Pregnancy and Fetal Development: Cephalic Presentation and Other Descriptive Ultrasonographic Findings from Clinically Healthy Bottlenose Dolphins (Tursiops truncatus) under Human Care

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Ultrasonography: Instrumentation and Methodology
2.3. Statistical Analyses
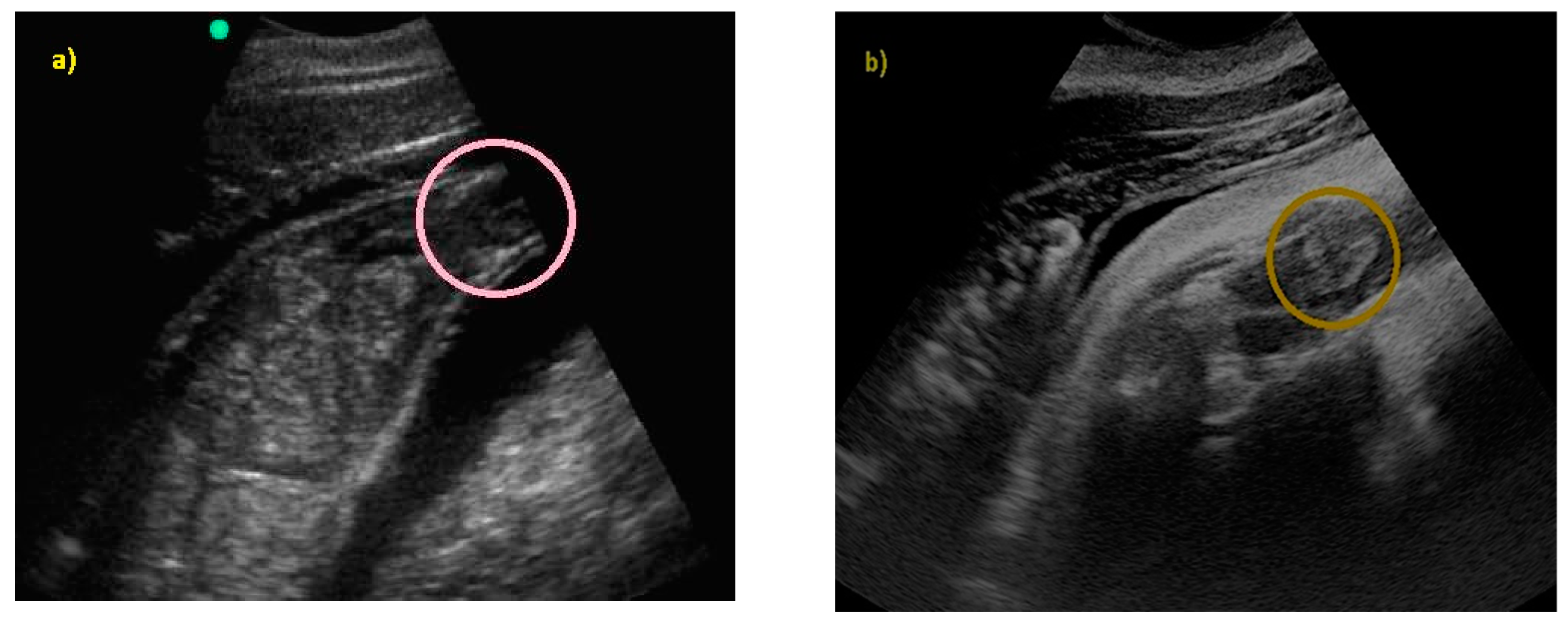
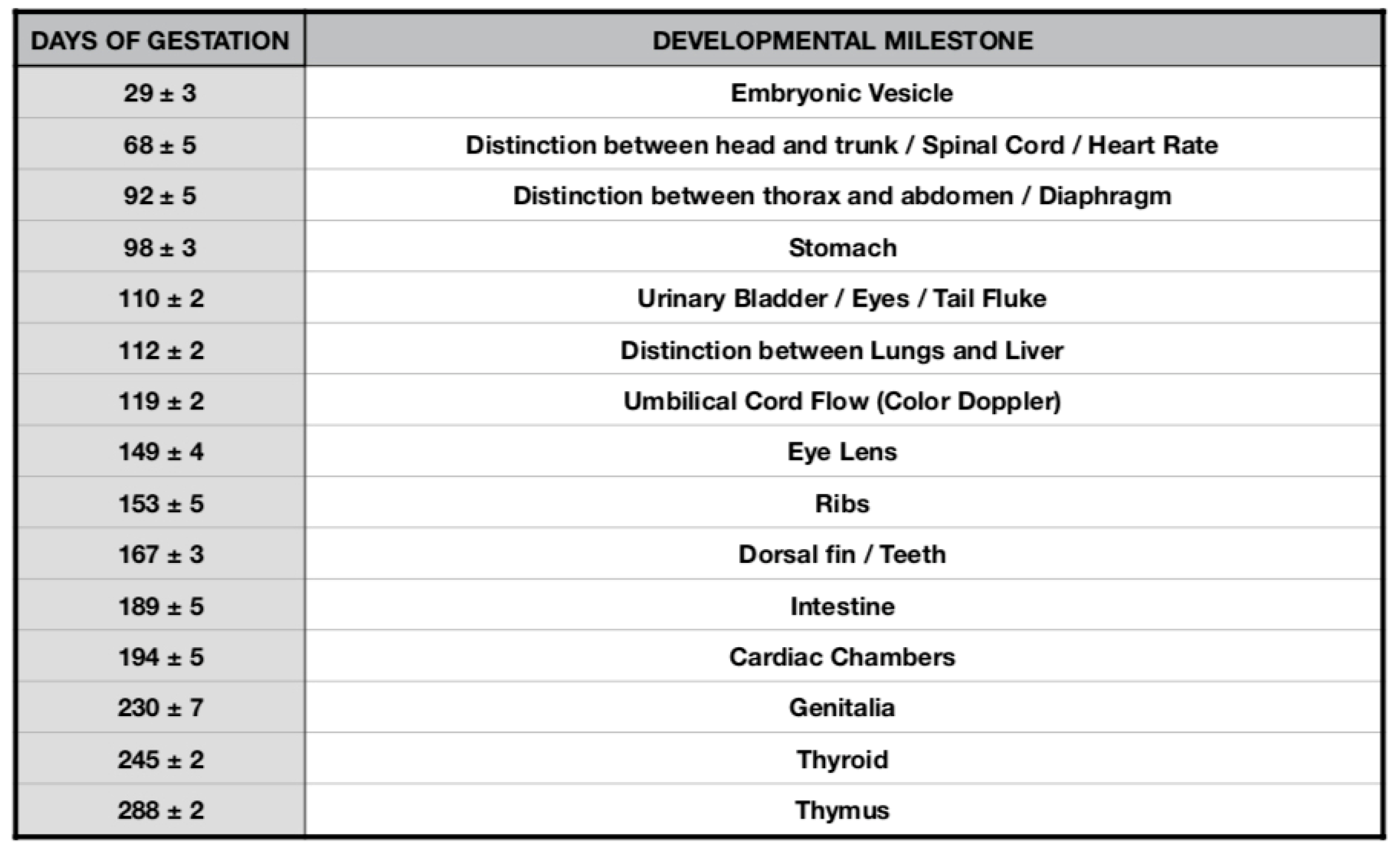
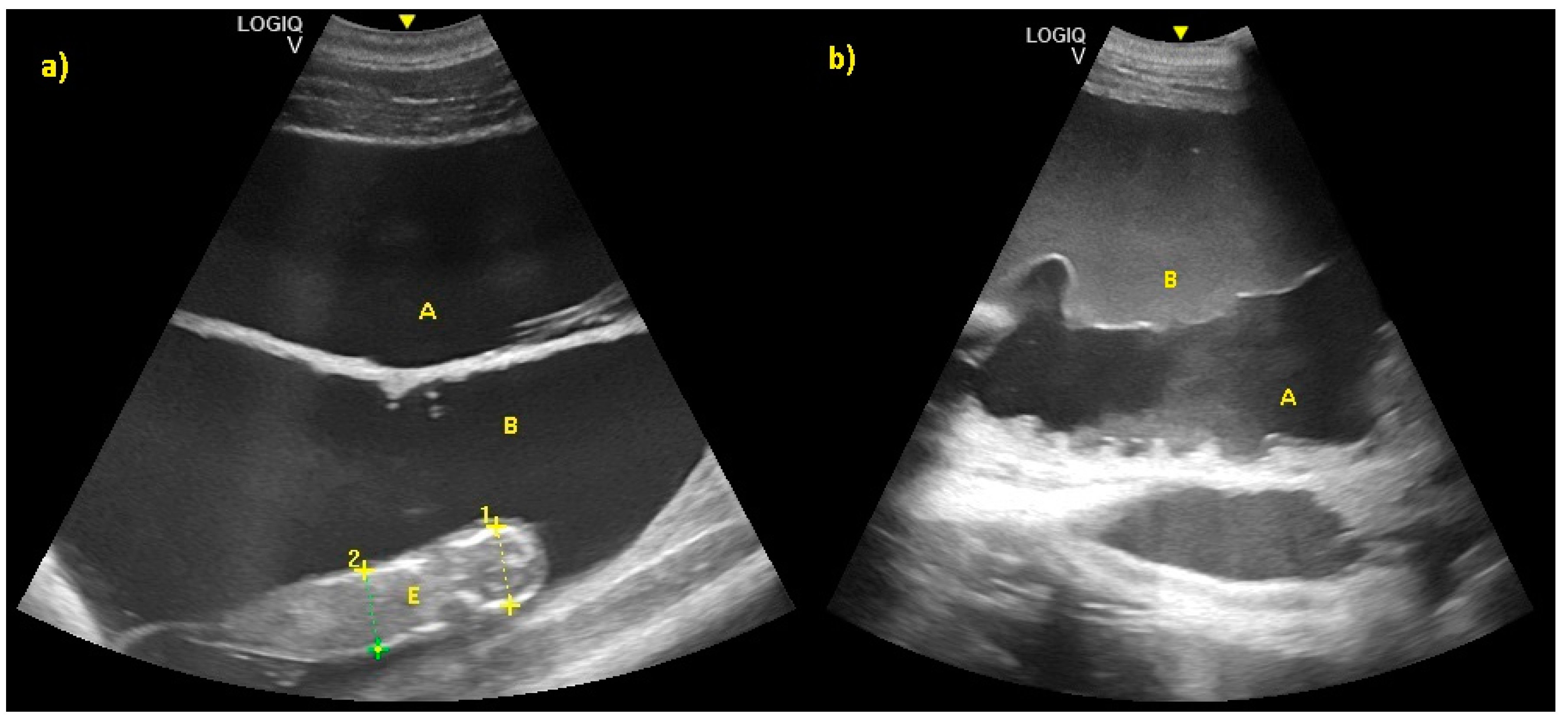
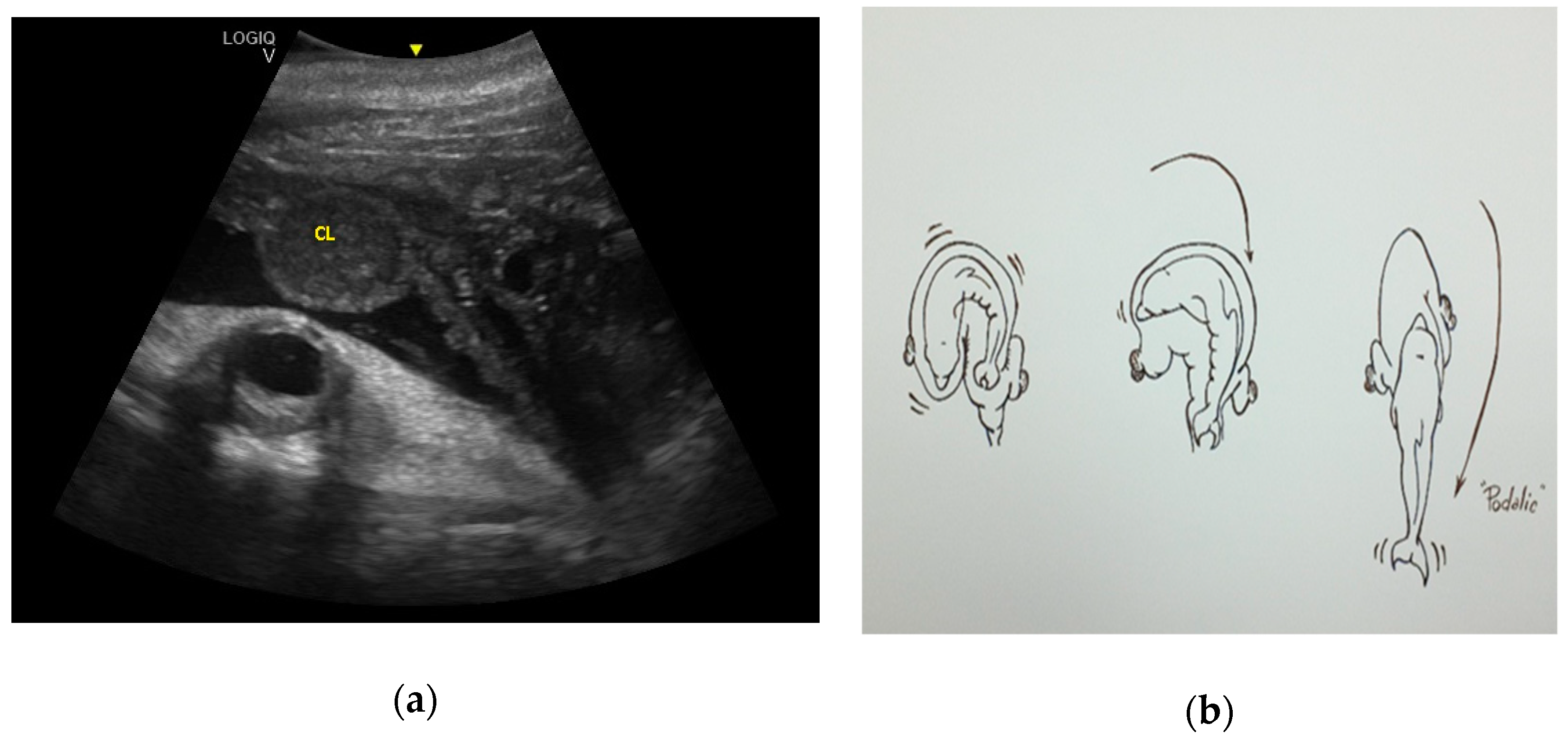
3. Results
4. Discussion
- (1)
- it must be conducted by an expert operator;
- (2)
- it depends on the features of the device;
- (3)
- animals must be trained for the voluntary medical behavior;
- (4)
- the animals must remain in water, which may not be safe for the instrumentation;
- (5)
- the external environment (and the light level, in particular) negatively affects results.
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Instrumentation and Methodology. Available online: https://www.amazon.nl/Handbook-Ultrasonography-Dolphins-Abdomen-English-ebook/dp/B00BRXUSPG (accessed on 24 May 2020).
- Fiorucci, L.; García-Párraga, D.; Macrelli, R.; Grande, F.; Flanagan, C.; Rueca, F.; Busechian, S.; Bianchi, B.; Arbelo, M.; Saviano, P. Determination of the main reference values in ultrasound examination of the gastrointestinal tract in clinically healthy bottlenose dolphins (Tursiops truncatus). Aquat. Mamm. 2015, 41, 284–294. [Google Scholar] [CrossRef]
- England, G.C.W. Ultrasound evaluation of pregnancy and spontaneous embryonic resorption in the bitch. J. Small Anim. Pract. 1992, 33, 430–436. [Google Scholar] [CrossRef]
- England, G.C.W. Ultrasonographic Assessment of Abnormal Pregnancy. J. Small Anim. Pract. 1998, 28, 849–868. [Google Scholar] [CrossRef]
- England, G.C.W.; Russo, M. Ultrasonographic characteristic of early pregnancy failure in bitches. Theriogenology 2006, 66, 1694–1698. [Google Scholar] [CrossRef] [PubMed]
- Aissi, A.; Alloui, N.; Slimani, C.; Touri, S. Preliminary study of the early ultrasonographic diagnosis of pregnancy and fetal development in dog. J. Anim. Vet. Adv. 2008, 7, 607–611. [Google Scholar]
- Nyland, T.G.; Mattoon, J.S. Physical principles, instrumentation and safety of Diagnostic Ultrasound. In Veterinary Diagnostic Ultrasound; WB Saunders: Philadelphia, PA, USA, 1995; pp. 3–18. [Google Scholar]
- Root, K.M.V. Pregnancy diagnosis and abnormalities of pregnancy in the dog. Theriogenology 2005, 64, 755–765. [Google Scholar] [CrossRef]
- Luvoni, C.G.; Beccaglia, M. The prediction of parturition date in canine pregnancy. Reprod. Dom. Anim. 2006, 41, 27–32. [Google Scholar] [CrossRef]
- Lamm, C.G.; Makoloski, C.L. Current avances in gestation and parturition in cats and dogs. Vet. Clin. N. Am. Small Anim. Pract. 2012, 42, 445–456. [Google Scholar] [CrossRef]
- Davidson, A.P.; Baker, T.W. Reproductive ultrasound of the bitch and queen. Top. Companion Anim. Med. 2009, 24, 55–63. [Google Scholar] [CrossRef]
- Adams, G.P.; Plotka, E.D.; Asa, C.S.; Ginther, O.J. Feasibility of characterizing reproductive events in large, non-domestic species by transrectal ultrasonic imaging. Zoo Biol. 1991, 10, 247–253. [Google Scholar] [CrossRef]
- Harrison, R.J.; Ridgway, S.H. Gonadal activity in some bottlenose dolphins (Tursiops truncatus). J. Zool. 1971, 165, 355–366. [Google Scholar] [CrossRef]
- Brook, F.M. Ultrasonographic imaging of the reproductive organs of the female bottlenose dolphin, Tursiops truncatus aduncus. Reproduction 2001, 121, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Neuenhoff, R.D.; Cowan, D.F.; Whitehead, H.; Marshall, C. Prenatal data impacts common bottlenose dolphin (Tursiops truncatus) growth parameters estimated by length-at-age curves. Mar. Mamm. Sci. 2011, 27, 195–216. [Google Scholar] [CrossRef]
- Kellar, N.M.; Speakman, T.R.; Smith, C.R.; Lane, S.M.; Balmer, B.C.; Trego, M.L.; Catelani, K.N.; Robbins, M.N.; Allen, C.D.; Wells, R.S.; et al. Low reproductive success rates of common bottlenose dolphins Tursiops truncatus in the northern Gulf of Mexico following the Deepwater Horizon disaster (2010–2015). Endanger. Species Res. 2017, 33, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Wells, R.S.; Smith, C.R.; Sweeney, J.C.; Townsend, F.I.; Fauquier, D.A.; Stone, R.; Langan, J.; Schwacke, L.H.; Rowles, T.K. Fetal survival of common bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Aquat. Mamm. 2014, 40, 252. [Google Scholar] [CrossRef]
- Robeck, T.R.; Curry, B.E.; McBain, J.F.; Kraemer, D.C. Reproductive biology of the bottlenose dolphin (Tursiops truncatus) and the potential application of advanced reproductive technologies. J. Zoo Wildl. Med. 1994, 25, 321–336. [Google Scholar]
- Robeck, T.R.; Atkinson, S.K.C.; Brook, F. Reproduction. In CRC Handbook of Marine Mammals Medicine, 2th ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC Press: Baca Raton, FL, USA, 2001; pp. 193–236. [Google Scholar]
- Robeck, T.R.; Steinman, K.J.; Yoshioka, M.; Jensen, E.; O’Brien, J.K.; Katsumata, E.; Gili, C.; McBain, J.F.; Sweeney, J.; Monfort, S.L. Estrous cycle characterisation and artificial insemination using frozen-thawed spermatozoa in the Bottlenose dolphin (Tursiops Truncatus). Reproduction 2005, 129, 659–674. [Google Scholar] [CrossRef] [Green Version]
- Ivančič, M.; Gomez, F.M.; Musser, W.B.; Barratclough, A.; Meegan, J.M.; Waitt, S.M.; Llerenas, A.C.; Jensen, E.C.; Smith, C.R. Ultrasonographic findings associated with normal pregnancy and fetal well-being in the bottlenose dolphin (Tursiops truncatus). Vet. Radiol. Ultrasound 2020, 2020, 1–12. [Google Scholar]
- Barratclough, A.; Gomez, F.M.; Morey, J.S.; Deming, A.; Parry, C.; Meegan, J.M.; Carlin, K.P.; Schwacke, L.; Venn-Watson, S.; Jensen, E.D.; et al. Pregnancy profiles in the common bottlenose dolphin (Tursiops truncatus): Clinical biochemical and hematological variations during healthy gestation and a successful outcome. Theriogenology 2020, 142, 92–103. [Google Scholar] [CrossRef]
- Williamson, P.; Gales, N.J.; Lister, S. Use of real-timeB-mode ultrasound for pregnancy diagnosis and measurement of fetal growth rate in captive bottlenose dolphins (Tursiops truncatus). J. Reprod. Fertil. 1990, 88, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Stone, L.R.; Johnson, R.L.; Sweeney, J.C.; Lewis, M.L. Fetal ultrasonography in dolphins with emphasis on gestational aging. In Zoo and Wild Animal Medicine: Current Therapy 4; Fowler, M.E., Miller., R.E., Eds.; WB Saunders: Philadelphia, PA, USA, 1999; pp. 501–506. [Google Scholar]
- Lacave, G.; Eggermont, M.; Verslycke, T.; Kinoshita, R. Prediction from ultrasonographic measurements of the expected delivery date in two species of bottlenose. Vet. Rec. 2004, 154, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sklansky, M. Fetal cardiovascular malformations and arrhythmias. In Creasy and Resnik’s Maternal-Fetal Medicine, 6th ed.; Creasy, R.K., Resnik, R., Iams, J.D., Lockwood, C.J., Moore, T.R., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2009; pp. 305–345. [Google Scholar]
- Sklansky, M.; Renner, M.; Clough, P.; Levine, G.; Campbell, M.; Stone, R.; Schmitt, T.; Chang, R.K.; Shannon-Rodriguez, J. Fetal Echocardiographic Evaluation of the Bottlenose Dolphin (Tursiops truncatus). J. Zoo Wildl. Med. 2010, 41, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Rychik, J.; Ayres, N.; Cuneo, B.; Gotteiner, N.; Hornberger, L.; Spevak, P.; Van der Veld, M. American Society of Echocardiography guidelines and standards for performance of the fetal echocardiogram. J. Am. Soc. Echocardiogr. 2004, 17, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Makikallio, K.; Räsänen, J.; Mäkikallio, T.; Vuolteenaho, O.; Huhta, J.C. Human fetal cardiovascular profile score and neonatal outcome in intrauterine growth restriction Ultrasound. Obstet. Gynecol. 2008, 31, 48–54. [Google Scholar]
- Powell, J.; Archibald, R.; Cross, C.; Rotstein, D.; Soop, V.; McFee, W. Multiple congenital cardiac abnormalities in an Atlantic bottlenose dolphin (Tursiops truncatus). J. Wildl. Dis. 2009, 45, 839–842. [Google Scholar] [CrossRef] [Green Version]
- García-Párraga, D.; Brook, F.; Crespo-Picazo, J.L.; Valls, M.; Penadés, M.; Ortega, J.; Corpa, J.M. Recurrent umbilical cord accidents in a bottlenose dolphin, Tursiops truncatus. Dis. Aquat. Org. 2014, 108, 177–180. [Google Scholar] [CrossRef]
- Smith, C.R.; Wong, S.K.; Jensen, E.D.; Venn-Watson, S.K. Fetal omphalocele in a common bottlenose dolphin (Tursiops truncatus). J. Zoo Wildl. Med. 2013, 44, 87–92. [Google Scholar] [CrossRef]
- Tanaka, M.; Izawa, T.; Kuwamura, M.; Ozaki, M.; Nakao, T.; Ito, S.; Yamate, J. A case of meconium aspiration syndrome in a bottlenose dolphin (Tursiops truncatus) calf. J. Vet. Med. Sci. 2014, 76, 81–84. [Google Scholar] [CrossRef] [Green Version]
- Brook, F.M. Ultrasound Diagnosis of Anencephaly in the Fetus of a Bottlenose Dolphin (Tursiops aduncas). J. Zoo Wildl. Med. 1994, 25, 569–574. [Google Scholar]
- Thewisen, J.G.M.; Heyning, J.E. Embryogenesis and development in Stenella atenuatta and other cetaceans. In Reproductive Biology and Phylogeny in Cetacea, Whales, Dolphins and Porpoises; Miller, D.L., Ed.; Science Publishers: Enflield, NH, USA, 2007; pp. 307–330. [Google Scholar]
- Reidenberg, J.S.; Laitman, J.T. Prenatal development in cetaceans. In Encyclopedia of Marine Mammals, 2nd ed.; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 220–230. [Google Scholar]
- Cozzi, B.; Huggenberger, S.; Oelschläger, H. Anatomy of Dolphins: Insights into Body Structure and Function; Academic Press: London, UK, 2017; pp. 1–438. [Google Scholar]
- Sterba, O.; Klima, M.; Schildger, B. Embryology of dolphins. Staging and ageing of embryos and fetuses of some cetaceans. Adv. Anat. Embryol. Cell Biol. 2000, 157, 1–133. [Google Scholar]
- Zarzosa, G.R.; Fernández, A.L.; Gomariz, F.M.; Cano, F.G.; Laguía, M.S.; Espinosa, A.A.; de los Ríos y Loshuertos, A.G. A Study of the Head during Prenatal and Perinatal Development of Two Fetuses and One Newborn Striped Dolphin (Stenella coeruleoalba) Using Dissections, Sectional Anatomy, CT, and MRI: Anatomical and Functional Implications in Cetaceans and Terrestrial Mammals. Animals 2019, 9, 1139. [Google Scholar]
- Michel, E.; Spörri, M.; Ohlerth, S.; Reichler, L. Prediction of parturition date in the bitch and queen. Reprod. Domest. Anim. 2011, 46, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Sedmera, D.; Misek, I.; Milan, K.; Thompson, R.P. Heart Development in the Spotted Dolphin (Stenella attenuata). Anat. Rec. 2003, 273A, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Adams-Brendemuehl, C.; Pipers, F.S. Antepartum evaluations of the equine fetus. J. Reprod. Fertil. 1987, 35, 565–573. [Google Scholar]
- Capellini, I.; Venditti, C.; Barton, R.A. Placentation and Maternal Investment in Mammals. Am. Nat. 2011, 177, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Berta, A.; Sumich, J.L.; Kovacs, K.M. Cetacean evolution and systematics. In Marine Mammal Evolutionary Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 2005; pp. 165–209. [Google Scholar]
- Smith, C.R.; Wong, S.K.; Jensen, E.D.; Ruiz, C.; Nollens, H. Overview of neonatal management techniques at the U.S. Navy Marine Mammal Program. Int. Assoc. Aquat. Anim. Med. Proc. Nassau. Bahamas 2006, 37, 164–167. [Google Scholar]
- Slijper, E.J. Functional morphology of the reproductive system in Cetacea. In Whales, Dolphins and Porpoises; Norris, K.S., Berkeley, L.A., Eds.; University of California Press: Berkeley, CA, USA, 1966; pp. 277–319. [Google Scholar]
- Deng, X.; Hao, Y.; Serres, A.; Wang, K.; Wang, D. Position at Birth and Possible Effects on Calf Survival in Finless Porpoises (Neophocaena asiaeorientalis). Aquat. Mamm. 2019, 45, 4. [Google Scholar] [CrossRef]
- Gol’din, P.E. Case of cephalic presentation of fetus in a harbor porpoise (Phocoena phocoena), with notes on other aquatic mammals. Vestn. zool. 2011, 45, 33–38. [Google Scholar]
- Essapian, F.K. Observations on Abnormalities of Parturition in Captive Bottle-Nosed Dolphins, Tursiops truncatus, and Concurrent Behavior of Other Porpoises. J. Mammal. 1963, 44, 405–414. [Google Scholar] [CrossRef]
- Hart, P.; Van der Kemp, J.S. Cephalic presentation observed in a white-beaked dolphin, Lagenorhynchus albirostris. Lutra 1999, 41, 21–24. [Google Scholar]
- Vladykov, V.D. Études sur les mammifères aquatiques. III. Chasse, biologie et valeur économique du marsouin blanc ou béluga (Delphinapterus leucas) du fleuve et du golfe du Saint-Laurent; Département des pêcheries de la province de Québec: Québec, QC, Canada, 1944; p. 194.
- Doan, K.H.; Douglas, C.W. Beluga of the Churchill region of Hudson Bay. Bull. Fish. Res. Board Can. 1953, 98, 1–27. [Google Scholar]
- Kleinenberg, S.E.; Yablokov, A.V.; Bel’kovich, V.M.; Tarasevich, M.N. Beluga: The Experience of Monographic Species Study; Nauka: Moscow, Russia, 1964; p. 456. [Google Scholar]
- Baker, J.R.; Martin, A.R. Causes of mortality and parasites and incidental lesions in harbour porpoises (Phocoena phocoena) from British waters. Vet. Rec. 1992, 130, 554–558. [Google Scholar] [CrossRef] [PubMed]













© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saviano, P.; Fiorucci, L.; Grande, F.; Macrelli, R.; Troisi, A.; Polisca, A.; Orlandi, R. Pregnancy and Fetal Development: Cephalic Presentation and Other Descriptive Ultrasonographic Findings from Clinically Healthy Bottlenose Dolphins (Tursiops truncatus) under Human Care. Animals 2020, 10, 908. https://doi.org/10.3390/ani10050908
Saviano P, Fiorucci L, Grande F, Macrelli R, Troisi A, Polisca A, Orlandi R. Pregnancy and Fetal Development: Cephalic Presentation and Other Descriptive Ultrasonographic Findings from Clinically Healthy Bottlenose Dolphins (Tursiops truncatus) under Human Care. Animals. 2020; 10(5):908. https://doi.org/10.3390/ani10050908
Chicago/Turabian StyleSaviano, Pietro, Letizia Fiorucci, Francesco Grande, Roberto Macrelli, Alessandro Troisi, Angela Polisca, and Riccardo Orlandi. 2020. "Pregnancy and Fetal Development: Cephalic Presentation and Other Descriptive Ultrasonographic Findings from Clinically Healthy Bottlenose Dolphins (Tursiops truncatus) under Human Care" Animals 10, no. 5: 908. https://doi.org/10.3390/ani10050908