Regulatory Potential of bHLH-Type Transcription Factors on the Road to Rubber Biosynthesis in Hevea brasiliensis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Suspension-Cultured Cells, Treatment with MeJA or DMSO
2.2. RNA Extraction, RNA-Seq and the Analysis
2.3. Search for TF Genes Highly Expressed in Latex
2.4. Phylogenetic Tree and Domain Search
2.5. Construction of Plasmids and In Vitro Protein Synthesis for gDB-seq
2.6. SDS-PAGE
2.7. gDB-Seq Analysis
2.8. Western Blot Analysis
2.9. Data Deposition
3. Results
3.1. Time-Course RNA-Seq Analysis in Cells Treated with MeJA
3.2. Characterization of the DEGs
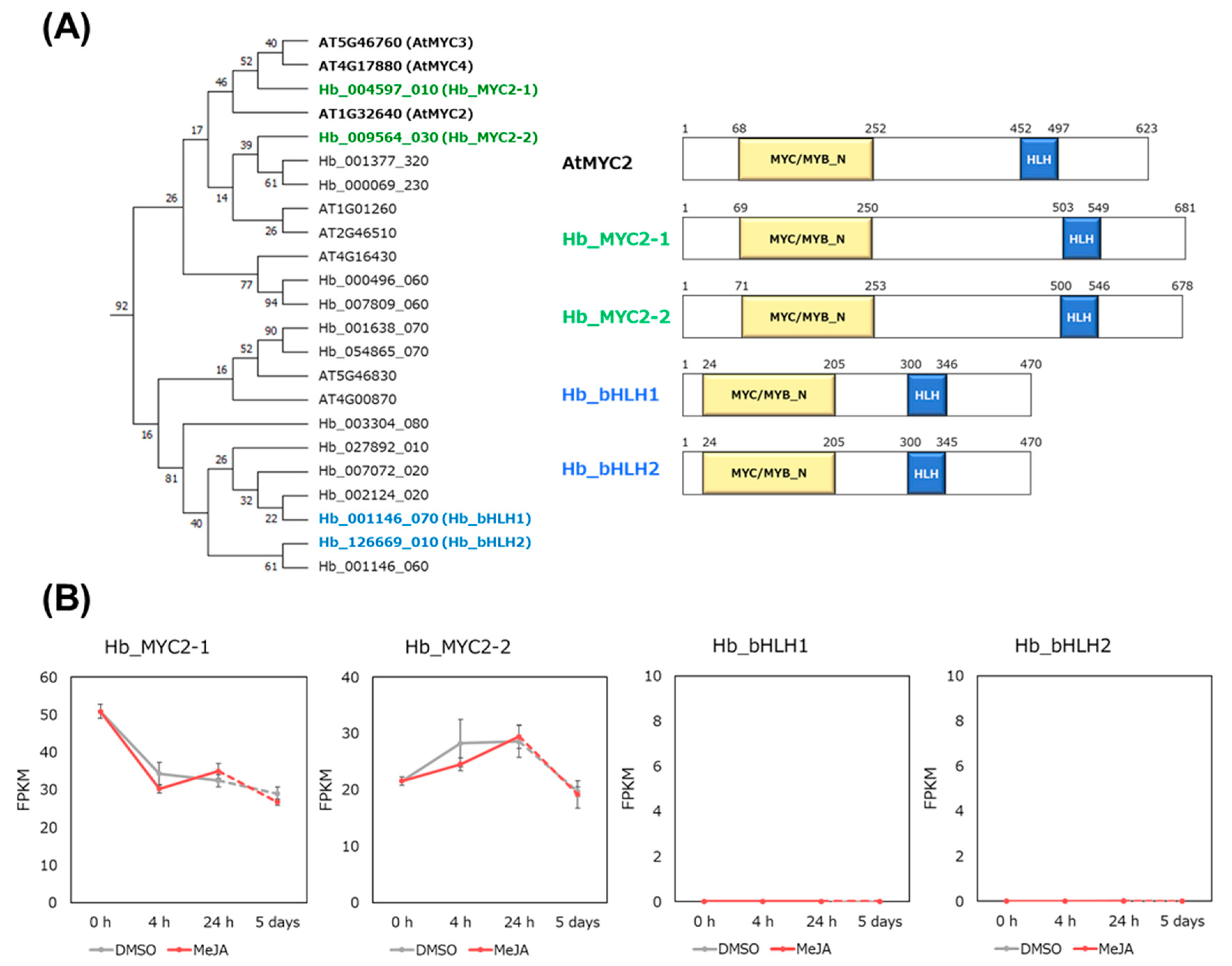
3.3. Hb_MYC2 and the Related bHLH TFs
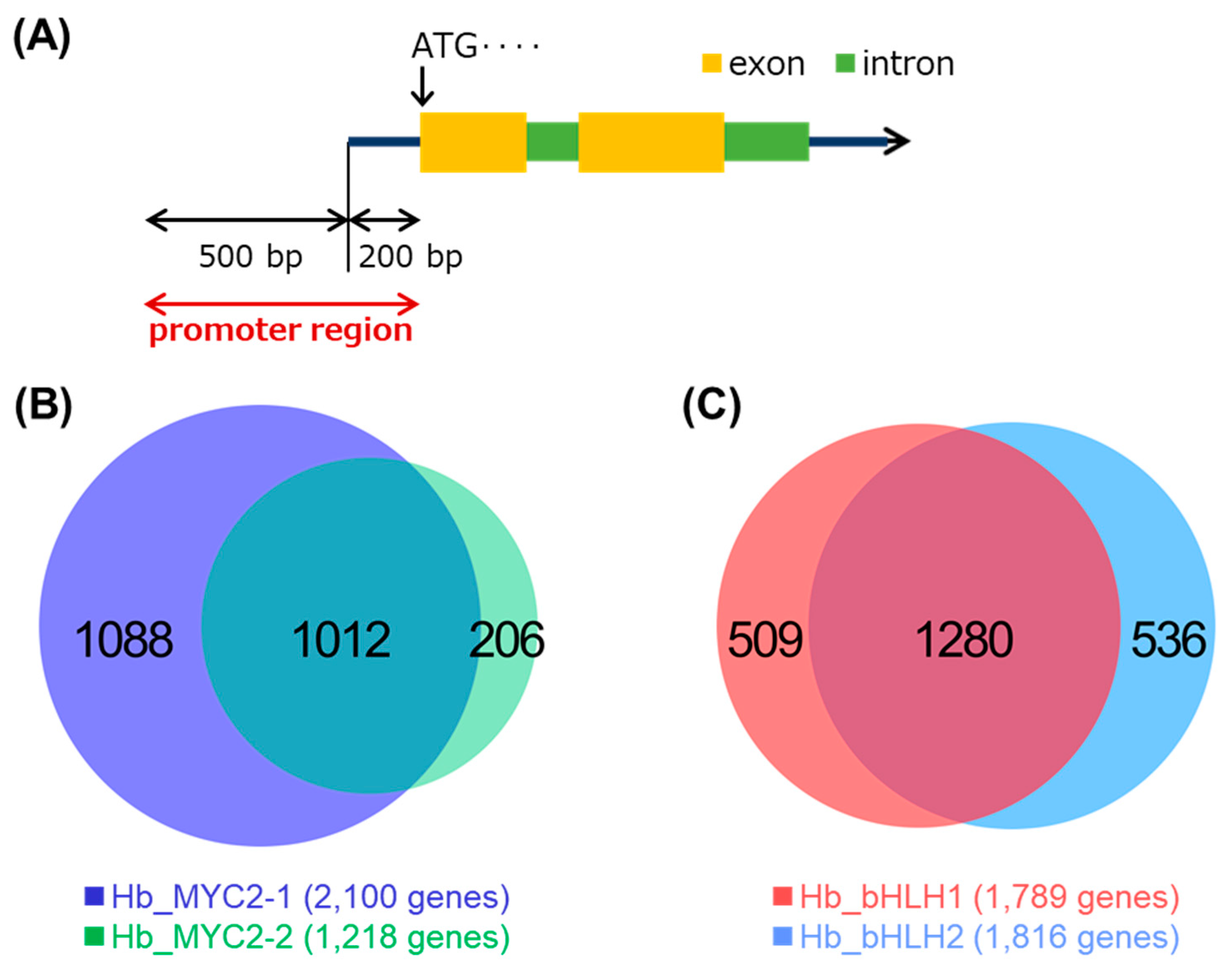
3.4. gDB-seq Analysis of Four bHLH TFs
3.5. DEGs Upon MeJA Include Candidate Genes Targeted by Hb_MYC2s
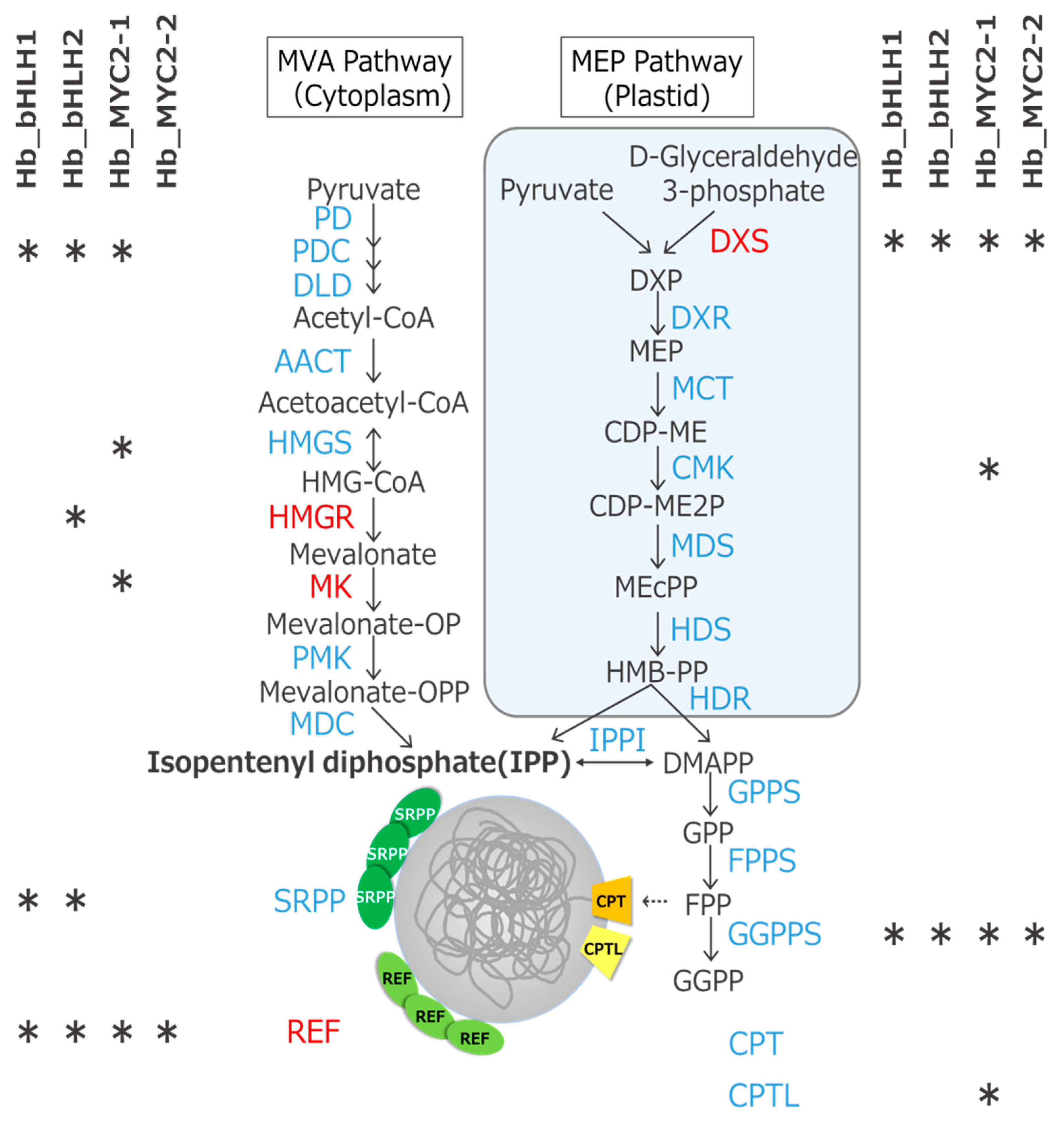
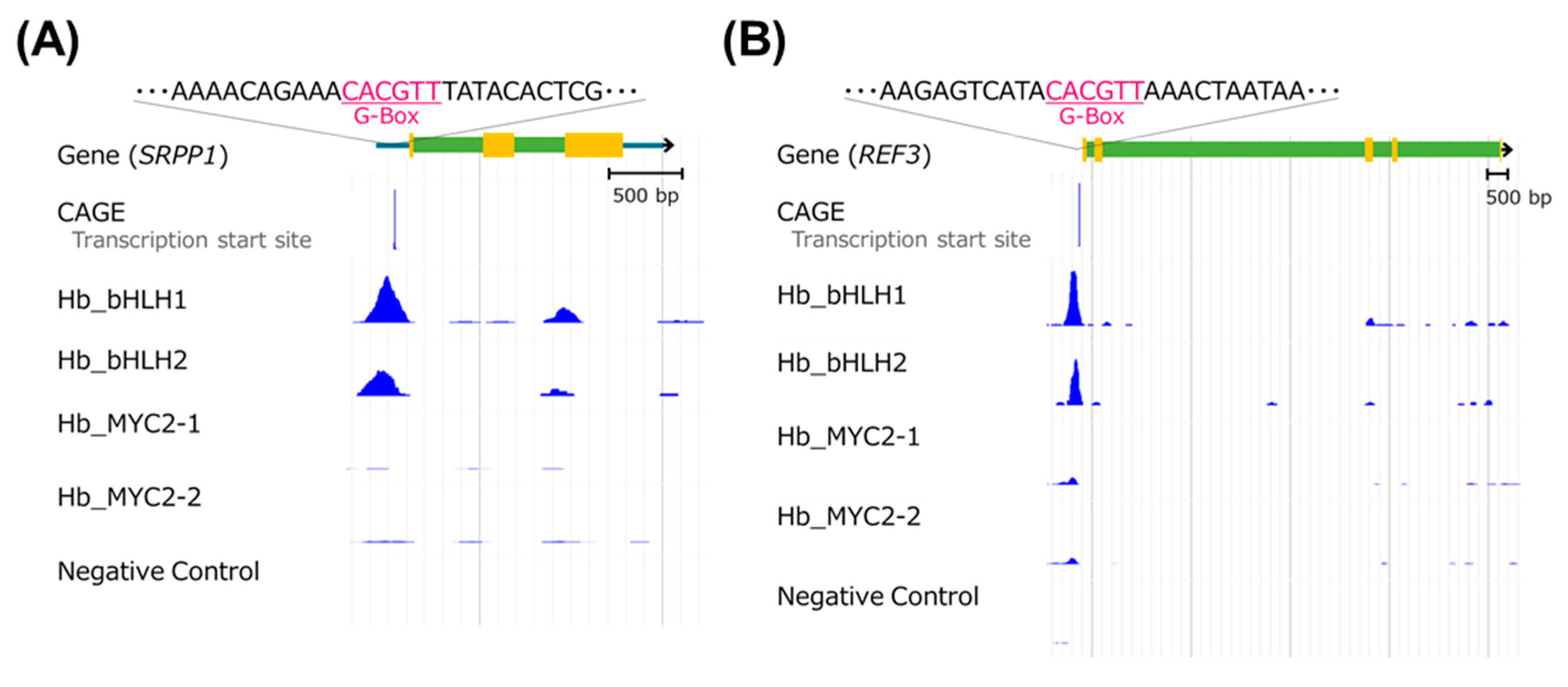
3.6. Candidate Genes of the Four TFs Include Rubber Biosynthesis-Related Ones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Van Beilen, J.B.; Poirier, Y. Establishment of new crops for the production of natural rubber. Trends Biotechnol. 2007, 25, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Mooibroek, H.; Cornish, K. Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 2000, 53, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Ryu, S.I.; Cornish, K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects. Plant Biotechnol. J. 2019, 17, 2041–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vranová, E.; Coman, D.; Gruissem, W. Network Analysis of the MVA and MEP Pathways for Isoprenoid Synthesis. Annu. Rev. Plant Boil. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Sando, T.; Takeno, S.; Watanabe, N.; Okumoto, H.; Kuzuyama, T.; Yamashita, A.; Hattori, M.; Ogasawara, N.; Fukusaki, E.; Kobayashi, A. Cloning and Characterization of the 2-C-Methyl-D-erythritol 4-Phosphate (MEP) Pathway Genes of a Natural-Rubber Producing Plant,Hevea brasiliensis. Biosci. Biotechnol. Biochem. 2008, 72, 2903–2917. [Google Scholar] [CrossRef] [Green Version]
- Asawatreratanakul, K.; Zhang, Y.-W.; Wititsuwannakul, D.; Wititsuwannakul, R.; Takahashi, S.; Rattanapittayaporn, A.; Koyama, T. Molecular cloning, expression and characterization of cDNA encoding cis-prenyltransferases from Hevea brasiliensis: A key factor participating in natural rubber biosynthesis. JBIC J. Boil. Inorg. Chem. 2003, 270, 4671–4680. [Google Scholar] [CrossRef] [Green Version]
- Dennis, M.S.; Light, D.R. Rubber elongation factor from Hevea brasiliensis. Identification, characterization, and role in rubber biosynthesis. J. Boil. Chem. 1989, 264, 18608–18617. [Google Scholar]
- Oh, S.K.; Kang, H.; Shin, D.-H.; Yang, J.; Chow, K.-S.; Yeang, H.Y.; Wagner, B.; Breiteneder, H.; Han, K.-H. Isolation, characterization, and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J. Boil. Chem. 1999, 274, 17132–17138. [Google Scholar] [CrossRef] [Green Version]
- Berthelot, K.; LeComte, S.; Estevez, Y.; Peruch, F. Hevea brasiliensis REF (Hev b 1) and SRPP (Hev b 3): An overview on rubber particle proteins. Biochimie 2014, 106, 1–9. [Google Scholar] [CrossRef]
- Yamashita, S.; Yamaguchi, H.; Waki, T.; Aoki, Y.; Mizuno, M.; Yanbe, F.; Ishii, T.; Funaki, A.; Tozawa, Y.; Miyagi-Inoue, Y.; et al. Identification and reconstitution of the rubber biosynthetic machinery on rubber particles from Hevea brasiliensis. eLife 2016, 5, 640. [Google Scholar] [CrossRef] [Green Version]
- Makita, Y.; Ng, K.K.; Singham, G.V.; Kawashima, M.; Hirakawa, H.; Sato, S.; Othman, A.S.; Matsui, M. Large-scale collection of full-length cDNA and transcriptome analysis in Hevea brasiliensis. Curr. Neuropharmacol. 2017, 24, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Hao, B.-Z.; Wu, J.-L. Laticifer Differentiation in Hevea brasiliensis: Induction by Exogenous Jasmonic Acid and Linolenic Acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Wang, J.; Li, Y.; Wu, S.; Yang, S.; Chao, J.; Chen, Y.; Zhang, S.; Shi, M.; Tian, W.-M. Comparative transcriptome analysis reveals phytohormone signalings, heat shock module and ROS scavenger mediate the cold-tolerance of rubber tree. Sci. Rep. 2018, 8, 4931. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, S.; Chao, J.; Deng, X.; Chen, Y.; Shi, M.; Tian, W.-M. Transcriptome Analysis of the Signalling Networks in Coronatine-Induced Secondary Laticifer Differentiation from Vascular Cambia in Rubber Trees. Sci. Rep. 2016, 6, 36384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirrello, J.; Leclercq, J.; Dessailly, F.; Rio, M.; Piyatrakul, P.; Kuswanhadi, K.; Tang, C.; Montoro, P. Transcriptional and post-transcriptional regulation of the jasmonate signalling pathway in response to abiotic and harvesting stress in Hevea brasiliensis. BMC Plant Boil. 2014, 14, 341. [Google Scholar] [CrossRef] [Green Version]
- Loh, S.C.; Othman, A.S.; Singham, G.V. Identification and characterization of jasmonic acid- and linolenic acid-mediated transcriptional regulation of secondary laticifer differentiation in Hevea brasiliensis. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Liu, J.-P.; Hu, J.; Liu, Y.-H.; Yang, C.-P.; Zhuang, Y.-F.; Guo, X.-L.; Li, Y.-J.; Zhang, L. Transcriptome analysis of Hevea brasiliensis in response to exogenous methyl jasmonate provides novel insights into regulation of jasmonate-elicited rubber biosynthesis. Physiol. Mol. Boil. Plants 2018, 24, 349–358. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The Master in Action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Zander, M.; Lewsey, M.G.; Clark, N.M.; Yin, L.; Bartlett, A.; Guzmán, J.P.S.; Hann, E.; Langford, A.E.; Jow, B.; Wise, A.; et al. Integrated multi-omics framework of the plant response to jasmonic acid. Nat. Plants 2020, 6, 290–302. [Google Scholar] [CrossRef]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.-M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, M.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH Transcription Factors MYC3 and MYC4 Are Targets of JAZ Repressors and Act Additively with MYC2 in the Activation of Jasmonate Responses[C][W]. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transciption. J. Exp. Bot. 2016, 68, 1303–1321. [Google Scholar] [CrossRef] [PubMed]
- Van Moerkercke, A.; Duncan, O.; Zander, M.; Šimura, J.; Broda, M.; Bossche, R.V.; Lewsey, M.G.; Lama, S.; Singh, K.B.; Ljung, K.; et al. A MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression and jasmonate levels. Proc. Natl. Acad. Sci. USA 2019, 116, 23345–23356. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Guo, N.; Yang, S.; Shi, M.; Chao, J.; Li, H.; Peng, S.; Tian, W.-M. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree, Hevea brasiliensis. J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Ohme-Takagi, M. Functional Analysis of Transcription Factors in Arabidopsis. Plant Cell Physiol. 2009, 50, 1232–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, Y.; Makita, Y.; Kawashima, M.; Hamasaki, H.; Yamamoto, Y.Y.; Matsui, M. Next-Generation Sequencing of Genomic DNA Fragments Bound to a Transcription Factor in Vitro Reveals Its Regulatory Potential. Genes 2014, 5, 1115–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, Y.; Makita, Y.; Shimohira, H.; Matsui, M. Time-Course Transcriptome Study Reveals Mode of bZIP Transcription Factors on Light Exposure in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1993. [Google Scholar] [CrossRef] [Green Version]
- Grafe, R. Isolation and characterization of cell lines of Nicotiana tabacum lacking nitrate reductase. Mol. Genet. Genom. 1978, 161, 67–76. [Google Scholar] [CrossRef]
- Lau, N.S.; Makita, Y.; Kawashima, M.; Taylor, T.D.; Kondo, S.; Othman, A.S.; Shu-Chien, A.C.; Matsui, M. The rubber tree genome shows expansion of gene family associated with rubber biosynthesis. Sci. Rep. 2016, 6, 28594. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.A.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2012, 31, 46–53. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makita, Y.; Kawashima, M.; Lau, N.S.; Othman, A.S.; Matsui, M. Construction of Pará rubber tree genome and multi-transcriptome database accelerates rubber researches. BMC Genom. 2018, 19, 922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Boil. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Buels, R.; Yao, E.; Diesh, C.M.; Hayes, R.; Munoz-Torres, M.; Helt, G.; Goodstein, D.; Elsik, C.G.; Lewis, S.; Stein, L.; et al. JBrowse: A dynamic web platform for genome visualization and analysis. Genome Boil. 2016, 17, 66. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, T.; A Meyer, C.; Eeckhoute, J.; Johnson, D.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based Analysis of ChIP-Seq (MACS). Genome Boil. 2008, 9, R137. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhou, L.-M.; Chen, Y.-Y.; Yang, S.-G.; Tian, W.-M. MYC genes with differential responses to tapping, mechanical wounding, ethrel and methyl jasmonate in laticifers of rubber tree (Hevea brasiliensis Muell. Arg.). J. Plant Physiol. 2011, 168, 1649–1658. [Google Scholar] [CrossRef]
- Zhai, J.; Hao, H.; Xiao, H.; Cao, Y.; Lin, X.; Huang, X. Identification of JAZ-interacting MYC transcription factors involved in latex drainage in Hevea brasiliensis. Sci. Rep. 2018, 8, 909. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Huang, S.-S.C.; Wise, A.; Castanon, R.; Nery, J.R.; Chen, H.; Watanabe, M.; Thomas, J.; Bar-Joseph, Z.; Ecker, J.R. A transcription factor hierarchy defines an environmental stress response network. Science 2016, 354, aag1550. [Google Scholar] [CrossRef] [Green Version]
- Yazaki, J.; Galli, M.; Kim, A.Y.; Nito, K.; Aleman, F.; Chang, K.N.; Carvunis, A.-R.; Quan, R.; Nguyen, H.; Song, L.; et al. Mapping transcription factor interactome networks using HaloTag protein arrays. Proc. Natl. Acad. Sci. USA 2016, 113, E4238–E4247. [Google Scholar] [CrossRef] [Green Version]
- Omalley, R.; Huang, S.-S.C.; Song, L.; Lewsey, M.G.; Bartlett, A.; Nery, J.R.; Galli, M.; Gallavotti, A.; Ecker, J.R. Cistrome and Epicistrome Features Shape the Regulatory DNA Landscape. Cell 2016, 165, 1280–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Li, H.; Guo, D.; Peng, S. Identification and characterization of the EIN3/EIL gene family in Hevea brasiliensis. Tree Genet. Genomes 2015, 11, 51. [Google Scholar] [CrossRef]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The Basic Helix-Loop-Helix Transcription Factor Family in Plants: A Genome-Wide Study of Protein Structure and Functional Diversity. Mol. Boil. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Time | 4 h | 24 h | 5 Days | |
|---|---|---|---|---|
| UP-regulated genes | 203 | 275 | 185 | |
| TF | UP-regulated | 15 | 23 | 6 |
| UP-regulated | NAC | ERF, WRKY, MYB, Tify, MYB-related | ― | |
| & | ||||
| Hb_MYC2 binding | ||||
| Rubber biosynthesis-related | UP-regulated | 1 | 1 | 2 |
| UP-regulated | ― | ― | MK3, REF7 | |
| & | ||||
| Hb_MYC2 binding | ||||
| DOWN-regulated genes | 167 | 144 | 181 | |
| TF | DOWN-regulated | 19 | 9 | 12 |
| DOWN-regulated | C2C2-GATA | M-type | ― | |
| & | ||||
| Hb_MYC2 binding | ||||
| Rubber biosynthesis-related | DOWN-regulated | 0 | 0 | 0 |
| DOWN-regulated | ― | ― | ― | |
| & | ||||
| Hb_MYC2 binding | ||||
| Sample | Number of Peaks | Number of Peaks in Promoter Regions | Number of TF-Targeted Candidate Genes |
|---|---|---|---|
| Hb_MYC2-1 | 39,429 | 2105 | 2100 |
| Hb_MYC2-2 | 24,291 | 1219 | 1218 |
| Hb_bHLH1 | 20,809 | 1797 | 1789 |
| Hb_bHLH2 | 18,995 | 1823 | 1816 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, T.; Kurihara, Y.; Makita, Y.; Okubo-Kurihara, E.; Kageyama, A.; Osada, E.; Shimada, S.; Tsuchida, H.; Shimada, H.; Matsui, M. Regulatory Potential of bHLH-Type Transcription Factors on the Road to Rubber Biosynthesis in Hevea brasiliensis. Plants 2020, 9, 674. https://doi.org/10.3390/plants9060674
Yamaguchi T, Kurihara Y, Makita Y, Okubo-Kurihara E, Kageyama A, Osada E, Shimada S, Tsuchida H, Shimada H, Matsui M. Regulatory Potential of bHLH-Type Transcription Factors on the Road to Rubber Biosynthesis in Hevea brasiliensis. Plants. 2020; 9(6):674. https://doi.org/10.3390/plants9060674
Chicago/Turabian StyleYamaguchi, Tomoko, Yukio Kurihara, Yuko Makita, Emiko Okubo-Kurihara, Ami Kageyama, Emi Osada, Setsuko Shimada, Hiroko Tsuchida, Hiroaki Shimada, and Minami Matsui. 2020. "Regulatory Potential of bHLH-Type Transcription Factors on the Road to Rubber Biosynthesis in Hevea brasiliensis" Plants 9, no. 6: 674. https://doi.org/10.3390/plants9060674