to antibiotics in Europe and its relationship to antibiotic consumption. Gut 2012, 62, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Megraud, F.; A O’Morain, C.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection—The Maastricht V/Florence Consensus Report. Gut 2016, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Gatta, L.; Zullo, A.; Perna, F.; Ricci, C.; De Francesco, V.; Tampieri, A.; Bernabucci, V.; Cavina, M.; Hassan, C.; Ierardi, E.; et al. A 10-day levofloxacin-based triple therapy in patients who have failed two eradication courses. Aliment. Pharmacol. Ther. 2005, 22, 45–49. [Google Scholar] [CrossRef]
- Tai, W.-C.; Lee, C.-H.; Chiou, S.-S.; Kuo, C.-M.; Liang, C.-M.; Lu, L.-S.; Chiu, C.-H.; Wu, K.-L.; Chiu, Y.-C.; Hu, T.-H.; et al. The Clinical and Bacteriological Factors for Optimal Levofloxacin-Containing Triple Therapy in Second-Line Helicobacter pylori Eradication. PLoS ONE 2014, 9, e105822. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.C.; Jacoby, G.A. Mechanisms of drug resistance: Quinolone resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 12–31. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, T.; Suzuki, H.; Hibi, T. Quinolone-Based Third-Line Therapy for Helicobacter pylori Eradication. J. Clin. Biochem. Nutr. 2009, 44, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. Review article: The global emergence of Helicobacter pylori antibiotic resistance. Aliment. Pharmacol. Ther. 2015, 43, 514–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaka, H.; Rüttgerodt, N.; Bohne, W.; Mueller, A.; Gross, U.; Kasang, C.; Mshana, S. Helicobacter pylori Mutations Conferring Resistance to Fluoroquinolones and Clarithromycin among Dyspeptic Patients Attending a Tertiary Hospital, Tanzania. Can. J. Gastroenterol. Hepatol. 2019, 2019, 8481375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, E.J.; Yun, S.-C.; Jung, H.-Y.; Lim, H.; Choi, K.-S.; Ahn, J.Y.; Lee, J.H.; Kim, D.H.; Choi, K.D.; Song, H.J.; et al. Meta-Analysis of First-Line Triple Therapy for Helicobacter pylori Eradication in Korea: Is It Time to Change? J. Korean Med. Sci. 2014, 29, 704–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.G.; Kim, J.J.; Lee, Y.C.; Choi, I.J.; Yoon, S.M.; Sung, J.K.; Kim, S.W.; Kim, H.S.; Jeon, S.W.; Lee, J.Y.; et al. Su1283 – Ten-Day Concomitant, 10-Day Sequential, and 7-Day Triple Therapy in First-Line Treatment of Helicobacter Pylori Infection: A Nationwide Randomized Trial in Korea. Gastroenterology 2019, 156. [Google Scholar] [CrossRef]
- A Fallone, C.; Moss, S.F.; Malfertheiner, P. Reconciliation of Recent Helicobacter pylori Treatment Guidelines in a Time of Increasing Resistance to Antibiotics. Gastroenterology 2019, 157, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hong, S.P.; Kwon, C.I.; Phyun, L.H.; Lee, B.S.; Song, H.U.; Ko, K.H.; Hwang, S.-G.; Park, P.W.; Rim, K.S.; et al. The efficacy of levofloxacin based triple therapy for Helicobacter pylori eradication. Korean J. Gastroenterol. 2006, 48, 19–24. [Google Scholar] [PubMed]
- Choi, K.H.; Chung, W.C.; Lee, K.-M.; Paik, C.N.; Kim, E.J.; Kang, B.K.; Oak, J.H.; Jung, S.H. Efficacy of Levofloxacin and Rifaximin based Quadruple Therapy in Helicobacter pylori Associated Gastroduodenal Disease: A Double-Blind, Randomized Controlled Trial. J. Korean Med. Sci. 2011, 26, 785–790. [Google Scholar] [CrossRef]
- Lim, J.H.; Kim, S.G.; Song, J.H.; Hwang, J.J.; Lee, N.H.; Han, J.P.; Hong, S.J.; Kim, J.H.; Jeon, S.W.; Kim, G.H.; et al. Efficacy of Levofloxacin-Based Third-Line Therapy for the Eradication of Helicobacter pylori in Peptic Ulcer Disease. Gut Liver 2016, 11, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Di Caro, S.; Fini, L.; Daoud, Y.; Grizzi, F.; Gasbarrini, A.; De Lorenzo, A.; Di Renzo, L.; McCartney, S.; Bloom, S. Levofloxacin/amoxicillin-based schemes vs quadruple therapy for Helicobacter pylori eradication in second-line. World J. Gastroenterol. 2012, 18, 5669–5678. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Ahn, J.Y.; Choi, K.D.; Jung, H.-Y.; Kim, J.M.; Baik, G.H.; Kim, B.; Park, J.C.; Jung, H.; Cho, S.J.; et al. Nationwide antibiotic resistance mapping of Helicobacter pylori in Korea: A prospective multicenter study. Helicobacter 2019, 24, e12592. [Google Scholar] [CrossRef]
- Carothers, J.J.; Bruce, M.G.; Hennessy, T.; Bensler, M.; Morris, J.M.; Reasonover, A.L.; Hurlburt, D.A.; Parkinson, A.J.; Coleman, J.M.; McMahon, B. The Relationship between Previous Fluoroquinolone Use and Levofloxacin Resistance in Helicobacter pylori Infection. Clin. Infect. Dis. 2007, 44, e5–e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Jung, H.C.; Song, I.S. Distribution of fluoroquinolone MICs in Helicobacter pylori strains from Korean patients. J. Antimicrob. Chemother. 2005, 56, 965–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tankovic, J.; Lascols, C.; Sculo, Q.; Petit, J.-C.; Soussy, C.-J. Single and Double Mutations in gyrA but Not in gyrB Are Associated with Low- and High-Level Fluoroquinolone Resistance in Helicobacter pylori. Antimicrob. Agents Chemother. 2003, 47, 3942–3944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Kim, N.; Nam, R.H.; Kim, J.M.; Park, J.Y.; Lee, S.M.; Kim, J.S.; Lee, D.H.; Jung, H.C. High Efficacy of Finafloxacin on Helicobacter pylori Isolates at pH 5.0 Compared with That of other Fluoroquinolones. Antimicrob. Agents Chemother. 2015, 59, 7629–7636. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.A.; Beckthold, B.; Wong, S.; Kureishi, A.; Bryan, L.E. Nucleotide sequence of the gyrA gene and characterization of ciprofloxacin-resistant mutants of Helicobacter pylori . Antimicrob. Agents Chemother. 1995, 39, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.-W.; Lee, G.H.; Jeong, J.-Y.; Lee, S.M.; Jung, J.H.; Choi, K.D.; Song, H.J.; Jung, H.-Y.; Kim, J.-H. Resistance of Helicobacter pylori strains to antibiotics in Korea with a focus on fluoroquinolone resistance. J. Gastroenterol. Hepatol. 2012, 27, 493–497. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, N.; Nam, R.H.; Park, J.H.; Kim, J.M.; Jung, H.C.; Song, I.S. Mutations of Helicobacter pylori Associated with Fluoroquinolone Resistance in Korea. Helicobacter 2011, 16, 301–310. [Google Scholar] [CrossRef]
- Miyachi, H.; Aoyama, N.; Shirasaka, D.; Miki, I.; Matsumoto, Y.; Azuma, T. Primary Levofloxacin-Resistance and gyrA/B Mutations among Helicobacter pylori in Japan. Am. J. Gastroenterol. 2006, 11, 9–243. [Google Scholar] [CrossRef]
- Saeed, A.; Bosch, A.; Bettiol, M.; González, D.N.; Erben, M.F.; Lamberti, Y. Novel Guanidine Compound against Multidrug-Resistant Cystic Fibrosis-Associated Bacterial Species. Molecules 2018, 23, 1158. [Google Scholar] [CrossRef] [Green Version]
- Bizzarri, B.; Al, E.A.E. Synthesis and in vitro Selective anti-Helicobacter pylori Activity of N-Substituted-2-oxo-2H-1-benzopyran-3-carboxamides. Eur. J. Med. Chem. 2006, 37, 208–212. [Google Scholar] [CrossRef]
- Akopyanz, N.; Bukanov, N.O.; Westblom, T.; Kresovich, S.; Berg, U.E. DNA diversity among clinical isolates of Helicobacter pylori detected by PCR-based RAPD fingerprinting. Nucleic Acids Res. 1992, 20, 5137–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.C.; Lee, S.-Y.; Pyo, J.H.; Kwon, D.H.; Rhee, J.C.; Kim, J. Isogenic Variation of Helicobacter pylori Strain Resulting in Heteroresistant Antibacterial Phenotypes in a Single Host In Vivo. Helicobacter 2005, 10, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kim, J.S.; Jung, H.C.; Kim, N.; Kim, Y.-J.; Song, I.S. Distribution of Antibiotic MICs for Helicobacter pylori Strains over a 16-Year Period in Patients from Seoul, South Korea. Antimicrob. Agents Chemother. 2004, 48, 4843–4847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Committee for Clinical Laboratory Standards (NCCLS). Performance Standards for Antimicrobial Susceptibility Testing: Twelfth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2002. [Google Scholar]
- Kim, S.J.; Kim, J.G.; Jung, K.; Hong, Y.H.; Kim, J.H.; Jung, H.R.; Kwon, J.H.; Yang, Y.H.; Kim, H.J.; Do, J.H.; et al. Antimicrobial resistance rate of Helicobacter pylori isolates and detection of mechanism of clarithromycin resistance. Korean J. Med. 2001, 61, 470–479. [Google Scholar]


{kind=link}
| Characteristics | 2005/2006 | 2017/2018 | p-Value |
|---|---|---|---|
| Number of patients (strains) | 143 (153) | 48 (48) | - |
| Mean age ± SD (years) | 48.6 ± 14.6 | 61.9 ± 8.7 | <0.001 |
| Sex (M/F) | 87/56 (60.8/39.2%) | 33/15 (68.8/31.2%) | 0.326 |
| Endoscopic diagnosis | <0.001 | ||
| Peptic ulcer disease | 65 (45.5%) | 15 (31.3%) | |
| Gastric cancer | 3 (2.1%) | 11 (22.9%) | |
| Gastric adenoma | 0 (0.0%) | 4 (8.3%) | |
| MALT lymphoma | 0 (0.0%) | 1 (2.1%) | |
| Gastritis | 75 (52.4%) | 17 (35.4%) |
| Prevalence of Resistant Strains (Resistant Strains/Total Strains, %) | |||
|---|---|---|---|
| 2005/2006 | 2017/2018 | p-Value | |
| Levofloxacin | 19.0% (29/153) | 43.8% (21/48) | <0.001 |
| Moxifloxacin | 19.0% (29/153) | 43.8% (21/48) | <0.001 |
| Group | Substitution in gyrA | Substitution in gyrB | Number of Patients | MIC (µg/mL) Value | |||
|---|---|---|---|---|---|---|---|
| LVX | MOX | ||||||
| Mean | p-Value * | Mean | p-Value * | ||||
| 2005/2006 (n = 29) | Asn-87→Lys | Asp-495→His | 3 (10.3%) | 4 | 0.771 | 4 | 0.823 |
| Asn-87→Lys | ND | 12 (41.4%) | 3.5 | 3.3 | |||
| Asp-91→Gly | ND | 12 (41.4%) | 3 | 3 | |||
| ND | ND | 2 (6.9%) | 3 | 3 | |||
| 2017/2018 (n = 21) | Asn-87→Lys | ND | 13 (62.0%) | 10.5 | 0.652 | 10.5 | 0.652 |
| Asp-91→Asn or Gly or Tyr | ND | 5 (23.8%) | 7.2 | 7.2 | |||
| Ala-88→Val | ND | 1 (4.8%) | 8 | 8 | |||
| Asn-87→Lys& Ala-88→Val | ND | 1 (4.8%) | 8 | 8 | |||
| Gly-85→Cys | ND | 1 (4.8%) | 8 | 8 | |||
| Isolated Strains | Amplified PCR Product Used in Transformation | Mutation in Transformed DNA | Transformation Frequency (MIC > 1 µg/mL) | Result | |
|---|---|---|---|---|---|
| First | Second | ||||
| Negative control Positive control QA2 QA1 QA4 QA12 | gyrB gyrA gyrB gyrB gyrB gyrB | None Asn87→Lys None Asp495→His Asp495→His Asp495→His | 2.8 × 10−9 5.7 × 10−6 3.0 × 10−9 3.4 × 10−9 2.9 × 10−9 2.5 × 10−9 | 2.5 × 10−9 4.1 × 10−6 4.9 × 10−9 3.1 × 10−9 3.4 × 10−9 1.9 × 10−9 | 2.65 × 10−9 4.9 × 10−6 3.95 × 10−9 3.25 × 10−9 3.15 × 10−9 2.2 × 10−9 |
| Donor Strain | MIC of Donor Strain (μg/mL) | gyrA Mutation of Donor Strain | MIC of Transformed Strains (μg/mL) | gyrA Mutation Detected in Transformed Strains | ||
|---|---|---|---|---|---|---|
| LVX | MOX | LVX | MOX | |||
| QB1 | 8 | 8 | Gly-85→Cys | 0.5→2 | 0.25→2 | Gly-85→Cys |
| QB16 | 8 | 8 | Ala-88→Val | 0.5→0.5 | 0.25→0.25 | Ala-88→Val |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhie, S.Y.; Park, J.Y.; Shin, T.-S.; Kim, J.W.; Kim, B.J.; Kim, J.G. Discovery of a Novel Mutation in DNA Gyrase and Changes in the Fluoroquinolone Resistance of Helicobacter pylori over a 14-Year Period: A Single Center Study in Korea. Antibiotics 2020, 9, 287. https://doi.org/10.3390/antibiotics9060287
Rhie SY, Park JY, Shin T-S, Kim JW, Kim BJ, Kim JG. Discovery of a Novel Mutation in DNA Gyrase and Changes in the Fluoroquinolone Resistance of Helicobacter pylori over a 14-Year Period: A Single Center Study in Korea. Antibiotics. 2020; 9(6):287. https://doi.org/10.3390/antibiotics9060287
Chicago/Turabian StyleRhie, Su Yeon, Jae Yong Park, Tae-Seop Shin, Jeong Wook Kim, Beom Jin Kim, and Jae Gyu Kim. 2020. "Discovery of a Novel Mutation in DNA Gyrase and Changes in the Fluoroquinolone Resistance of Helicobacter pylori over a 14-Year Period: A Single Center Study in Korea" Antibiotics 9, no. 6: 287. https://doi.org/10.3390/antibiotics9060287