In Vitro Modeling of Reoxygenation Effects on mRNA and Protein Levels in Hypoxic Tumor Cells upon Entry into the Bloodstream

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies used for Western Blot
2.2. Cell Lines and Culture Conditions
2.3. Densitometric Analysis
2.4. Quantitative RT-PCR
3. Results
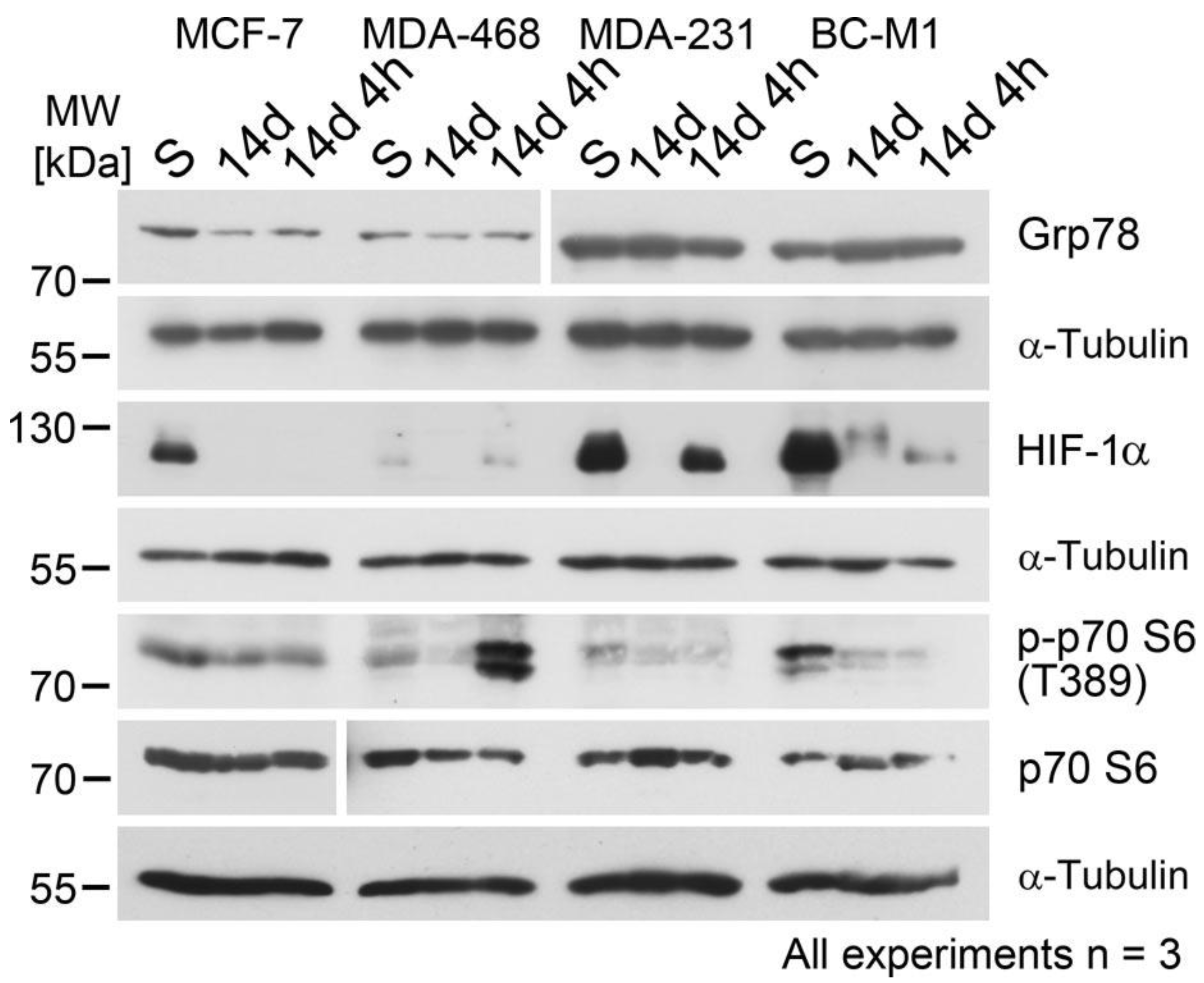
3.1. The Response of Hypoxia Response Proteins to Hypoxia and Reoxygenation
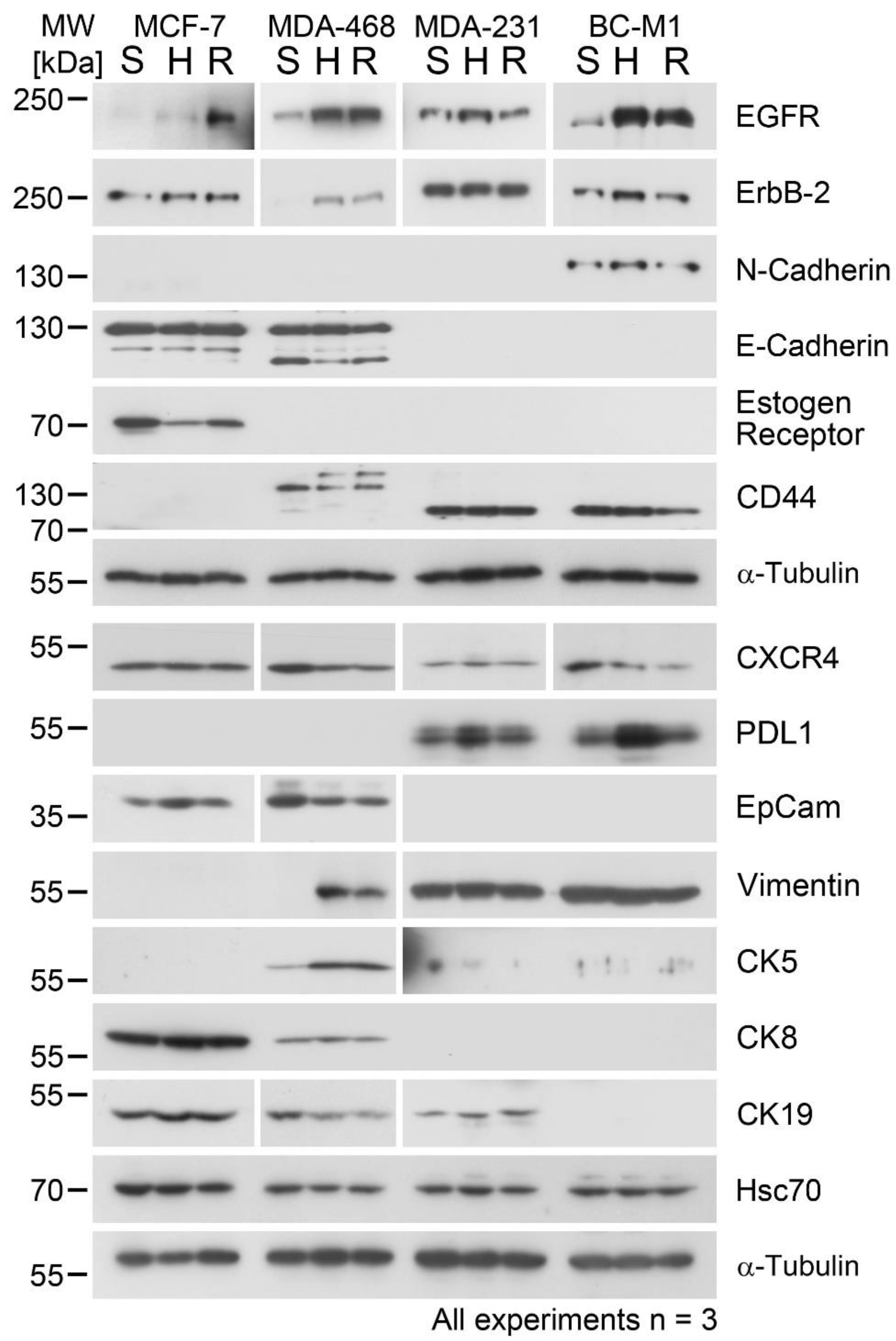
3.2. Cellular Responses to Hypoxia and Reoxygenation in Breast Cancer-Relevant Proteins
3.3. Changes of Epithelial Differentiation Marker Proteins under Hypoxia and Reoxygenation
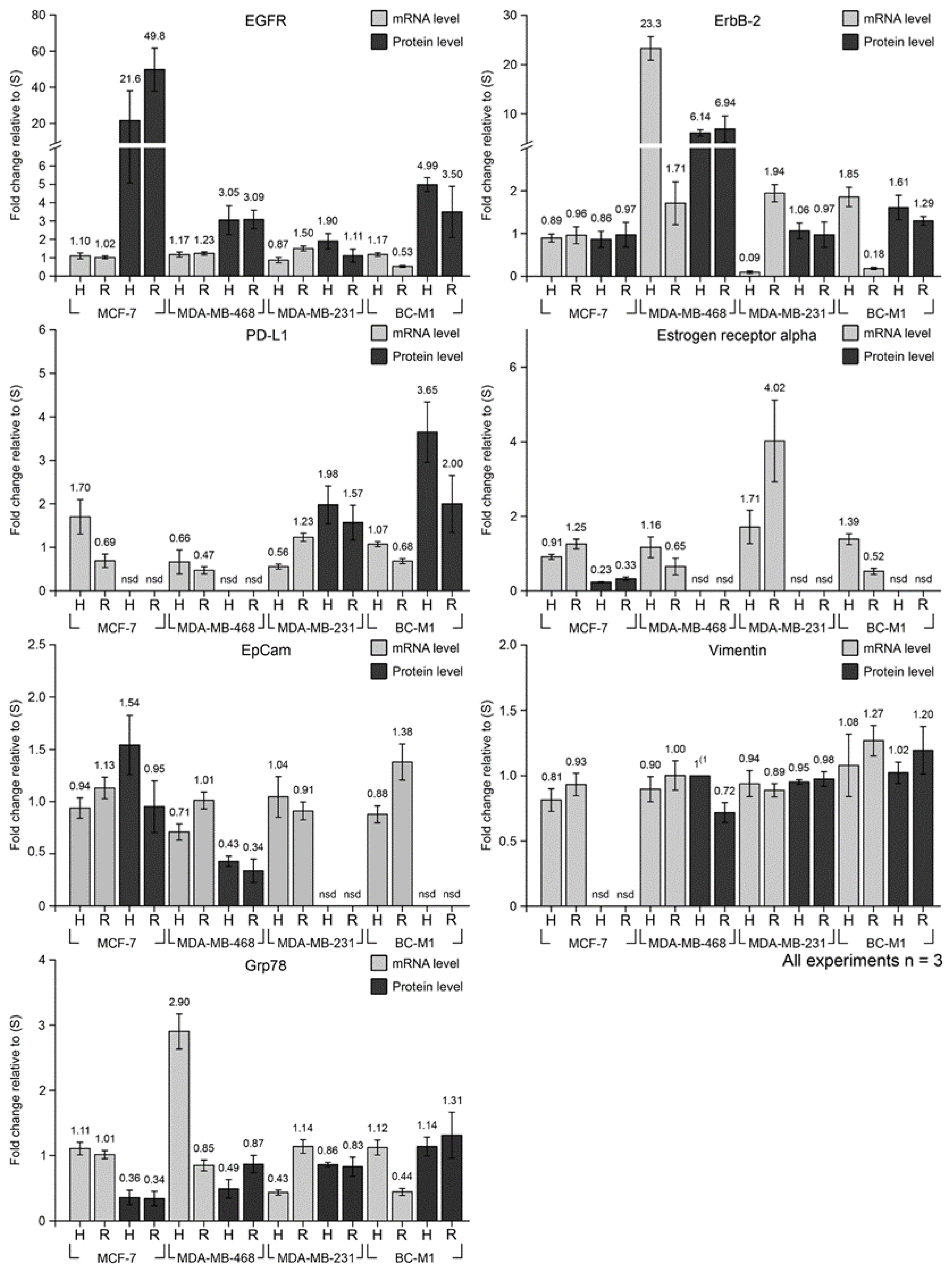
3.4. Cellular Responses to Hypoxia and Reoxygenation on mRNA Level
3.5. Discordance of mRNA and Protein Levels under Standard Conditions, Hypoxia, and Reoxygenation
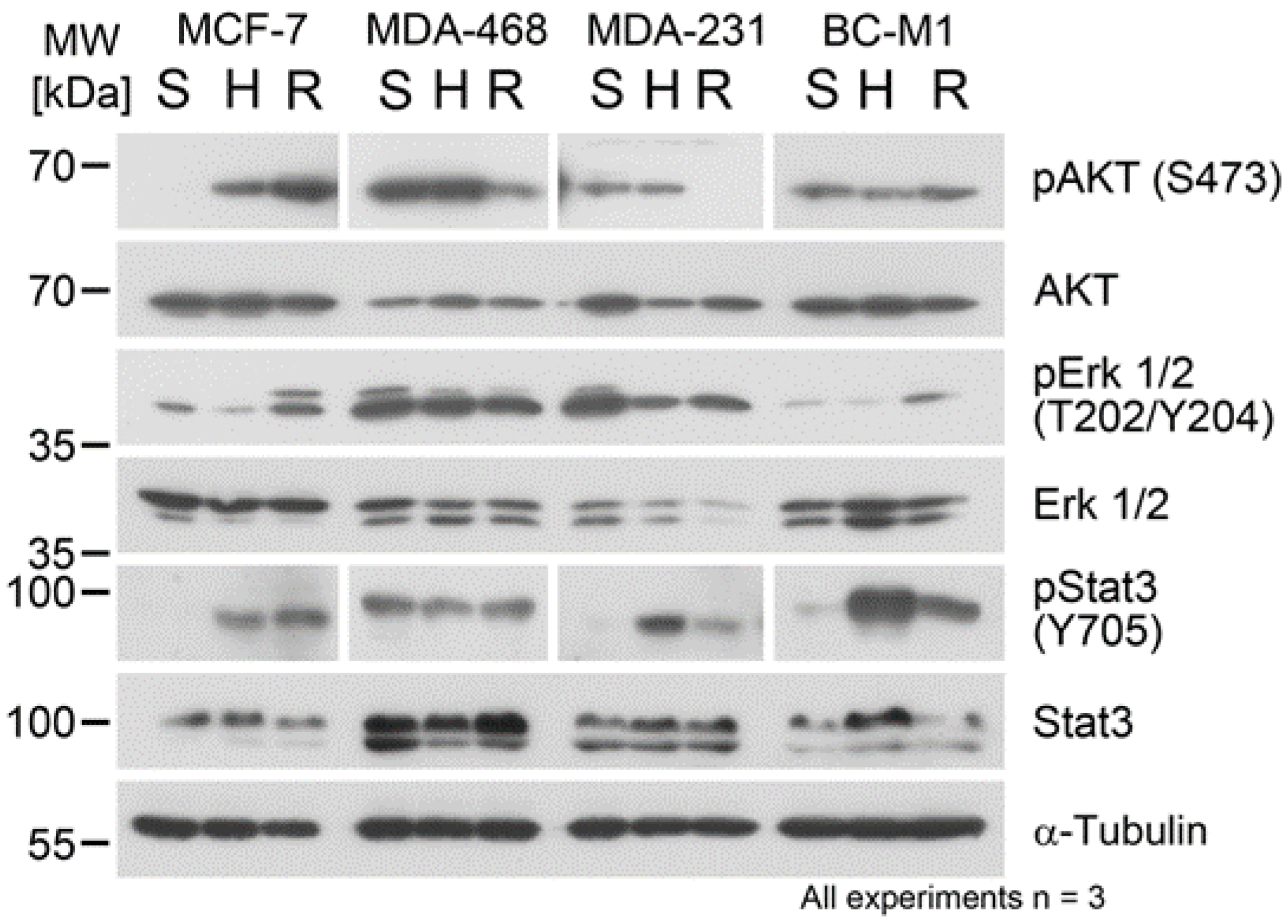
3.6. Hypoxia and Reoxygenation Affected the Protein Phosphorylation Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alix-Panabieres, C.; Pantel, K. Challenges in circulating tumour cell research. Nat. Rev. Cancer 2014, 14, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Pantel, K.; Alix-Panabieres, C.; Riethdorf, S. Cancer micrometastases. Nat. Rev. Clin. Oncol. 2009, 6, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Vogl, F.D.; Naume, B.; Janni, W.; Osborne, M.P.; Coombes, R.C.; Schlimok, G.; Diel, I.J.; Gerber, B.; Gebauer, G.; et al. A pooled analysis of bone marrow micrometastasis in breast cancer. N. Engl. J. Med. 2005, 353, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Kentenich, C.; Janni, W.; Hepp, F.; de Waal, J.; Willgeroth, F.; Sommer, H.; Pantel, K. Lack of effect of adjuvant chemotherapy on the elimination of single dormant tumor cells in bone marrow of high-risk breast cancer patients. J. Clin. Oncol. 2000, 18, 80–86. [Google Scholar] [CrossRef]
- Korsching, E.; Jeffrey, S.S.; Meinerz, W.; Decker, T.; Boecker, W.; Buerger, H. Basal carcinoma of the breast revisited: An old entity with new interpretations. J. Clin. Pathol. 2008, 61, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ridgway, L.D.; Wetzel, M.D.; Ngo, J.; Yin, W.; Kumar, D.; Goodman, J.C.; Groves, M.D.; Marchetti, D. The identification and characterization of breast cancer CTCs competent for brain metastasis. Sci. Transl. Med. 2013, 5, 180ra148. [Google Scholar] [CrossRef] [Green Version]
- Ameri, K.; Luong, R.; Zhang, H.; Powell, A.A.; Montgomery, K.D.; Espinosa, I.; Bouley, D.M.; Harris, A.L.; Jeffrey, S.S. Circulating tumour cells demonstrate an altered response to hypoxia and an aggressive phenotype. Br. J. Cancer 2010, 102, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Salnikov, A.V.; Liu, L.; Platen, M.; Gladkich, J.; Salnikova, O.; Ryschich, E.; Mattern, J.; Moldenhauer, G.; Werner, J.; Schemmer, P.; et al. Hypoxia induces EMT in low and highly aggressive pancreatic tumor cells but only cells with cancer stem cell characteristics acquire pronounced migratory potential. PLoS ONE 2012, 7, e46391. [Google Scholar] [CrossRef]
- Xie, K.; Huang, S. Regulation of cancer metastasis by stress pathways. Clin. Exp. Metastasis 2003, 20, 31–43. [Google Scholar] [CrossRef]
- Parmar, K.; Mauch, P.; Vergilio, J.A.; Sackstein, R.; Down, J.D. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc. Natl. Acad. Sci. USA 2007, 104, 5431–5436. [Google Scholar] [CrossRef] [Green Version]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Turcotte, R.; et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiozawa, Y.; Pedersen, E.A.; Havens, A.M.; Jung, Y.; Mishra, A.; Joseph, J.; Kim, J.K.; Patel, L.R.; Ying, C.; Ziegler, A.M.; et al. Human prostate cancer metastases target the hematopoietic stem cell niche to establish footholds in mouse bone marrow. J. Clin. Invest. 2011, 121, 1298–1312. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S.; O’Connell, J.T.; Gonzalez Herrera, K.N.; Wikman, H.; Pantel, K.; Haigis, M.C.; de Carvalho, F.M.; Damascena, A.; Domingos Chinen, L.T.; Rocha, R.M.; et al. PGC-1alpha mediates mitochondrial biogenesis and oxidative phosphorylation in cancer cells to promote metastasis. Nat. Cell Biol. 2014, 16, 992–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkowiak, K.; Kwiatkowski, M.; Buck, F.; Gorges, T.M.; Nilse, L.; Assmann, V.; Andreas, A.; Muller, V.; Wikman, H.; Riethdorf, S.; et al. Disseminated Tumor Cells Persist in the Bone Marrow of Breast Cancer Patients through Sustained Activation of the Unfolded Protein Response. Cancer Res. 2015, 75, 5367–5377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wouters, B.G.; Koritzinsky, M. Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat. Rev. Cancer 2008, 8, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. Glucose-regulated proteins in cancer: Molecular mechanisms and therapeutic potential. Nat. Rev. Cancer 2014, 14, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Koritzinsky, M.; Magagnin, M.G.; van den Beucken, T.; Seigneuric, R.; Savelkouls, K.; Dostie, J.; Pyronnet, S.; Kaufman, R.J.; Weppler, S.A.; Voncken, J.W.; et al. Gene expression during acute and prolonged hypoxia is regulated by distinct mechanisms of translational control. EMBO J. 2006, 25, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Baird, S.D.; Turcotte, M.; Korneluk, R.G.; Holcik, M. Searching for IRES. RNA 2006, 12, 1755–1785. [Google Scholar] [CrossRef] [Green Version]
- Kallergi, G.; Markomanolaki, H.; Giannoukaraki, V.; Papadaki, M.A.; Strati, A.; Lianidou, E.S.; Georgoulias, V.; Mavroudis, D.; Agelaki, S. Hypoxia-inducible factor-1alpha and vascular endothelial growth factor expression in circulating tumor cells of breast cancer patients. Breast Cancer Res. 2009, 11, R84. [Google Scholar] [CrossRef]
- Ward, J.P. Oxygen sensors in context. Biochim. Biophys. Acta 2008, 1777, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.T.; Lee, A.S. Measurement and modification of the expression level of the chaperone protein and signaling regulator GRP78/BiP in mammalian cells. Methods Enzymol. 2011, 490, 217–233. [Google Scholar] [PubMed]
- Eichelser, C.; Stuckrath, I.; Muller, V.; Milde-Langosch, K.; Wikman, H.; Pantel, K.; Schwarzenbach, H. Increased serum levels of circulating exosomal microRNA-373 in receptor-negative breast cancer patients. Oncotarget 2014, 5, 9650–9663. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pantel, K.; Dickmanns, A.; Zippelius, A.; Klein, C.; Shi, J.; Hoechtlen-Vollmar, W.; Schlimok, G.; Weckermann, D.; Oberneder, R.; Fanning, E.; et al. Establishment of micrometastatic carcinoma cell lines: A novel source of tumor cell vaccines. J. Natl. Cancer Inst. 1995, 87, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Bartkowiak, K.; Effenberger, K.E.; Harder, S.; Andreas, A.; Buck, F.; Peter-Katalinic, J.; Pantel, K.; Brandt, B.H. Discovery of a novel unfolded protein response phenotype of cancer stem/progenitor cells from the bone marrow of breast cancer patients. J. Proteome Res. 2010, 9, 3158–3168. [Google Scholar] [CrossRef]
- Meng, S.; Tripathy, D.; Frenkel, E.P.; Shete, S.; Naftalis, E.Z.; Huth, J.F.; Beitsch, P.D.; Leitch, M.; Hoover, S.; Euhus, D.; et al. Circulating tumor cells in patients with breast cancer dormancy. Clin Cancer Res 2004, 10, 8152–8162. [Google Scholar] [CrossRef] [Green Version]
- Helczynska, K.; Kronblad, A.; Jogi, A.; Nilsson, E.; Beckman, S.; Landberg, G.; Pahlman, S. Hypoxia promotes a dedifferentiated phenotype in ductal breast carcinoma in situ. Cancer Res. 2003, 63, 1441–1444. [Google Scholar]
- Reddy, R.K.; Mao, C.; Baumeister, P.; Austin, R.C.; Kaufman, R.J.; Lee, A.S. Endoplasmic reticulum chaperone protein GRP78 protects cells from apoptosis induced by topoisomerase inhibitors: Role of ATP binding site in suppression of caspase-7 activation. J. Biol. Chem. 2003, 278, 20915–20924. [Google Scholar] [CrossRef] [Green Version]
- Jaakkola, P.; Mole, D.R.; Tian, Y.M.; Wilson, M.I.; Gielbert, J.; Gaskell, S.J.; Kriegsheim, A.; Hebestreit, H.F.; Mukherji, M.; Schofield, C.J.; et al. Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001, 292, 468–472. [Google Scholar] [CrossRef]
- Connolly, E.; Braunstein, S.; Formenti, S.; Schneider, R.J. Hypoxia inhibits protein synthesis through a 4E-BP1 and elongation factor 2 kinase pathway controlled by mTOR and uncoupled in breast cancer cells. Mol. Cell Biol. 2006, 26, 3955–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citri, A.; Yarden, Y. EGF-ERBB signalling: Towards the systems level. Nat Rev Mol Cell Biol 2006, 7, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College Of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. J. Clin. Oncol. 2010, 28, 2784–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazel, M.; Jacot, W.; Pantel, K.; Bartkowiak, K.; Topart, D.; Cayrefourcq, L.; Rossille, D.; Maudelonde, T.; Fest, T.; Alix-Panabieres, C. Frequent expression of PD-L1 on circulating breast cancer cells. Mol. Oncol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Pantel, K. Tumor cell dissemination: Emerging biological insights from animal models and cancer patients. Cancer Cell 2013, 23, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Keller, L.; Pantel, K. Unravelling tumour heterogeneity by single-cell profiling of circulating tumour cells. Nat. Rev. Cancer 2019, 19, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Voss, M.J.; Moller, M.F.; Powe, D.G.; Niggemann, B.; Zanker, K.S.; Entschladen, F. Luminal and basal-like breast cancer cells show increased migration induced by hypoxia, mediated by an autocrine mechanism. BMC Cancer 2011, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Effenberger, K.E.; Borgen, E.; Eulenburg, C.Z.; Bartkowiak, K.; Grosser, A.; Synnestvedt, M.; Kaaresen, R.; Brandt, B.; Nesland, J.M.; Pantel, K.; et al. Detection and clinical relevance of early disseminated breast cancer cells depend on their cytokeratin expression pattern. Breast Cancer Res. Treat 2011, 125, 729–738. [Google Scholar] [CrossRef] [Green Version]
- DiGiovanna, M.P.; Stern, D.F.; Edgerton, S.M.; Whalen, S.G.; Moore, D., 2nd; Thor, A.D. Relationship of epidermal growth factor receptor expression to ErbB-2 signaling activity and prognosis in breast cancer patients. J. Clin. Oncol. 2005, 23, 1152–1160. [Google Scholar] [CrossRef]
- Riethdorf, S.; Muller, V.; Zhang, L.; Rau, T.; Loibl, S.; Komor, M.; Roller, M.; Huober, J.; Fehm, T.; Schrader, I.; et al. Detection and HER2 expression of circulating tumor cells: Prospective monitoring in breast cancer patients treated in the neoadjuvant GeparQuattro trial. Clin. Cancer Res. 2010, 16, 2634–2645. [Google Scholar] [CrossRef] [Green Version]
- Hofman, P.; Heeke, S.; Alix-Panabieres, C.; Pantel, K. Liquid biopsy in the era of immuno-oncology: Is it ready for prime-time use for cancer patients? Ann. Oncol. 2019, 30, 1448–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strati, A.; Koutsodontis, G.; Papaxoinis, G.; Angelidis, I.; Zavridou, M.; Economopoulou, P.; Kotsantis, I.; Avgeris, M.; Mazel, M.; Perisanidis, C.; et al. Prognostic significance of PD-L1 expression on circulating tumor cells in patients with head and neck squamous cell carcinoma. Ann. Oncol. 2017, 28, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Riethdorf, S.; Hildebrandt, L.; Heinzerling, L.; Heitzer, E.; Fischer, N.; Bergmann, S.; Mauermann, O.; Waldispuhl-Geigl, J.; Coith, C.; Schon, G.; et al. Detection and Characterization of Circulating Tumor Cells in Patients with Merkel Cell Carcinoma. Clin. Chem. 2019, 65, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Janning, M.; Kobus, F.; Babayan, A.; Wikman, H.; Velthaus, J.L.; Bergmann, S.; Schatz, S.; Falk, M.; Berger, L.A.; Bottcher, L.M.; et al. Determination of PD-L1 Expression in Circulating Tumor Cells of NSCLC Patients and Correlation with Response to PD-1/PD-L1 Inhibitors. Cancers 2019, 11, 835. [Google Scholar] [CrossRef] [Green Version]
- Kulasinghe, A.; Perry, C.; Kenny, L.; Warkiani, M.E.; Nelson, C.; Punyadeera, C. PD-L1 expressing circulating tumour cells in head and neck cancers. BMC Cancer 2017, 17, 333. [Google Scholar] [CrossRef] [Green Version]
- Carcereri de Prati, A.; Butturini, E.; Rigo, A.; Oppici, E.; Rossin, M.; Boriero, D.; Mariotto, S. Metastatic Breast Cancer Cells Enter Into Dormant State and Express Cancer Stem Cells Phenotype Under Chronic Hypoxia. J. Cell Biochem. 2017, 118, 3237–3248. [Google Scholar] [CrossRef]
- O’Dwyer, P.J.; Yao, K.S.; Ford, P.; Godwin, A.K.; Clayton, M. Effects of hypoxia on detoxicating enzyme activity and expression in HT29 colon adenocarcinoma cells. Cancer Res. 1994, 54, 3082–3087. [Google Scholar]
- Webb, T.E.; Hughes, A.; Smalley, D.S.; Spriggs, K.A. An internal ribosome entry site in the 5’ untranslated region of epidermal growth factor receptor allows hypoxic expression. Oncogenesis 2015, 4, e134. [Google Scholar] [CrossRef] [Green Version]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Product | Cell Line | ΔCq Under S (mRNA) | Condition | x-Fold Change (mRNA) | STDEV (mRNA) | x-Fold Change (Protein) | STDEV (Protein) | p-Value Protein Level vs. mRNA Level | p-Value Hyp vs. Reox (Protein Level) | p-Value Hyp vs. Reox (mRNA Level) |
|---|---|---|---|---|---|---|---|---|---|---|
| PD-L1 | MDA-MB-231 | 6.73 | Hyp | 0.56 | 0.06 | 1.98 | 0.44 | 0.029 | 0.004 | 0.045 |
| Reox | 1.23 | 0.09 | 1.57 | 0.40 | 0.277 | |||||
| MDA-MB-468 | 13.97 | Hyp | 0.66 | 0.27 | no signals | NA | NA | 0.347 | ||
| Reox | 0.47 | 0.08 | no signals | NA | ||||||
| MCF-7 | 16.52 | Hyp | 1.70 | 0.40 | no signals | NA | NA | 0.036 | ||
| Reox | 0.69 | 0.15 | no signals | NA | ||||||
| BC-M1 | 6.52 | Hyp | 1.07 | 0.06 | 3.65 | 0.69 | 0.022 | 0.024 | 0.001 | |
| Reox | 0.68 | 0.06 | 2.00 | 0.66 | 0.073 | |||||
| Vimentin | MDA-MB-231 | −2.78 | Hyp | 0.94 | 0.10 | 0.95 | 0.02 | 0.880 | 0.422 | 0.496 |
| Reox | 0.89 | 0.05 | 0.98 | 0.06 | 0.119 | |||||
| MDA-MB-468 | 7.11 | Hyp | 0.90 | 0.10 | 1.00 1 | 0 | NA | 0.023 | 0.309 | |
| Reox | 1.00 | 0.11 | 0.72 | 0.08 | NA | |||||
| MCF-7 | 8.26 | Hyp | 0.81 | 0.09 | no signals | NA | NA | 0.178 | ||
| Reox | 0.93 | 0.09 | no signals | NA | ||||||
| BC-M1 | −3.30 | Hyp | 1.08 | 0.24 | 1.02 | 0.08 | 0.715 | 0.237 | 0.309 | |
| Reox | 1.27 | 0.12 | 1.20 | 0.18 | 0.609 | |||||
| EGFR | MDA-MB-231 | 2.69 | Hyp | 0.87 | 0.15 | 1.90 | 0.41 | 0.037 | 0.028 | 0.006 |
| Reox | 1.50 | 0.13 | 1.11 | 0.35 | 0.185 | |||||
| MDA-MB-468 | 0.61 | Hyp | 1.17 | 0.14 | 3.05 | 0.79 | 0.050 | 0.887 | 0.572 | |
| Reox | 1.23 | 0.09 | 3.09 | 0.50 | 0.021 | |||||
| MCF-7 | 8.40 | Hyp | 1.10 | 0.16 | 21.58 | 16.50 | 0.165 | 0.227 | 0.496 | |
| Reox | 1.02 | 0.08 | 49.76 | 11.89 | 0.019 | |||||
| BC-M1 | 3.51 | Hyp | 1.17 | 0.10 | 4.99 | 0.38 | 0.002 | 0.133 | 0.002 | |
| Reox | 0.53 | 0.05 | 3.50 | 1.39 | 0.066 | |||||
| EpCAM | MDA-MB-231 | 4.33 | Hyp | 1.04 | 0.19 | no signals | NA | NA | 0.366 | |
| Reox | 0.91 | 0.09 | no signals | NA | ||||||
| MDA-MB-468 | −0.21 | Hyp | 0.71 | 0.08 | 0.43 | 0.05 | 0.011 | 0.178 | 0.010 | |
| Reox | 1.01 | 0.08 | 0.34 | 0.11 | 0.002 | |||||
| MCF-7 | 0.80 | Hyp | 0.94 | 0.10 | 1.54 | 0.28 | 0.052 | 0.089 | 0.081 | |
| Reox | 1.13 | 0.10 | 0.95 | 0.25 | 0.341 | |||||
| BC-M1 | 10.65 | Hyp | 0.88 | 0.08 | no signals | NA | NA | 0.022 | ||
| Reox | 1.38 | 0.17 | no signals | NA | ||||||
| ErbB2 | MDA-MB-231 | 8.91 | Hyp | 0.09 | 0.03 | 1.06 | 0.18 | 0.010 | 0.370 | 0.003 |
| Reox | 1.94 | 0.20 | 0.97 | 0.30 | 0.013 | |||||
| MDA-MB-468 | 13.42 | Hyp | 23.26 | 2.39 | 6.14 | 0.64 | 0.004 | 0.618 | 0.003 | |
| Reox | 1.71 | 0.50 | 6.94 | 2.66 | 0.072 | |||||
| MCF-7 | 7.19 | Hyp | 0.89 | 0.09 | 0.86 | 0.19 | 0.822 | 0.199 | 0.622 | |
| Reox | 0.96 | 0.20 | 0.97 | 0.29 | 0.963 | |||||
| BC-M1 | 6.44 | Hyp | 1.85 | 0.23 | 1.61 | 0.29 | 0.327 | 0.247 | 0.006 | |
| Reox | 0.18 | 0.03 | 1.29 | 0.10 | 0.001 | |||||
| ERα | MDA-MB-231 | 19.09 | Hyp | 1.71 | 0.45 | no signals | NA | NA | 0.052 | |
| Reox | 4.02 | 1.10 | no signals | NA | ||||||
| MDA-MB-468 | 14.55 | Hyp | 1.16 | 0.28 | no signals | NA | NA | 0.072 | ||
| Reox | 0.65 | 0.22 | no signals | NA | ||||||
| MCF-7 | 4.59 | Hyp | 0.91 | 0.07 | 0.23 | 0.01 | 0.003 | 0.072 | 0.027 | |
| Reox | 1.25 | 0.13 | 0.33 | 0.05 | 0.003 | |||||
| BC-M1 | 13.12 | Hyp | 1.39 | 0.15 | no signals | NA | NA | 0.003 | ||
| Reox | 0.52 | 0.08 | no signals | NA | ||||||
| Grp78 | MDA-MB-231 | 1.83 | Hyp | 0.43 | 0.04 | 0.86 | 0.04 | <0.001 | 0.672 | 0.003 |
| Reox | 1.14 | 0.10 | 0.83 | 0.15 | 0.003 | |||||
| MDA-MB-468 | 5.92 | Hyp | 2.90 | 0.27 | 0.49 | 0.14 | 0.001 | 0.085 | 0.003 | |
| Reox | 0.85 | 0.08 | 0.87 | 0.13 | 0.834 | |||||
| MCF-7 | 3.63 | Hyp | 1.11 | 0.10 | 0.36 | 0.11 | 0.001 | 0.903 | 0.227 | |
| Reox | 1.01 | 0.06 | 0.34 | 0.11 | 0.002 | |||||
| BC-M1 | 2.40 | Hyp | 1.12 | 0.11 | 1.14 | 0.15 | 0.862 | 0.295 | 0.003 | |
| Reox | 0.44 | 0.05 | 1.31 | 0.35 | 0.047 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartkowiak, K.; Koch, C.; Gärtner, S.; Andreas, A.; Gorges, T.M.; Pantel, K. In Vitro Modeling of Reoxygenation Effects on mRNA and Protein Levels in Hypoxic Tumor Cells upon Entry into the Bloodstream. Cells 2020, 9, 1316. https://doi.org/10.3390/cells9051316
Bartkowiak K, Koch C, Gärtner S, Andreas A, Gorges TM, Pantel K. In Vitro Modeling of Reoxygenation Effects on mRNA and Protein Levels in Hypoxic Tumor Cells upon Entry into the Bloodstream. Cells. 2020; 9(5):1316. https://doi.org/10.3390/cells9051316
Chicago/Turabian StyleBartkowiak, Kai, Claudia Koch, Sebastian Gärtner, Antje Andreas, Tobias M Gorges, and Klaus Pantel. 2020. "In Vitro Modeling of Reoxygenation Effects on mRNA and Protein Levels in Hypoxic Tumor Cells upon Entry into the Bloodstream" Cells 9, no. 5: 1316. https://doi.org/10.3390/cells9051316