Glycera sheikhmujibi n. sp. (Annelida: Polychaeta: Glyceridae): A New Species of Glyceridae from the Saltmarsh of Bangladesh

1
Department of Fisheries and Marine Science, Noakhali Science and Technology University, Sonapur 3814, Noakhali, Bangladesh
2
Research Cell, Noakhali Science and Technology University, Sonapur 3814, Noakhali, Bangladesh
3
Australian Museum Research Institute, Australian Museum, 1 William Street, NSW 2010, Australia
4
Department of Biological Sciences, Macquarie University, North Ryde 2109, Australia
*
Author to whom correspondence should be addressed.
http://zoobank.org/urn:lsid:zoobank.org:pub:C2CEE260-5B4E-43B7-9251-2A0AB37EAB59
Diversity 2020, 12(6), 213; https://doi.org/10.3390/d12060213
Submission received: 8 May 2020
/
Revised: 22 May 2020
/
Accepted: 24 May 2020
/
Published: 26 May 2020
(This article belongs to the Section Animal Diversity)
Abstract
:A new species of glycerid polychaete, Glycera sheikhmujibi, is described from the saltmarsh on the central coast of Bangladesh. The species is identified based on morphological characteristics using both a light microscope and scanning electron microscope (SEM). The species is characterized by the presence of three distinct types of proboscideal papillae: type 1 papillae (conical with three transverse ridges), type 2 (conical with a straight, median, longitudinal ridge), and type 3 (round, shorter, and broader, with a straight, median, longitudinal ridge). It has a Y-shaped aileron with gently incised triangular base, almost equal-size digitiform noto- and neuropodial lobes in the mid-body, and long ventral cirri at the posterior end. The new species is compared with its related species, previously described from the Bay of Bengal region. A key to all these species is provided.
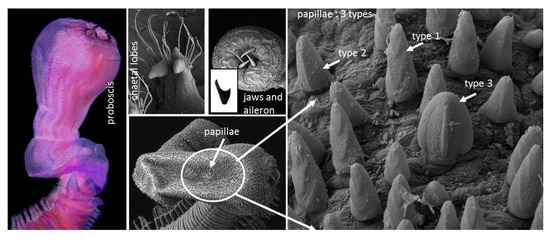
1. Introduction
The family Glyceridae currently has 87 accepted species (80 species of Glycera, one species of Glycerella, and five species of Hemipodus) (World Register of Marine Species, WoRMS; www.marinespecies.org). Among the three genera, Glycera and Glycerella possess biramous parapodia, whereas Hemipodus includes species with only uniramous parapodia throughout the body [1]. The ailerons, accessory supports on the proboscis, are rod-like in Glycerella, and mostly triangular-shaped or a more complicated structure having outer and inner rami in Glycera. Both Glycera and Hemipodus have spinigerous compound chaetae, but in Glycerella, additional compound falcigers are present [1]. Böggemann [2] revised all species of Glycera previously described worldwide and synonymised many of the previously described species (166), accepting only 36 as valid species. This was based on morphological data only, but subsequently he has used molecular studies to support some of these widely distributed species [3,4]. They are easily distinguishable from other polychaetes, as they have a pointed prostomium and eversible axial proboscis with numerous papillae. Glycera are widely distributed from tropical to temperate regions, and from intertidal to abyssal depths, inhabiting mainly soft bottom (sand/mud) sediments [1,3,5,6]. Glycerids are generally considered to be carnivorous burrowers, capturing and killing prey with their strong, well-developed jaws connected to venom glands that supply venom [7,8].
Polychaetes have been poorly studied in Bangladesh. While some benthic ecological studies have been carried out in the area [9,10,11], these studies have provided no taxonomic details, apart from the listing of polychaete species and their abundance data. None of these studies have deposited any material, so the validity of these species cannot be confirmed. Pramanik et al. [12] reported a new record of Glyceridae, Glycera lancadivae Schmarda, 1861 [13], which seems to be similar to Glycera brevicirris Grube, 1870 [14] (known from Sri Lanka and the Andaman Sea).
Of the 80 accepted species of Glycera Lamarck, 1818 [15], only 10 species have been recorded from the Bay of Bengal region (WoRMS; www.marinespecies.org). This region includes Bangladesh, India, Myanmar, Sri Lanka, and the tip of Andaman Nicobar Islands. Only one of these 10 species has been recorded from Bangladesh [12], in the northeastern corner of the Bay of Bengal: G. lancadivae Schmarda, 1861 [13]. Muir and Hossain [16] reported an unidentifiable glycerid fragment from the Halishahar Coast of Bangladesh, and they also provided taxonomic keys for identifying 14 species from the Bay of Bengal and Indo-Pacific regions. Subsequently, Hossain and Hutchings [17] emphasized the possibility of undescribed polychaete taxa from Bangladeshi coastal waters. Hence, the aim of this report is to describe a new species of Glycera from the Bangladesh Coast, and to provide an updated key to all species of Glycera recorded from the Bay of Bengal region.
2. Materials and Methods
2.1. Study Site Description
The study area is located in an upper tidal channel of the lower Meghna river estuary, the largest estuarine ecosystem of Bangladesh, characterized by sunny tropical weather with monsoonal influence [18,19]. Mean annual temperature and rainfall in the study area are 25.5 °C and 2980 mm, respectively. According to the Köppen–Geiger climate classification, this climate is considered to be Am (tropical monsoon climate). The monsoon is characterized by strong southeastern winds with high rainfall, humidity, cloud cover, thunderstorms, cyclones, and occasional storm surges [18,19]. Almost all year round, the area is influenced by the incoming tide from Bay of Bengal. Tides are of a semi-diurnal type, with two high and two low waters during a lunar day. The tide varies with respect to magnitude, ranging from 0.07 m during neap tide to 4.42 m during spring tide [20]. The wave height of the estuary varies from 0 to 4 m [21]. However, the tidal wave is considerably slanted as it moves inside the channel, so that with increasing distance from the channel opening, the duration of flood becomes shorter than during the ebb tide. The lower, deeper areas near the opening of the channel are characterized by strong estuarine influence, with higher current velocity and stronger tides, while the estuarine water inflow is substantially reduced in the upper shallow areas. In addition, the upper part of the channel receives a continuous freshwater supply, especially during monsoons, through a small system of tidal creeks and streams.
2.2. Sample Collection and Analysis
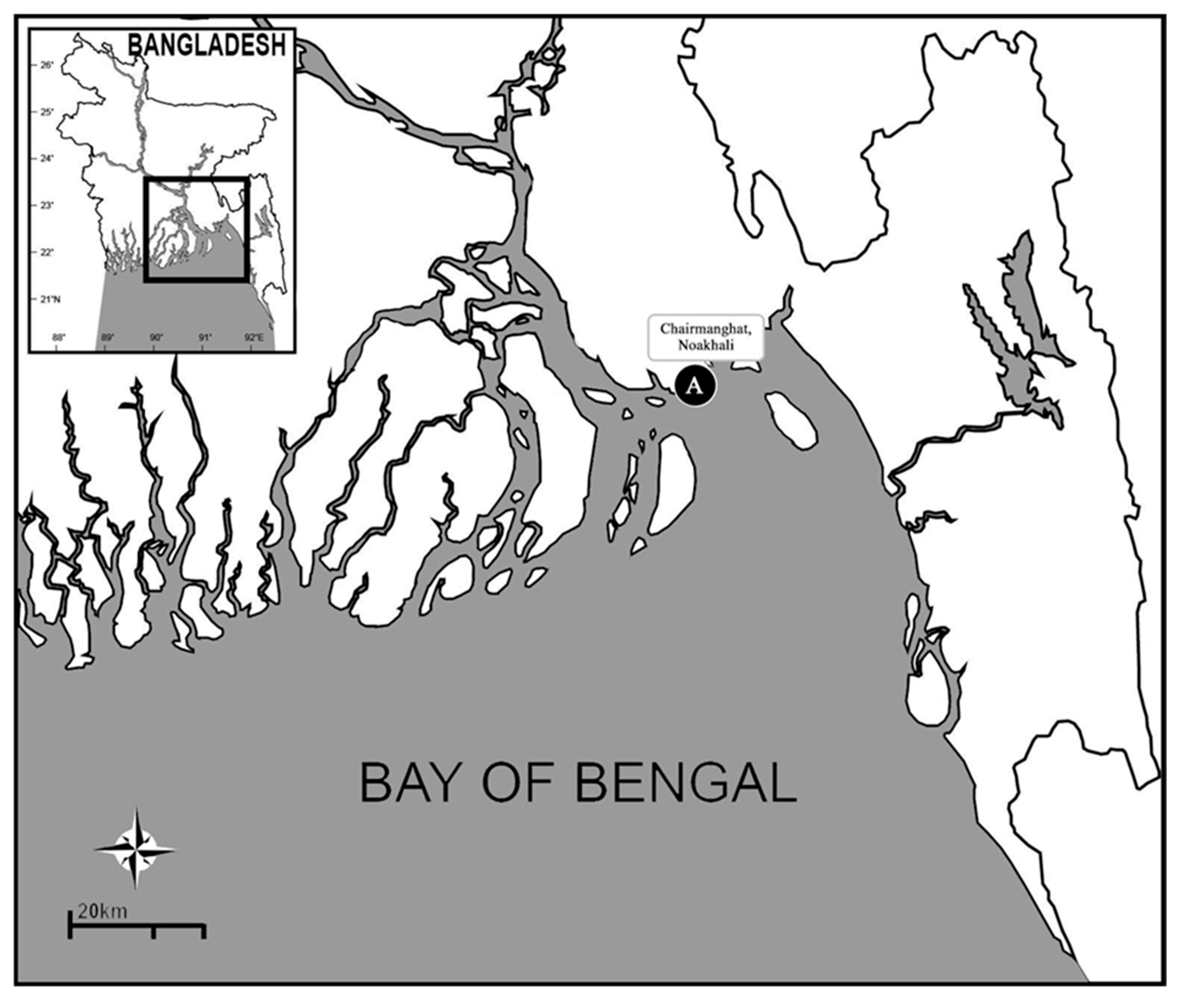
Sediment samples were collected during April 2015 from Chairman Ghat (22°30′48.3876″ N, 91°5′6.6078″ E, Noakhali, Chittagong division), using a hand-held corer with a depth penetration of 10 cm (Figure 1). The collected samples were washed through a 0.5 mm mesh hand sieve, and polychaetes retained on the sieve were placed into plastic vials and fixed with 5% formalin in the field. After 2 days, the samples were washed with freshwater and transferred to 70% ethyl alcohol for further examination [17].
Material was examined using stereo (Motic 6.5x–50X Zoom Stereo) and compound (Carl Zeiss, Oberkochen, Germany) microscopes. Scanning electron microscope (SEM) observations were made with a Zeiss EVO LS15 SEM with a Robinson Backscatter Detector after critical-point drying and coating with 20 nm gold at the Australian Museum [17]. Specimens were photographed using a light microscope (Leica MZ16, Leica Microsystems, Wetzlar, Germany ) and Spot flex 15.2 (Leica Microsystems, Wetzlar, Germany ) with a camera attached. In some cases, the material was stained with methylene blue to increase the resolution of diagnostic characters. All material examined was deposited at the Australian Museum, Sydney (AM).
3. Results
3.1. Systematics
3.2. Material Examined
The holotype was AM W.48048, Bangladesh, Chairman Ghat, Noakhali, 22°30′48.3876″ N, 91°5′6.6078″ E; mid-intertidal zone of the saltmarsh, a tributary of Meghna Estuary; collected by Hossain, M.B, 3 April 2015. The paratypes were AM W.48049, and AM W.49884 (MI1726 and MI1727), as well as AMW.49885 (MI1728 and MI1729) mounted for SEM.
3.3. Generic Identification
This species has been placed into the genus Glycera based on its overall morphological similarities with other species of Glycera, including the presence of different types of dense papillae on their proboscis. The genus Glycera Lamarck, 1818 [15] can easily be identified from other genera by the following unique characters [2,15]: acutely pointed, usually ringed prostomium with four terminal tentacles; and a long, eversible, club-like proboscis, provided with four hooked horny jaws and accessory lateral ailerons. The ailerons possess a more complicated structure with outer and inner rami, and sometimes an interramal plate. Parapodia have two anterior lobes with cirri and one or two posterior lobes, as well as the ventral chaetae compound and dorsal capillary chaetae.
3.4. Diagnosis
The salient features of the new species are (i) the presence of three types of proboscideal papillae—type 1 papillae (main type), which are conical with three transverse ridges; type 2 papillae, which are conical with straight, median, longitudinal ridges; and type 3 papillae, which are round, shorter, and broader, with straight, median, longitudinal ridges; (ii) Y-shaped ailerons with gently incised triangular bases; and (iii) digitiform noto- and neuropodial lobes of almost equal size in the mid-body and long ventral cirri at the posterior end.
3.5. Description
3.5.1. Holotype
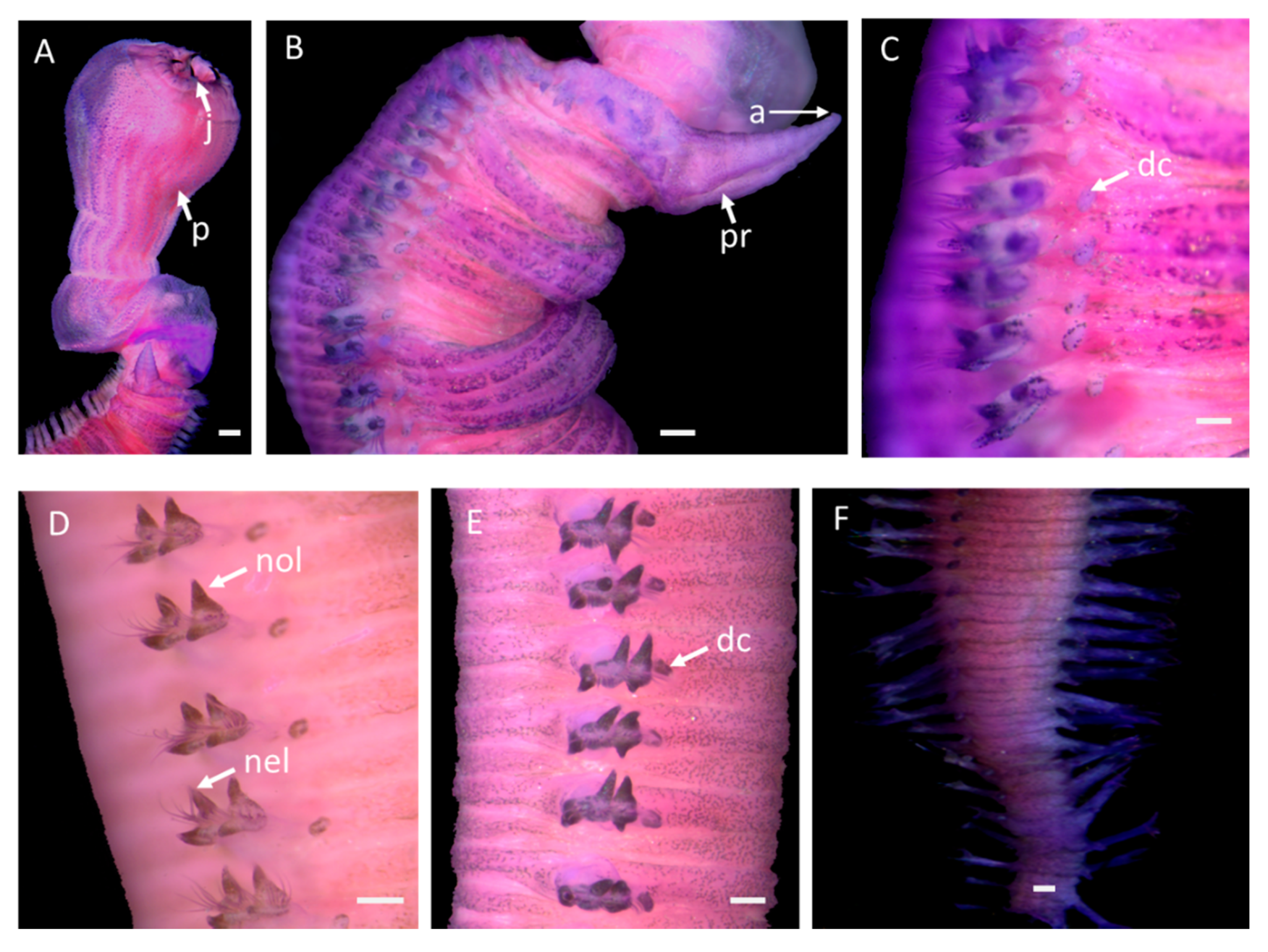
The holotype has an incomplete, cylindrical body, which is elongated and tapered at both ends (Figure 2A,B). The body reaches up to 42 mm long, with up to 158 segments, and has a width of 2.2 mm in the middle part of the body; preserved specimens in alcohol are whitish with numerous scattered small black pigmented spots (Figure 2C,D). The body segment is biannulate (Figure 2E), and the anterior annulus slightly shorter than posterior annulus.
The prostomium is conical, pointed, and distinctly separated into about ten rings; the terminal ring has four antennae, and no nuchal organs or eyes (Figure 2A,B and Figure 3A,C).
The parapodia of the first two segments are uniramous, with a prechaetal and a postchaetal lobe, while the subsequent parapodia are biramous (Figure 2B and Figure 3D). There are two unequal, triangular to digitiform, prechaetal and postchaetal anterior lobes, and lobes of similar length in mid-body (Figure 2C, Figure 3D, and Figure 4A,D). Knob-like dorsal cirri from the second parapodium are inserted most clearly at the base of the anterior parapodia, as well as on the body wall far above parapodial base in the mid-body, and again near the posterior base (Figure 2D–F). Ventral cirri are not distinct in the anterior part, but are well-developed in the posterior part (Figure 4F). Retractile branchiae are present at chaetigers 27–31. Notochaetae slender capillaries with one margin covered with spines or hairs (Figure 4A,C). There are neurochaetae homogomph spinigers (based on SEM) (Figure 4B), as well as a pygidium with a terminal pair of slender, elongated cirri (Figure 3G).
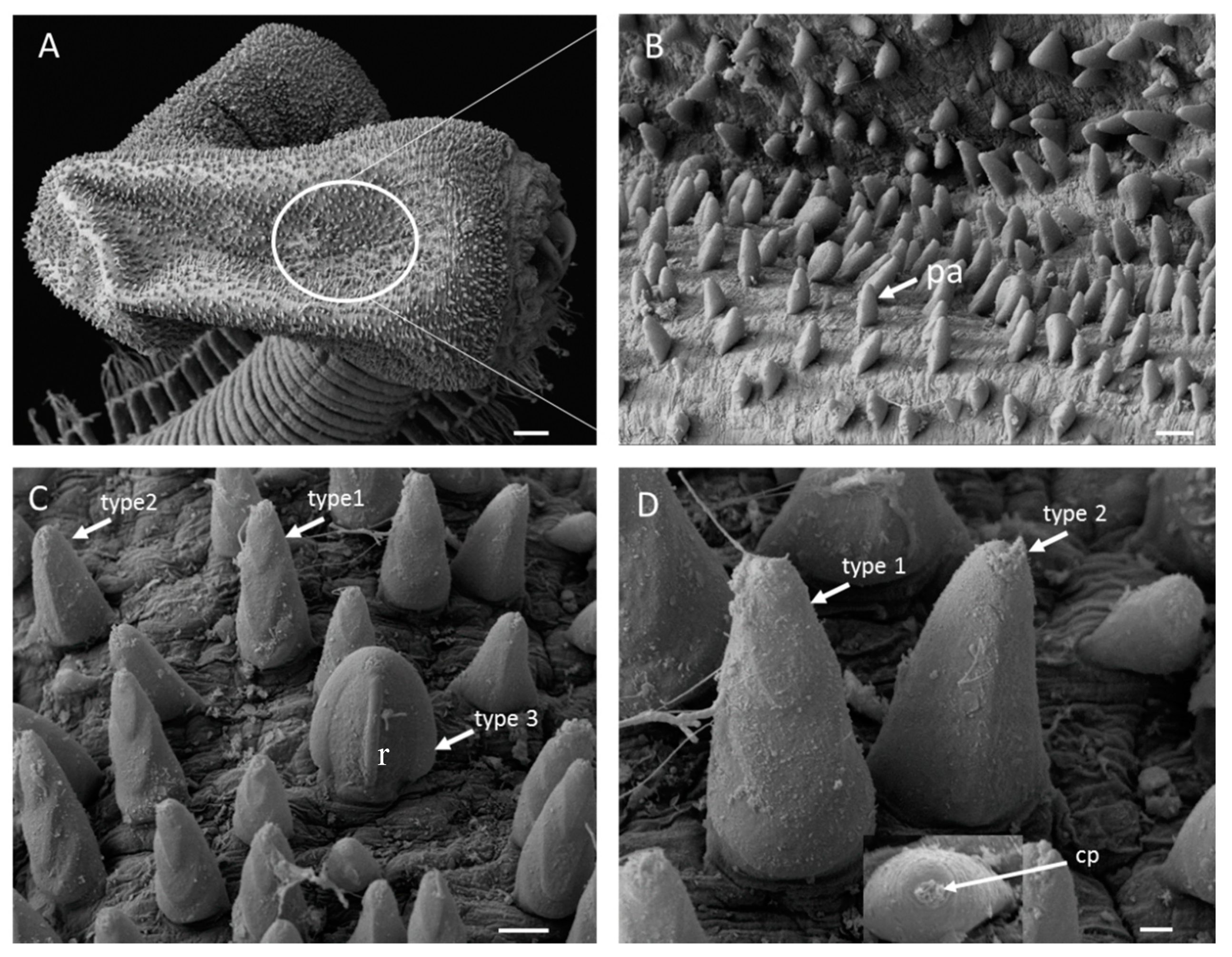
The proboscis is very long, equal to 28 segments, bell-shaped, and densely covered with papillae, which are arranged in distinct longitudinal rows (Figure 2A and Figure 3A,B). The papillae consist of three types: (1) numerous, conical papillae with three “V”- shaped ridges on posterior surface from top to bottom (Figure 5C,D); (2) a few long, conical papillae, with one straight, median longitudinal ridge (Figure 5C,D); and (3) very few, slightly shorter and broader, rounded papillae with a single very distinct median longitudinal ridge (Figure 5C,D). Among the three types, type 2 is the longest. All papillae have small ciliated pores (Figure 5D) and are smooth anteriorly. The terminal part of proboscis has four black hook-shaped jaws and accessory “Y”- shaped ailerons with gently incised triangular bases (Figure 3B).
3.5.2. Etymology
The new species Glycera sheikhmujibi n. sp. was named in memory of Bangabandhu Sheikh Mujibur Rahman (shortened as Sheikh Mujib; 1920–1975), founder of Bangladesh. Sheikh Mujib was president (in absentia from 17 April 1971 to 12 January 1972), prime minister (12 January 1972 to 24 January 1975), and then president again of Bangladesh (25 January 1975 to 15 August 1975) in the early to mid-70s of the last century. He made an undeniable and significant contribution to the emergence of independent Bangladesh. He inspired the people of East Pakistan to stand up against the oppression of the Pakistani ruler and fight for the independence of the country with his solid, dedicated, and visionary leadership. After independence, he realized that the growth and prosperity of the country would have not been possible without proper research in different fields of science, and hence he established many research organizations, including BARC (Bangladesh Agricultural Research Council) and BCSIR (Bangladesh Council of Scientific and Industrial Research). While he was in power, he invested his wholehearted efforts building a "Sonar Bangla" to ensure peace and prosperity for all. Unfortunately, he was assassinated, along with most of his family members, on 15 of August 1975 by a group of disgruntled army officials. This year (17 March 2020 to 17 March 2021), Bangladesh is celebrating the “Mujib Year”, on the occasion of the 100th birth anniversary of Sheikh Mujibur Rahman. The United Nations Educational, Scientific and Cultural Organisation (UNESCO) has also decided to jointly celebrate the “Mujib Year” with Bangladesh at its 40th General Assembly.
3.6. Distribution, Ecology, and Habitat
Glycera sheikhmujibi n. sp. is one of 11 species in the genus Glycera distributed in the Bay of Bengal region, and is the second species from Bangladeshi coastal waters. Currently, it is only known from the type locality on the central coast of Bangladesh; however, increased sample collection from other parts of the coastline might extend its distribution range. Sympatric species include Nephtys bangladeshi, Lumbrineris spp., Capitella spp., Goniada spp., Nereis spp., Magelona spp., and Naidadae, as well as the crustacean groups Ocypodidae, Palaemonidae, Gammaridae, and Harpecticoida. The species was collected from the muddy saltmarsh zone (intertidal zone), with a water depth range of 0.5 to 1.0 m during high tide. The zone is densely covered with the grass Spartina spp. and connected to the Meghna River Estuary, which falls to the Bay of Bengal near Hatiya and Swandwip islands. The average salinity, dissolved oxygen, pH, alkalinity, and temperature was 6 ppt, 9.15 ppm, 7.72, 180 ppm, and 29 °C, respectively.
4. Discussion
The main diagnostic characteristics for the identification of glycerid species include shape and number of pre- and post-chaetal lobes, presence or absence of branchiae, shape of the aileron, and the structure of proboscidial papillae [2]. However, Fiege and Böggemann [23] and Rizzo et al. [1] found that parapodial lobes and branchiae are not reliable characters, because branchiae are retractable in some species, and the shape (size) and number of pre- and post-chaetal lobes are difficult to evaluate for some species. Therefore, they suggested the proboscidial papillae and ailerons to be the most reliable characters for the identification of species of Glycera. Glycera sheikhmujibi n. sp. can easily be distinguished from all other species of Glycera by the presence of three distinct types and shapes of proboscidial papillae (Table 1).
Glycera sheikhmujibi n. sp. shares no diagnostic characteristics with either Glycera lancadivae, the only known species from Bangladesh [12], or fragments of another glycerid described by Muir and Hossain [16]; however, it clearly differs in many other aspects, especially in the form of three distinct types of proboscidial papillae. Again, Böggemann [2] mentioned that G. lancadivae is a nomen dubium, and it is almost similar to G. brevicirris. Glycera sheikhmujibi n. sp. seems to resemble Glycera nicobarica Grube, 1867 [32] and Glycera macintoshi Grube, 1877 [33] in the shape of ailerons, parapodial lobes, and types and shapes of the proboscidial papillae (Table 1). However, G. nicobarica and G. macintoshi have only two different types of papillae: G. nicobarica possesses few ovate papillae without ridges, and numerous leaf-like ones with five to six “U”-shaped ridges, and G. macintoshi has conical proboscidial papillae with three transverse ridges. Recently, G. nicobarica has been synonymized with Glycera unicornis Lamarck, 1818 by Read [24]. It has been argued that the proboscidial papillae may vary due to preservation, sample preparation, and development of papillae; however, for the sample collection and preservation of G. sheikhmujibi n. sp., standard procedures were followed.
In addition, G. sheikhmujibi n. sp. differs from G. macintoshi by the presence of two equal triangular pre- and postchaetal lobes, whereas shorter, rounded neuropodial postchaetal lobes are present in G. macintoshi. Glycera. embranchiata Krishnamoorthi, 1962 [26] (known from India) is a nomen dubium, as there is no description of the species in the original report, and G. rutilans Grube, 1877 [33] (known from Sri Lanka) is nomen nudum, as reported in Böggemann [2]. Glycera convoluta, G. longipinnis, G. rouxii, and Glycinde oligodon, all reported from the Bay of Bengal region, have not been accepted by Böggemann [2], as G. convoluta is a junior synonym of G. tridactyla, G. longipinnis is a junior synonym of G. sphyrabrancha, G. rouxii is a junior synonym of G. unicornis, and Glycinde oligodon belongs to the family Goniadidae. Although Glycera tesselata Grube, 1863 [31] is a good species and is accepted by Böggemann [2] and Read [24], it is poorly described in the original description.
The first procedure in any ecological work or applied research with organisms is an exercise in taxonomy. Taxonomy provides the fundamental understanding about the components of biodiversity, which is badly needed for effective decision-making regarding conservation, management, and sustainable use of the studied organisms. In addition, the loss of biodiversity due to human activities and climate change should be of major concern to everyone, because it threatens the functioning of an ecosystem. Despite this, there is very little information on the taxonomy of polychaetes from Bangladeshi coastal waters compared with those of neighbouring countries. To date, only thirty species have been identified from Bangladesh, which is a very low number compared to known polychaete species (~10,000) in the world. With a diverse coastline of about 720 km, it is hoped that the number of polychaete species will be increased with further studies.
Key to the species of genus Glycera from the Bay of Bengal region (modified from Muir and Hossain [15]) is as follows:
- Proboscideal papillae do not have a terminal fingernail structure 2– Proboscideal papillae have a terminal fingernail structure 6
- There is one postchaetal lobe in all parapodia 3– There are two postchaetal lobes (at least) on the mid-body parapodia 5
- Mid-body, the notopodial prechaetal lobes are shorter than the neuropodial lobes, and branchiae are absentGlycera lapidum Quatrefages, 1866 [24].– Mid-body, the prechaetal lobes are about same length or longer than the notopodial lobes. Branchiae are present or absent 4
- The proboscideal papillae are digitiform and without ridges, ailerons have deeply incised bases, and simple digitiform branchiae are situated termino-dorsally on the parapodiaGlycera sphyrabrancha Schmarda, 1861 [12].– Conical proboscideal papillae with 5–20 transverse ridges, ailerons have slightly arched bases, and branchiae are absentGlycera oxycephala Ehlers, 1887 [25].
- Ailerons have gently incised bases; long, mid-body postchaetal lobes are digitiform and of about equal length; three types of proboscideal papillae, with the main type having fewer than three ridges; and branchiae are absent Glycera sheikhmujibi n. sp.– Ailerons have an interamal plate and triangular bases; mid-body parapodia have slender, triangular, notopodial and distinctly shorter, rounded neuropodial postchaetal lobes. The retractile branchiae are situated medially on anterior side of parapodia 7
- Digitiform proboscideal papillae have a straight, median, longitudinal ridgeGlycera tesselata Grube, 1863 [26].– Digitiform proboscideal papillae have 6–20 transverse ridgesGlycera brevicirris Grube, 1870 [13].
- Parapodia have slender, triangular, notopodial and distinctly shorter, rounded, neuropodial postchaetal lobes; and branchiae are simple, digitiform, and retractileGlycera nicobarica Grube, 1867 [18].– Parapodia have two slender, triangular postchaetal lobes of about the same length, or notopodial lobes that are only slightly longer than the neuropodial lobes; branchiae are digitiform and retractile with 1–2 rami Glycera unicornis Lamarck, 1818 [14].
- The mid-body parapodia have two slender. triangular postchaetal lobes of about the same length 9– The mid-body parapodia have slender, triangular notopodial and shorter, more or less rounded, neuropodial postchaetal lobes 10
- Parapodia do not have branchiae Glycera onomichiensis Izuka, 1912 [27].– There are 1–5 digitiform branchial rami situated dorsally on the parapodial basesGlycera cinnamomea Grube, 1874 [19].
- In mid-body and posterior parapodia, the neuropodial postchaetal lobes are more or less rounded. Simple digitiform branchiae are situated termino-dorsally on the parapodia 11– In the posterior parapodia are neuropodial postchaetal lobes as long as the notopodial lobes, and equally slender and triangular. Simple digitiform branchiae are situated medio-dorsally on the parapodia Glycera posterobranchia Hoagland, 1920 [28].
- All biramous parapodia have two postchaetal lobes. Proboscideal papillae have long, medium, or short stalks. 12– In the anterior parapodia, there is only one, medially inserted, slender triangular postchaetal lobe. The proboscideal papillae have short stalksGlycera macrobranchia Moore, 1911 [29].
- Proboscideal papillae have long stalks 13– Proboscideal papillae have medium-length or short stalks 14
- There are stalk without ridges and ailerons with pointed triangular basesGlycera alba Müller, 1776 [30].– There are stalks with numerous ridges and ailerons with triangular basesGlycera natalensis Day, 1967 [31].
- The proboscideal papillae have short stalks, the prostomium consists of about 11–15 rings, and ailerons have triangular basesGlycera tridactyla Schmarda, 1861 [12].– The proboscideal papillae have medium-length stalks, the prostomium consists of about 19–28 rings, and ailerons have pointed triangular basesGlycera africana Arwidsson, 1899 [32].
Author Contributions
Conceptualization: M.B.H.; methodology: M.B.H.; formal analysis: M.B.H.; investigation: M.B.H.; writing—original draft preparation: M.B.H.; writing—review and editing: P.H.; visualization: P.H.; resources: P.H.; supervision: P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Thanks are due to AMRI (Australian Museum Research Institute) for providing lab space, facilities, and access to the library. We would like to thank Sue Lindsay, who helped greatly with the SEM photographs. Field and lab assistance was provided by some students, namely Amir, Jewel, Tanmay, and Rasel, and the Milonof Fisheries and Marine Science Department are acknowledged.
Conflicts of Interest
The authors have no conflict of interest.
References
- Rizzo, A.E.; Steiner, T.M.; Amaral, A.C.Z. Glyceridae Grube 1850 (Annelida: Polychaeta) from southern and southeastern brazil, including a new species of Glycera. Biota Neotrop. 2007, 7, 41–59. [Google Scholar] [CrossRef] [Green Version]
- Böggemann, M. Revision of the Glyceridae Grube 1850 (Annelida: Polychaeta). Abh. Senckenb. Naturforsch Ges. 2002, 555, 1–249. [Google Scholar]
- Böggemann, M.; Bienhold, C.; Gaudron, S.M. A new species of Glyceridae (Annelida: “Polychaeta”) recovered from organic substrate experiments at cold seeps in the eastern Mediterranean Sea. Mar. Biodivers. 2012, 42, 47–54. [Google Scholar] [CrossRef]
- Böggemann, M. Glyceriformia (Annelida) of the abyssal SW Atlantic and additional material from the SE Atlantic. Mar. Biodivers. 2016, 46, 227–241. [Google Scholar] [CrossRef]
- Choi, H.K.; Jung, T.W.; Yoon, S.M. A New Record of Glycerid Polychaete, Glycera fallax (Polychaeta: Glyceridae) from Korea. Korean J. Environ. Biol. 2015, 33, 274–278. [Google Scholar] [CrossRef]
- Sotomayor-Garcia, A.; Rueda, J.L.; Sánchez-Guillamón, O.; Vázquez, J.T.; Palomino, D.; Fernández-Salas, L.M.; López-González, N.; González-Porto, M.; Urra, J.; Santana-Casiano, J.M.; et al. Geomorphic features, main habitats and associated biota on and around the newly formed Tagoro submarine volcano, Canary Islands. In Seafloor Geomorphology as Benthic Habitat; Harris, P.T., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 51, pp. 835–846. [Google Scholar]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Ocean. Mar. Biol. Ann. Rev. 1979, 17, 193–284. [Google Scholar]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S. Macrobenthos of Intertidal zone of Halishahar Coast, Chittagong. Ph.D. Thesis, Department of Zoology, University of Chittagong, Chittagong, Bangladesh, 1990. [Google Scholar]
- Belaluzzaman, A.M. Ecology of the intertidal macrobenthic fauna in Cox’s Bazar coastal area, Bangladesh. Master’s Thesis, Institute of Marine Sciences and Fisheries, University of Chittagong, Chittagong, Bangladesh, 1995. [Google Scholar]
- Hossain, M.J.; Sarker, M.J.; Uddin, M.N.; Islam, A.; Tumpa, I.J.; Hossain, Z. Macrobenthos presence in the estuarine waters of the Meghna River, Ramghati, Laksmipur, Bangladesh. World Appl. Sci. J. 2018, 36, 598–604. [Google Scholar]
- Pramanik, M.N.; Chowdhury, S.H.; Kabir, S.M.H. Annelida. In Encyclopedia of Flora and Fauna of Bangladesh (Annelida, Echinodermata, Acanthocephala and Minor Phyla); Ahmad, M., Ahmed, A.T.A., Rahman, A.K.A., Ahmed, Z.U., Begum, Z.N.T., Hassan, M.A., Khondker, M., Eds.; Asiatic Society of Bangladesh: Dhaka, Bangladesh, 2009; Volume 16, pp. 1–91. [Google Scholar]
- Schmarda, L.K. Neue wirbellose Thiere beobachtet und gesammelt auf einer Reise un die Erdr 1853 bis 1857: Erster Band (zweite halfte) Turbellarian, Rotatorien un Anneliden; Wilhelm Engelmann: Leipzig, Germany, 1861; pp. 1–164. [Google Scholar]
- Grube, A.E. Bemerkungen uber die familie der Glycereen. Schlesisch. Gesellsch. fur vaterlandisch. cultur Breslau Jahresber 1870, 47, 56–68. [Google Scholar]
- Lamarck, J.B. Histoire naturelle des Animaux sans Vertèbres, préséntant les caractères généraux et particuliers de ces animaux, leur distribution, leurs classes, leurs familles, leurs genres, et la citation des principales espèces qui s’y rapportent; precedes d’une Introduction offrant la determination des caracteres essentiels de l‘Animal, sa distinction du vegetal et desautres corps naturels, enfin; l’Exposition des Principes fondamentaux de la Zoologie: Paris, France, 1818; Volume 5, p. 612. [Google Scholar]
- Muir, A.I.; Hossain, M.M.M. The intertidal polychaete (Annelida) fauna of the Sitakunda coast (Chittagong, Bangladesh), with notes on the Capitellidae, Glyceridae, Lumbrineridae, Nephtyidae, Nereididae and Phyllodocidae of the “Northern Bay of Bengal Ecoregion”. ZooKeys 2014, 419, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.B.; Hutchings, P. Nephtys bangladeshi n. sp., a new species of Nephtyidae (Annelida: Phyllodocida) from Bangladesh coastal waters. Zootaxa 2016, 4079, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Flura, M.A.; Akhery, N.; Mohosena, B.T.; Masud, H.K. Physico-chemical and biological properties of water from the river Meghna, Bangladesh. Int. J. Fish. Aquac. Stud. 2016, 4, 161–165. [Google Scholar]
- Sharif, A.S.M.; Bakar, M.A.; Bhuyan, M.S. Assessment of water quality of the lower Meghna river estuary using multivariate analyses and RPI. Intl. J. Chem. Pharm. Technol. 2017, 2, 57–73. [Google Scholar]
- Syed, Z.H.; Choi, G.; Byeon, S. A numerical approach to predict water levels in ungauged regions—Case study of the meghna river estuary, Bangladesh. Water 2018, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, M.M.H.; Hasan, M.M.; Bisshas, S.; Hossain, A.A.; Biswas, T.K. Fish biodiversity and their present conservation status in the Meghna River of Bangladesh. Int. J. Fish. Aquac. Stud. 2017, 5, 446–455. [Google Scholar]
- Grube, A.E. Die Familien der Anneliden. Archiv für Naturgeschichte Berlin 1850, 16, 249–364. [Google Scholar]
- Fiege, D.; Boggemann, M. Scanning electron microscopy of the proboscidial papillae of some European Glyceridae. Bull. Mar. Sci. 1997, 60, 559–563. [Google Scholar]
- World Polychaeta Database. Accessed through: World Register of Marine Species. Available online: http://marinespecies.org/aphia.php?p=taxdetails&id=129296 (accessed on 30 April 2020).
- Grube, A.E. Descriptiones Annulatorum novorum mare Ceylonicum habitantium ab honoratissimo Holdsworth collectorum. Proceed. Zool. Soci. London 1874, 325–329. [Google Scholar]
- Krishnamoorthi, B. Salinity tolerance and volume regulation in four species of polychaetes. Proceed. Indian Acad. Sci. 1962, 56, 363–371. [Google Scholar]
- Grube, A.E. Über eine Sammlung von wirbellosen Seethieren, welche Herr Dr. Eugen Reimann dem hiesigen zoologischen Museum zum Geschenk gemacht. Jahres-Bericht der Schlesiche Gesellschaft fuer vaterlandische Cultur. Breslau 1877, 54, 48–51. [Google Scholar]
- Hoagland, R.A. Polychaetous annelids collected by the United States fisheries steamer Albatross during the Philippine expedition of 1907–1909. Bull. U.S. Nat. Mus. 1920, 100, 603–635. [Google Scholar]
- McIntosh, W.C. Report on the Annelida Polychaeta collected by H.M.S. Challenger during the years 1873–1876. Reports on the Scientific Results of the Voyage of HMS “Challenger”. Zoology 1885, 12, 1–554. [Google Scholar]
- Grube, A.E. Annulata Semperiana. Beiträge zur Kenntniss der Annelidenfauna der Philippinen nach den von Herrn Prof. Semper mitgebrachten Sammlungen. Mémoires l’Académie Impériale des Sciences de St. Pétersbourg 1878, 25, 1–300. [Google Scholar]
- Grube, A.E. Beschreibung neuer oder wenig bekannter Anneliden. Sechster Beitrag; Archiv für Naturgeschichte: Berlin, Germany, 1863; Volume 29, pp. 37–69. [Google Scholar]
- Grube, A.E. Reise der Österreichischen Frégatte Novara um die Erde inden Jahren 1857, 1858, 1859. Unter den Befehlen des Commodore B. von Wüllerstorf-Urbair. Novara-Expedition. Ser. Zoologischer Theil. 1867, 2, 1–46. [Google Scholar]
- Monatsberichte der Könglich Preussischen Akademie der Wissenschaften zu. Available online: https://www.biodiversitylibrary.org/page/35723826#page/581/mode/1up (accessed on 26 May 2020).
Figure 1.
Map showing the collection area (A). Source: [17].
Figure 1.
Map showing the collection area (A). Source: [17].

Figure 2.
Glycera sheikhmujibi n. sp.: (A) anterior dorsal view, (B) anterior lateral view, (C) lateral view of anterior chaetigers, (D) mid-body chaetigers, (E) lower mid-body chaetigers, (F) posterior chaetigers. j; jaw, a: antenna, p: proboscis, pr: prostomium, dc: dorsal cirri, nol = notopodial lobe, nel = neuropodial lobe; scale bars: 0.1 mm.
Figure 2.
Glycera sheikhmujibi n. sp.: (A) anterior dorsal view, (B) anterior lateral view, (C) lateral view of anterior chaetigers, (D) mid-body chaetigers, (E) lower mid-body chaetigers, (F) posterior chaetigers. j; jaw, a: antenna, p: proboscis, pr: prostomium, dc: dorsal cirri, nol = notopodial lobe, nel = neuropodial lobe; scale bars: 0.1 mm.

Figure 3.
Glycera sheikhmujibi n. sp. scanning electron microscope (SEM) images of the (A) anterior lateral view, (B) terminal ring of the jaws, (C) enlarged view of prostomium, (D) anterior chaetigers, (E) mid-body chaetigers, (F) lower mid-body chaetigers, and (G) posterior chaetigers. ai: aileron, j: jaw, a: antenna, vc: ventral ciri, nol= notopodial lobe, nel= neuropodial lobe, ac: annul ciri; scale bars: A and B = 200 µm, C and D= 100 µm, E = 20 µm, F = 10 µm, G = 20 µm.
Figure 3.
Glycera sheikhmujibi n. sp. scanning electron microscope (SEM) images of the (A) anterior lateral view, (B) terminal ring of the jaws, (C) enlarged view of prostomium, (D) anterior chaetigers, (E) mid-body chaetigers, (F) lower mid-body chaetigers, and (G) posterior chaetigers. ai: aileron, j: jaw, a: antenna, vc: ventral ciri, nol= notopodial lobe, nel= neuropodial lobe, ac: annul ciri; scale bars: A and B = 200 µm, C and D= 100 µm, E = 20 µm, F = 10 µm, G = 20 µm.

Figure 4.
Glycera sheikhmujibi n. sp. SEM images of the (A) anterior parapodium, (B) compound neurochatae, (C) capillary notochaetae, (D) mid-body parapodia, (E) posterior parapodium, and (F) posterior end parapodia. h: homogomph spinigers, nol: notopodial lobe, nel: neuropodial lobe, s: spines, vc: ventral ciri. Scale bars: A, B, C, and E = 10 µm; D = 20 µm; F = 30 µm.
Figure 4.
Glycera sheikhmujibi n. sp. SEM images of the (A) anterior parapodium, (B) compound neurochatae, (C) capillary notochaetae, (D) mid-body parapodia, (E) posterior parapodium, and (F) posterior end parapodia. h: homogomph spinigers, nol: notopodial lobe, nel: neuropodial lobe, s: spines, vc: ventral ciri. Scale bars: A, B, C, and E = 10 µm; D = 20 µm; F = 30 µm.

Figure 5.
Glycera sheikhmujibi n. sp. SEM images of the (A) proboscis; (B) enlarged view of the proboscideal papillae; (C) posterior side view of type 1, type 2, and type 3 papillae; and (D) enlarged view of type 1and type 2 papillae. cp: ciliated pore, pa: papillae, r: ridge. Scale bars: A = 100 µm, B = 20 µm, C = 10 µm, and D = 2 µm.
Figure 5.
Glycera sheikhmujibi n. sp. SEM images of the (A) proboscis; (B) enlarged view of the proboscideal papillae; (C) posterior side view of type 1, type 2, and type 3 papillae; and (D) enlarged view of type 1and type 2 papillae. cp: ciliated pore, pa: papillae, r: ridge. Scale bars: A = 100 µm, B = 20 µm, C = 10 µm, and D = 2 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of Glycera recorded from near the Bay of Bengal Region, with their comparative characteristics, as per World Register of Maine Species, WoRMS [24].
Table 1.
List of Glycera recorded from near the Bay of Bengal Region, with their comparative characteristics, as per World Register of Maine Species, WoRMS [24].
| Species | Type Locality | Data Source | Body Length, Width (Max) in mm, No. of Chaetigers (Holotype/Neotype) | Body Pigmentation | Prostomium Shape, No. of Rings, and No. of Prostomium Antenna | Proboscis Arrangement, and Type and Shape of Papillae |
|---|---|---|---|---|---|---|
| Glycera brevicirris Grube, 1870 [14] | Indonesia, Philippines, Indian Ocean | Based on Rizzo et al. [1] | 13 mm, 2 mm for 44 chaetigers (syntype) | Yellowish | Conical with 10 rings | Two types of papillae; type 1—numerous, digitiform, anteriorly smooth; and type 2—isolated, broader with longitudinal ridge |
| G. cinnamomea Grube, 1874 [25] | Sri Lanka, India | Based on original description | 94 mm, 3 mm at 75 chaetiger, 195 chaetigers (holotype) | White | Conical with 11 rings | Papillae short and digitiform |
| G. embranchiata Krishnamoorthi, 1962 [26] | India | Nomen dubium | - | - | ||
| G. lancadivae Schmarda, 1861 [13] | Sri Lanka | Based on original description | 50 mm, 3mm, 158 (holotype) | Yellowish-brown | Reversed cone, below the jaws. 16 larger | Hair-like papillae |
| G. manorae Fauvel, 1932 [27] | Manora Shoal, Karachi, India | Based on original description | 70 mm, 5 mm, n/a (holotype) | n/a | Acutely conical, with 10–12 rings and four tentacles | Small, cylindrical, unguiculate papillae |
| G. posterobranchia Hoagland, 1920 [28] | Marinduque Island, Philippines | Original | 75 mm, 3 mm, 90 (holotype) | Light brown | n/a, equal in length to first eight chaetigers | Two kinds of papillae: sucker-like and large blunt conical |
| G. rutilans Grube in McIntosh, 1885 [29] | Sri Lanka | Nomen nudum | ||||
| G. sagittariae McIntosh, 1885 [29] | Off Arrou Island, Madras coast, India | Based on original description | 110, 5, n/a | Not availavle (n/a) | n/a | Two types of papillae: short, globular or ovate; and long and slender without terminal nail- like appendage. |
| G. subaenea Grube, 1878 [30] | Philippines | Based on original description | 52 mm, 2.2 mm for 166 (holotype) segments | n/a | n/a | n/a |
| G. tesselata Grube, 1863 [31] | India | Based on original description | 48 mm, 6 mm, incomplete | n/a | Prostomium with 16–17 rings | n/a |
| Glycera sheikhmujibi n. sp. | Noakhali, Bangladesh | Based on present study | 42 mm and 2.2 mm for 158 segments | Whitish with black numerous black spot | Conical prostomium with 10–11 rings | Three types of papillae: type 1—numerous, digitiform with 2–3 ridges; type 2—isolated with one median straight ridge; and type 3—broader with a longitudinal ridge |
| Jaws and Aileron | Noto- & neuropodia; anteriror and posterior chaetigers | Branchiae | Chaetae | Remarks | ||
| Glycera brevicirris Grube, 1870 [14] | Aileron is triangular, with an elongated process on one side | Anterior parapodia uniramous with a pre- and postchaetal lobe where dorsal cirrus absent; posterior parapodia biramous with two triangular to digitiform prechaetal lobes | Absent | Parapodia with 2–4 simple capillary notochaetae and 5–13 spinigerous neurochaetae | ||
| G. cinnamomea Grube, 1874 [25] | n/a | n/a | ||||
| G. embranchiata Krishnamoorthi, 1962 [26] | Nomen dubium | Böggemann [2] reported as a nomen nudum | ||||
| G. lancadivae Schmarda, 1861 [13] | Jaws with large tooth and have a bent process | Parapodia biramous and tongue-like, but in the anterior segments cylindrical; dull, cone-shaped ventral cirrus in the first segments, pointed cone in the posterior segments | Branchiae absent in posterior segments | Few chaetae—curved and capillary, and partly spinigers | ||
| G. manorae Fauvel, 1932 [27] | Jaws obliquely truncated, aileron is triangular with an elongated process on one side | Parapodia with two sharp, triangular, equal anterior lobes and two similar but shorter blunt posterior lobes; dorsal cirrus elongated, knob near base of parapodia and ventral cirrus triangular | Retractile branchiae beginning at about the 17th foot | Dorsal capillary chaetae with narrow wings; ventral chaetae compound homogomph with finely serrated terminal piece. | ||
| G. posterobranchia Hoagland, 1920 [28] | Jaws with lateral appendages | Anterior parapodia with long, conical dorsal lobes; posterior parapodia elongated, divided into one dorsal and two ventral rami; dorsal cirrus a rounded tubercle and ventral cirrus similar shape to dorsal lobe | Branchiae begins at 25th chaetiger as small knobs at the dorsal base of parapodia | Two kinds: simple dorsal capillaries with finely serrated edges and ventral compounds with finely serrated blade | ||
| G. rutilans Grube in McIntosh, 1885 [29] | Nomen nudum | Böggemann [2] reported as a nomen nudum | ||||
| G. sagittariae Fauvel, 1932 [27] | Aileron with two long dagger-like processes | Parapodia with two equal elongated, tapering anterior lobes, and two equal, blunt, triangular posterior lobes; dorsal cirrus more or less remote | Present, simple and short beginning at 40th segment | n/a | ||
| G. subaenea Grube, 1878 [30] | n/a | Posterior parapodial lobes longer than the anterior ones; lower lobes triangular and wider than the upper ones, anterior lobes equally long, rounded | Branchiae present and positioned at the anterior wall of parapodium, separated into 2–3 fingerlike filaments, longer than ventral cirrus | n/a | ||
| G. tesselata Grube, 1863 [31] | n/a | n/a | n/a | n/a | ||
| Glycera sheikhmujibi n.sp. | Dark, hook-shaped jaws and ailerons with gently incised bases | Parapodia with digitiform prechaetal and postchaetal lobes; knob-like dorsal cirrus along the body and long ventral cirrus present posteriorly | Branchiae present, retractile, commencing from the 27th to 31st segments | 5–6 slender capillary notochaetae |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hossain, M.B.; Hutchings, P. Glycera sheikhmujibi n. sp. (Annelida: Polychaeta: Glyceridae): A New Species of Glyceridae from the Saltmarsh of Bangladesh. Diversity 2020, 12, 213. https://doi.org/10.3390/d12060213
AMA Style
Hossain MB, Hutchings P. Glycera sheikhmujibi n. sp. (Annelida: Polychaeta: Glyceridae): A New Species of Glyceridae from the Saltmarsh of Bangladesh. Diversity. 2020; 12(6):213. https://doi.org/10.3390/d12060213
Chicago/Turabian StyleHossain, M. Belal, and Pat Hutchings. 2020. "Glycera sheikhmujibi n. sp. (Annelida: Polychaeta: Glyceridae): A New Species of Glyceridae from the Saltmarsh of Bangladesh" Diversity 12, no. 6: 213. https://doi.org/10.3390/d12060213
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.