Comparison of Antimicrobial Resistance and Pan-Genome of Clinical and Non-Clinical Enterococcus cecorum from Poultry Using Whole-Genome Sequencing


 and
and
Abstract
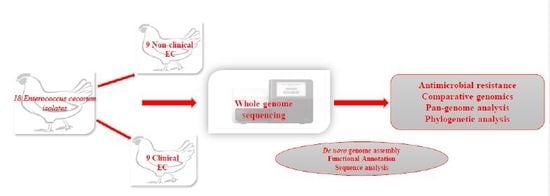
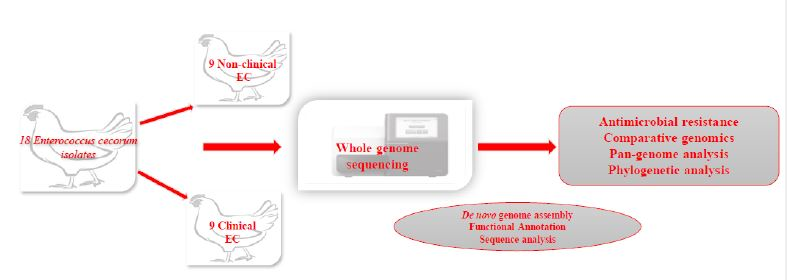
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and DNA Extraction
2.2. Sequencing, Assembly, and Annotation
2.3. Antimicrobial Susceptibility Testing and Prediction of Antimicrobial Resistance Genes
2.4. Pan-Genome Construction
2.5. Core-Genome Phylogenetic Tree
2.6. Nucleotide Accession Numbers
3. Results
3.1. Genome Characteristics
3.2. Antimicrobial Resistance Genes
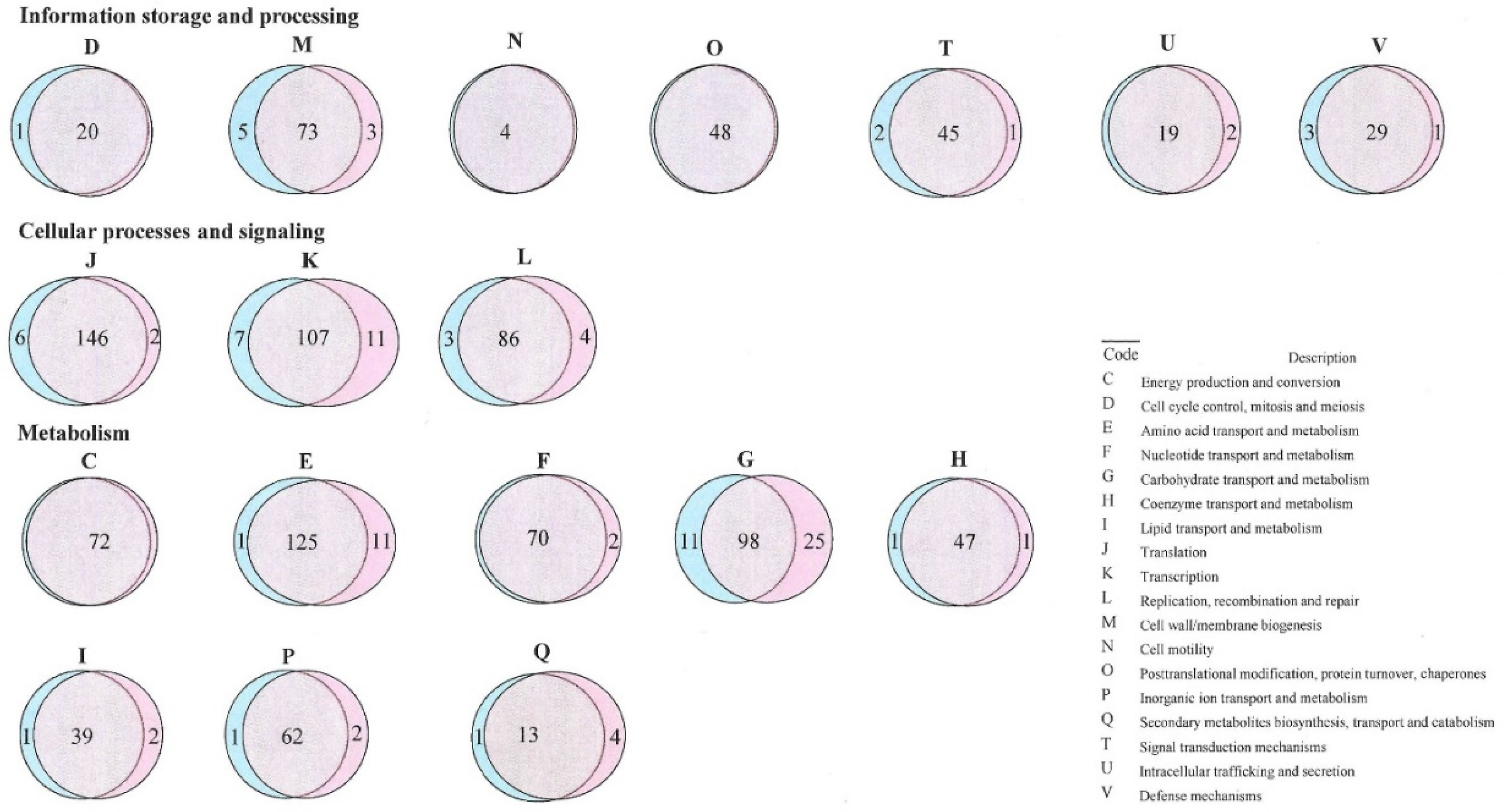
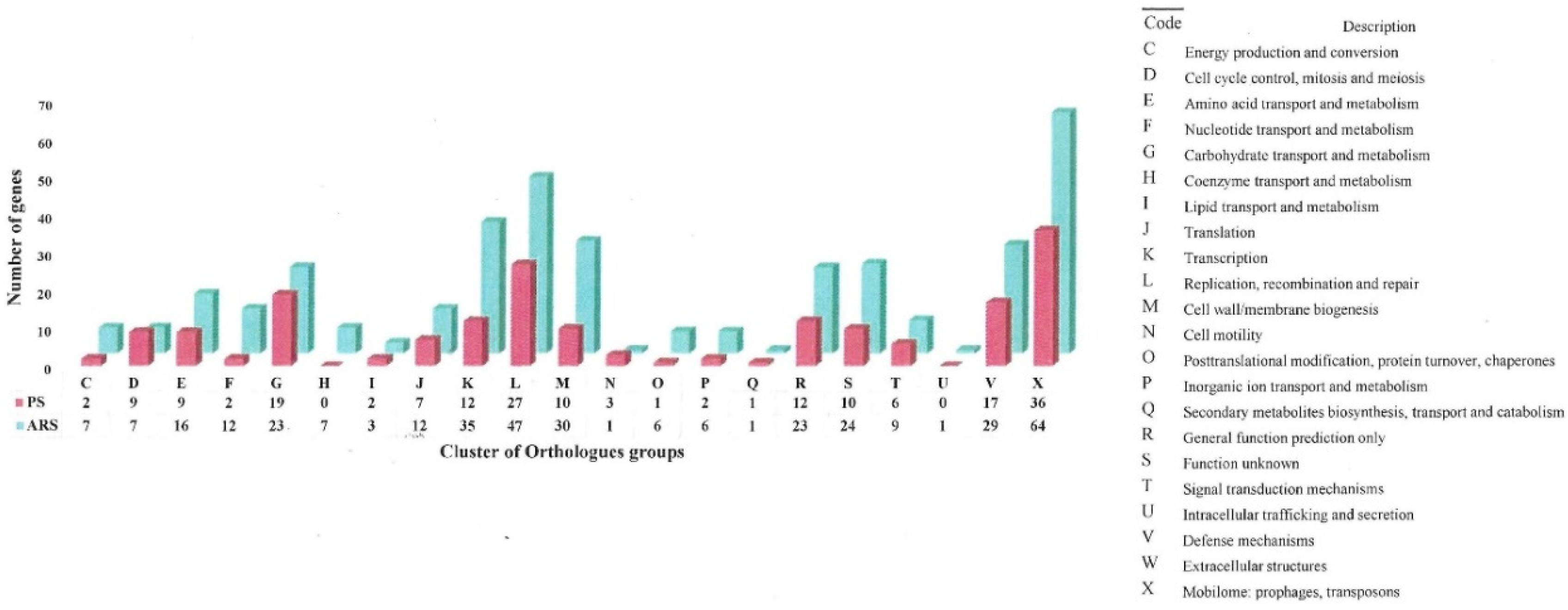
3.3. Pan-Genome Analysis
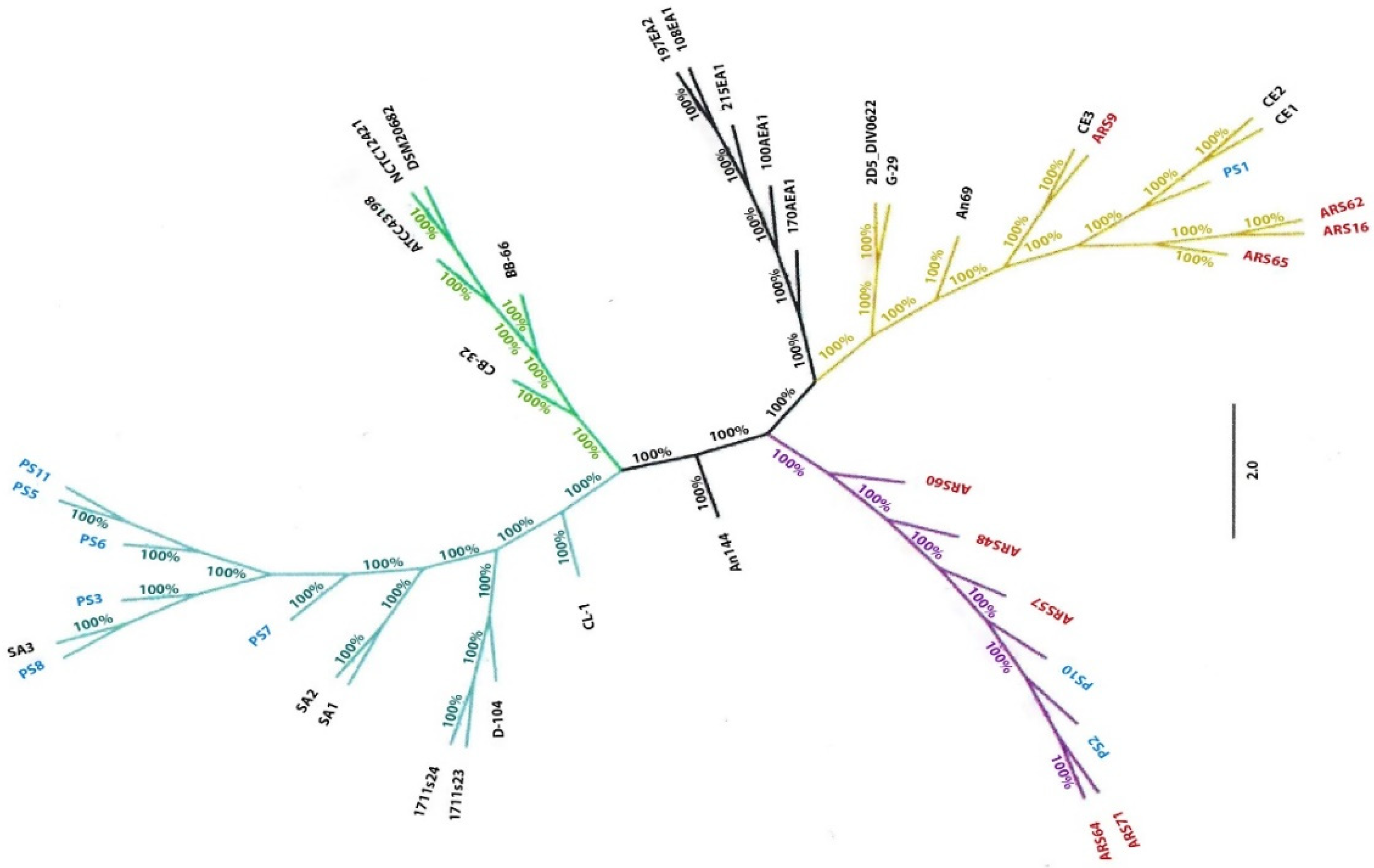
3.4. Core Genome Phylogenetic Analysis
3.5. Prophage Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Devriese, L.A.; Hommez, J.; Wijfels, R.; Haesebrouck, F. Composition of the enterococcal and streptococcal intestinal flora of poultry. J. Appl. Bacteriol. 1991, 71, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.A.; Laurier, L.; De Herdt, P.; Haesebrouck, F. Enterococcal and streptococcal species isolated from faeces of calves, young cattle and dairy cows. J. Appl. Bacteriol. 1992, 72, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.A.; Hommez, J.; Pot, B.; Haesebrouck, F. Identification and composition of the streptococcal and enterococcal flora of tonsils, intestines and faeces of pigs. J. Appl. Bacteriol. 1994, 77, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Baele, M.; Devriese, L.A.; Butaye, P.; Haesebrouck, F. Composition of enterococcal and streptococcal flora from pigeon intestines. J. Appl. Microbiol. 2002, 92, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Forster, R.J.; Yu, H.; Chambers, J.R.; Wheatcroft, R.; Sabour, P.M.; Chen, S. Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum. FEMS Microbiol. Ecol. 2002, 41, 171–179. [Google Scholar] [CrossRef]
- Devriese, L.A.; Dutta, G.N.; Farrow, J.A.E.; van de Kerckhove, A.; Phillips, B.A. Streptococcus cecorum, a new species isolated from chickens. Int. J. Syst. Bacteriol. 1983, 33, 772–776. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.M.; MacKenzie, G.; McGiliveray, N.C.; Brown, L.; Devriese, L.A.; Baele, M. Isolation of Enterococcus cecorum from bone lesions in broiler chickens. Vet. Rec. 2002, 150, 27. [Google Scholar]
- Devriese, L.A.; Cauwerts, K.; Hermans, K.; Wood, A. Enterococcus cecorum septicemia as a cause of bone and joint lesions resulting in lameness in broiler chickens. Vlaam. Diergeneeskd. Tijdschr. 2002, 71, 219–221. [Google Scholar]
- Borst, L.B.; Suyemoto, M.M.; Sarsour, A.H.; Harris, M.C.; Martin, M.P.; Strickland, J.D.; Oviedo, E.O.; Barnes, H.J. Pathogenesis of Enterococcal Spondylitis Caused by Enterococcus cecorum in Broiler Chickens. Vet. Pathol. 2017, 54, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.; Petersen, H.; Teske, L.; Rautenschlein, S. Colonization patterns of Enterococcus cecorum in two different broiler production cycles detected with a newly developed quantitative real-time PCR. BMC Microbiol. 2017, 17, 106. [Google Scholar] [CrossRef]
- De Herdt, P.D.P.; Van Steelant, J.; Swam, H.; Tanghe, L.; Van Goethem, S.; Vanrobaeys, M. Enterococcus cecorum osteomyelitis and arthritis in broiler chickens. Vlaams Diergeneeskd. Tijdschr. 2008, 78, 44–48. [Google Scholar]
- Jung, A.; Rautenschlein, S. Comprehensive report of an Enterococcus cecorum infection in a broiler flock in Northern Germany. BMC Vet. Res. 2014, 10, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makrai, L.; Nemes, C.; Simon, A.; Ivanics, E.; Dudas, Z.; Fodor, L.; Glavits, R. Association of Enterococcus cecorum with vertebral osteomyelitis and spondylolisthesis in broiler parent chicks. Acta Vet. Hung. 2011, 59, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Kense, M.J.; Landman, W.J. Enterococcus cecorum infections in broiler breeders and their offspring: Molecular epidemiology. Avian Pathol. 2011, 40, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Szeleszczuk, P.D.B.; Zbikowski, A.; Dolka, I.; Peryga, M. First case of enterococcal spondylitis in broiler chickens in Poland. Med. Weter 2013, 69, 298–303. [Google Scholar]
- Albini, F.; Lobsiger, S.T.; Renggli, H. Infection with the capnophilic bacteria Enterococcus cecorum in broiler chickens. Schweiz Arch. Tierheilkd 2014, 156, 295–298. [Google Scholar] [CrossRef]
- Stalker, M.J.; Brash, M.L.; Weisz, A.; Ouckama, R.M.; Slavic, D. Arthritis and osteomyelitis associated with Enterococcus cecorum infection in broiler and broiler breeder chickens in Ontario, Canada. J. Vet. Diagn. Investig. 2010, 22, 643–645. [Google Scholar] [CrossRef] [Green Version]
- Robbins, K.M.; Suyemoto, M.M.; Lyman, R.L.; Martin, M.P.; Barnes, H.J.; Borst, L.B. An outbreak and source investigation of enterococcal spondylitis in broilers caused by Enterococcus cecorum. Avian Dis. 2012, 56, 768–773. [Google Scholar] [CrossRef]
- Borst, L.B.; Suyemoto, M.M.; Robbins, K.M.; Lyman, R.L.; Martin, M.P.; Barnes, H.J. Molecular epidemiology of Enterococcus cecorum isolates recovered from enterococcal spondylitis outbreaks in the southeastern United States. Avian Pathol. 2012, 41, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Wijetunge, D.S.; Dunn, P.; Wallner-Pendleton, E.; Lintner, V.; Lu, H.; Kariyawasam, S. Fingerprinting of poultry isolates of Enterococcus cecorum using three molecular typing methods. J. Vet. Diagn. Investig. 2012, 24, 1166–1171. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.R.; Kariyawasam, S.; Borst, L.B.; Frye, J.G.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A. Antimicrobial resistance, virulence determinants and genetic profiles of clinical and nonclinical Enterococcus cecorum from poultry. Lett. Appl. Microbiol. 2015, 60, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Aitchison, H.; Poolman, P.; Coetzer, M.; Griffiths, C.; Jacobs, J.; Meyer, M.; Bisschop, S. Enterococcal-related vertebral osteoarthritis in South African broiler breeders: A case report. J. S. Afr. Vet. Assoc. 2014, 85, 1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeshan, B.; Khaing, A.T.; Daud, N.H. Enterococcal-Associated Vertebral Osteoarthritis (Evoa) in Broiler Chicken in Malaysia: A Case Report. In Proceedings of the 2nd World Veterinary Poultry Association (WVPA) and World Poultry Science Association (WPSA) (Malaysia Branch) Scientifix Conference, Kuala Lumpur, Malaysia, 21–23 September 2015; pp. 77–80. [Google Scholar]
- Jung, A.; Chen, L.R.; Suyemoto, M.M.; Barnes, H.J.; Borst, L.B. A Review of Enterococcus cecorum Infection in Poultry. Avian Dis. 2018, 62, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Borst, L.B.; Suyemoto, M.M.; Scholl, E.H.; Fuller, F.J.; Barnes, H.J. Comparative genomic analysis identifies divergent genomic features of pathogenic Enterococcus cecorum including a type IC CRISPR-Cas system, a capsule locus, an epa-like locus, and putative host tissue binding proteins. PLoS ONE 2015, 10, e0121294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Metzner, M.; Ryll, M. Comparison of pathogenic and non-pathogenic Enterococcus cecorum strains from different animal species. BMC Microbiol. 2017, 17, 33. [Google Scholar] [CrossRef] [Green Version]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2015, 43, D261–D269. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Panchenko, A.R.; Shoemaker, B.A.; Thiessen, P.A.; Geer, L.Y.; Bryant, S.H. CDD: A database of conserved domain alignments with links to domain three-dimensional structure. Nucleic Acids Res. 2002, 30, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Prabha, R.; Singh, D.P.; Somvanshi, P.; Rai, A. Functional profiling of cyanobacterial genomes and its role in ecological adaptations. Genom. Data 2016, 9, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- McDermott, P.F.; Tyson, G.H.; Kabera, C.; Chen, Y.; Li, C.; Folster, J.P.; Ayers, S.L.; Lam, C.; Tate, H.P.; Zhao, S. Whole-Genome Sequencing for detecting antimicrobial resistance in nontyphoidal Salmonella. Antimicrob. Agents Chemother. 2016, 60, 5515–5520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Tyson, G.H.; Chen, Y.; Li, C.; Mukherjee, S.; Young, S.; Lam, C.; Folster, J.P.; Whichard, J.M.; McDermott, P.F. Whole-Genome Sequencing analysis accurately predicts antimicrobial resistance phenotypes in Campylobacter spp. Appl. Environ. Microbiol. 2016, 82, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.B.; Frimodt-Moller, N.; Aarestrup, F.M. Presence of erm gene classes in gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef]
- Roberts, M.C.; Sutcliffe, J.; Courvalin, P.; Jensen, L.B.; Rood, J.; Seppala, H. Nomenclature for macrolide and macrolide-lincosamide-streptogramin B resistance determinants. Antimicrob. Agents Chemother. 1999, 43, 2823–2830. [Google Scholar] [CrossRef] [Green Version]
- Rollins, L.D.; Lee, L.N.; LeBlanc, D.J. Evidence for a disseminated erythromycin resistance determinant mediated by Tn917-like sequences among group D streptococci isolated from pigs, chickens, and humans. Antimicrob. Agents Chemother. 1985, 27, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Boerlin, P.; Nicholson, V.; Brash, M.; Slavic, D.; Boyen, F.; Sanei, B.; Butaye, P. Diversity of Enterococcus cecorum from chickens. Vet. Microbiol. 2012, 157, 405–411. [Google Scholar] [CrossRef]
- Dolka, B.; Chrobak-Chmiel, D.; Makrai, L.; Szeleszczuk, P. Phenotypic and genotypic characterization of Enterococcus cecorum strains associated with infections in poultry. BMC Vet. Res. 2016, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A. Prevalence of streptogramin resistance in enterococci from animals: Identification of vatD from animal sources in the USA. Int. J. Antimicrob. Agents 2007, 30, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A. First report of vatB and vgaB from Enterococcus gallinarum in the USA. Int. J. Antimicrob. Agents 2008, 31, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; Leon-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Li, T.; Zhu, Y.; Wang, B.; Zou, X.; Li, M. Mechanisms of linezolid resistance in staphylococci and enterococci isolated from two teaching hospitals in Shanghai, China. BMC Microbiol. 2014, 14, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.E.; Deshpande, L.M.; Jones, R.N. Linezolid update: Stable in vitro activity following more than a decade of clinical use and summary of associated resistance mechanisms. Drug Resist. Updat. 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Hou, Q.; Wang, Y.; Li, J.; Li, W.; Kwok, L.Y.; Sun, Z.; Zhang, H.; Zhong, Z. Comparative genomic analysis of Enterococcus faecalis: Insights into their environmental adaptations. BMC Genom. 2018, 19, 527. [Google Scholar] [CrossRef] [Green Version]
- Raven, K.E.; Reuter, S.; Gouliouris, T.; Reynolds, R.; Russell, J.E.; Brown, N.M.; Torok, M.E.; Parkhill, J.; Peacock, S.J. Genome-based characterization of hospital-adapted Enterococcus faecalis lineages. Nat. Microbiol. 2016, 1, 15033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakshi, U.; Sarkar, M.; Paul, S.; Dutta, C. Assessment of virulence potential of uncharacterized Enterococcus faecalis strains using pan genomic approach—Identification of pathogen-specific and habitat-specific genes. Sci. Rep. 2016, 6, 38648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghattargi, V.C.; Gaikwad, M.A.; Meti, B.S.; Nimonkar, Y.S.; Dixit, K.; Prakash, O.; Shouche, Y.S.; Pawar, S.P.; Dhotre, D.P. Comparative genome analysis reveals key genetic factors associated with probiotic property in Enterococcus faecium strains. BMC Genom. 2018, 19, 652. [Google Scholar] [CrossRef] [PubMed]
- Medini, D.; Donati, C.; Tettelin, H.; Masignani, V.; Rappuoli, R. The microbial pan-genome. Curr. Opin. Genet. Dev. 2005, 15, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Ghattargi, V.C.; Nimonkar, Y.S.; Burse, S.A.; Davray, D.; Kumbhare, S.V.; Shetty, S.A.; Gaikwad, M.A.; Suryavanshi, M.V.; Doijad, S.P.; Utage, B.; et al. Genomic and physiological analyses of an indigenous strain, Enterococcus faecium 17OM39. Funct. Integr. Genom. 2018, 18, 385–399. [Google Scholar] [CrossRef]
- Naito, M.; Pawlowska, T.E. The role of mobile genetic elements in evolutionary longevity of heritable endobacteria. Mob. Genet. Elem. 2016, 6, e1136375. [Google Scholar] [CrossRef] [Green Version]
- Jacobitz, A.W.; Kattke, M.D.; Wereszczynski, J.; Clubb, R.T. Sortase Transpeptidases: Structural Biology and Catalytic Mechanism. Adv. Protein Chem. Struct. Biol. 2017, 109, 223–264. [Google Scholar] [CrossRef] [Green Version]
- Bonacina, J.; Suarez, N.; Hormigo, R.; Fadda, S.; Lechner, M.; Saavedra, L. A genomic view of food-related and probiotic Enterococcus strains. DNA Res. 2017, 24, 11–24. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Resistance Profile | Antibiotics Resistance Genes |
|---|---|---|
| PS1 | Lincomycin-Tetracycline | lnuB, linB, tetL, tetM, tetO |
| PS2 | Erythromycin-Kanamycin-Lincomycin-Tylosin | ermG, ermB, mefA, msrD, aph3-III, lnuC |
| PS3 | Erythromycin-Streptomycin-Tetracycline | ermG, mefA, msrD, tetL, tetM |
| PS5 | Erythromycin-Tylosin-Tetracycline | ermG, mefA, msrD, tetL, tetM |
| PS6 | Erythromycin-Tetracycline | ermG, mefA, msrD, tetL, tetM |
| PS7 | Lincomycin | lnuC |
| PS8 | Erythromycin-Streptomycin-Tetracycline | ermG, mefA, msrD, tetL, tetM |
| PS11 | Erythromycin-Tetracycline | ermG, mefA, msrD, tetL, tetM |
| ARS9 | Linezolid-Quinupristin/Dalfopristin | ermB |
| ARS16 | Erythromycin-Kanamycin-Lincomycin-Quinupristin/Dalfopristin-Tetracycline | mefA, msrD, aph3-III, lnuB, linB, tetL, tetM, tetO |
| ARS48 | Erythromycin-Lincomycin-Quinupristin/Dalfopristin-Tylosin-Tetracycline | ermB, mefA, msrD, lnuB, linB, tetM |
| ARS57 | Lincomycin-Linezolid-Quinupristin/Dalfopristin-Tylosin-Tetracycline | ermB, mefA, msrD, lnuB, linB, lnuD |
| ARS60 | Kanamycin-Lincomycin-Quinupristin/Dalfopristin-Tetracycline | ermB, msrD, aph3-III, lnuB, linB, tetL, tetM |
| ARS62 | Kanamycin-Lincomycin-Quinupristin/Dalfopristin-Tylosin-Tetracycline | ermB, msrD, aph3-III, lnuB, linB, tetL, tetM, tetO |
| ARS64 | Erythromycin-Lincomycin-Tylosin-Tetracycline | ermB, tetL, tetM |
| ARS65 | Chloramphenicol-Lincomycin-Linezolid-Quinupristin/Dalfopristin-Tylosin-Tetracycline | ermB, mefA, msrD, lnuB, linB, tetL, tetM, tetO |
| ARS71 | Erythromycin-Lincomycin-Tylosin-Tetracycline | ermB, tetL, tetM |
| Isolate | Length | GC | CDS | Best Match | |
|---|---|---|---|---|---|
| Phage | Accession Number | ||||
| ARS16 | 36.3 | 37.17 | 47 | PHAGE_Strept_5093 | NC_012753 |
| ARS48 | 31.1 | 37.87 | 43 | PHAGE_Entero_EFC_1 | NC_025453 |
| ARS57 | 47.3 | 37.59 | 52 | PHAGE_Entero_EFC_1 | NC_025453 |
| ARS62 | 36.3 | 37.17 | 47 | PHAGE_Strept_5093 | NC_012753 |
| ARS64 | 36.1 | 35.41 | 43 | PHAGE_Lister_vB_LmoS_188 | NC_028871 |
| ARS65 | 33.6 | 37.60 | 43 | PHAGE_Entero_EFC_1 | NC_025453 |
| 34.4 | 36.85 | 23 | PHAGE_Bacter_Diva | NC_028788 | |
| 14.7 | 39.26 | 17 | PHAGE_Strept_5093 | NC_012753 | |
| 14.5 | 38.74 | 16 | PHAGE_Strept_5093 | NC_012753 | |
| PS2 | 23.9 | 37.30 | 35 | PHAGE_Strept_5093 | NC_012753 |
| PS5 | 30.7 | 35.37 | 36 | PHAGE_Bacill_phBC6A52 | NC_004821 |
| 35.8 | 36.8 | 47 | PHAGE_Strept_5093 | NC_012753 | |
| PS6 | 30.7 | 35.37 | 36 | PHAGE_Bacill_phBC6A52 | NC_004821 |
| 35.8 | 36.8 | 47 | PHAGE_Strept_5093 | NC_012753 | |
| PS8 | 32.2 | 36.34 | 39 | PHAGE_Strept_5093 | NC_012753 |
| PS11 | 30.7 | 35.37 | 36 | PHAGE_Bacill_phBC6A52 | NC_004821 |
| 35.8 | 36.8 | 47 | PHAGE_Strept_5093 | NC_012753 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, P.; Gupta, S.K.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A.; Kariyawasam, S.; Frye, J.G.; Jackson, C.R. Comparison of Antimicrobial Resistance and Pan-Genome of Clinical and Non-Clinical Enterococcus cecorum from Poultry Using Whole-Genome Sequencing. Foods 2020, 9, 686. https://doi.org/10.3390/foods9060686
Sharma P, Gupta SK, Barrett JB, Hiott LM, Woodley TA, Kariyawasam S, Frye JG, Jackson CR. Comparison of Antimicrobial Resistance and Pan-Genome of Clinical and Non-Clinical Enterococcus cecorum from Poultry Using Whole-Genome Sequencing. Foods. 2020; 9(6):686. https://doi.org/10.3390/foods9060686
Chicago/Turabian StyleSharma, Poonam, Sushim K. Gupta, John B. Barrett, Lari M. Hiott, Tiffanie A. Woodley, Subhashinie Kariyawasam, Jonathan G. Frye, and Charlene R. Jackson. 2020. "Comparison of Antimicrobial Resistance and Pan-Genome of Clinical and Non-Clinical Enterococcus cecorum from Poultry Using Whole-Genome Sequencing" Foods 9, no. 6: 686. https://doi.org/10.3390/foods9060686