Hemp (Cannabis sativa L.) Protein Hydrolysates Promote Anti-Inflammatory Response in Primary Human Monocytes

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Compounds and Sampling
2.2. Hemp Protein Products Preparation
2.3. Blood Collection and Monocyte Isolation
2.4. Treatments of Cells in Culture
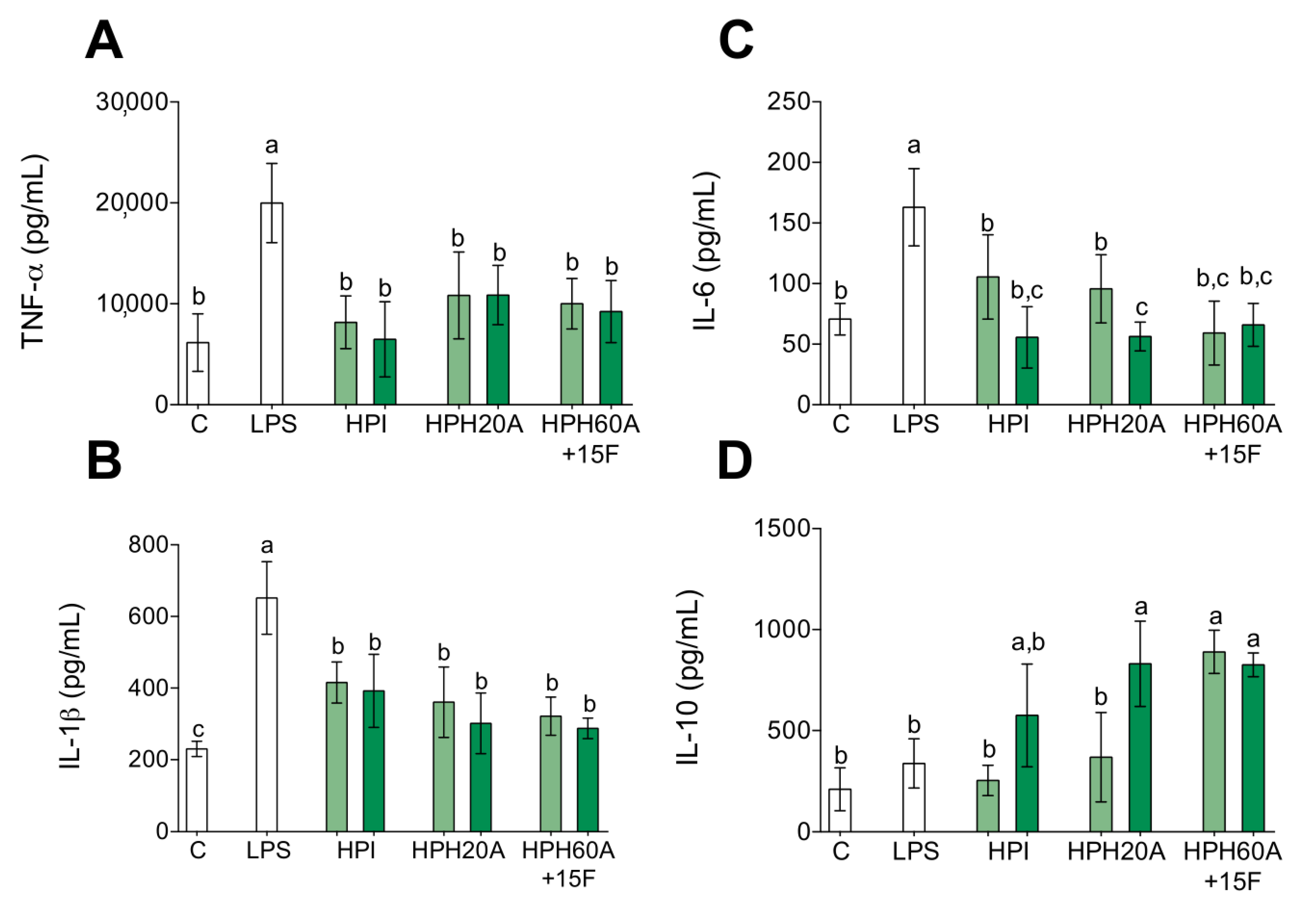
2.5. Measurement of Cytokine Release
2.6. RNA Isolation and RT-qPCR Analysis
2.7. Statistical Method
3. Results
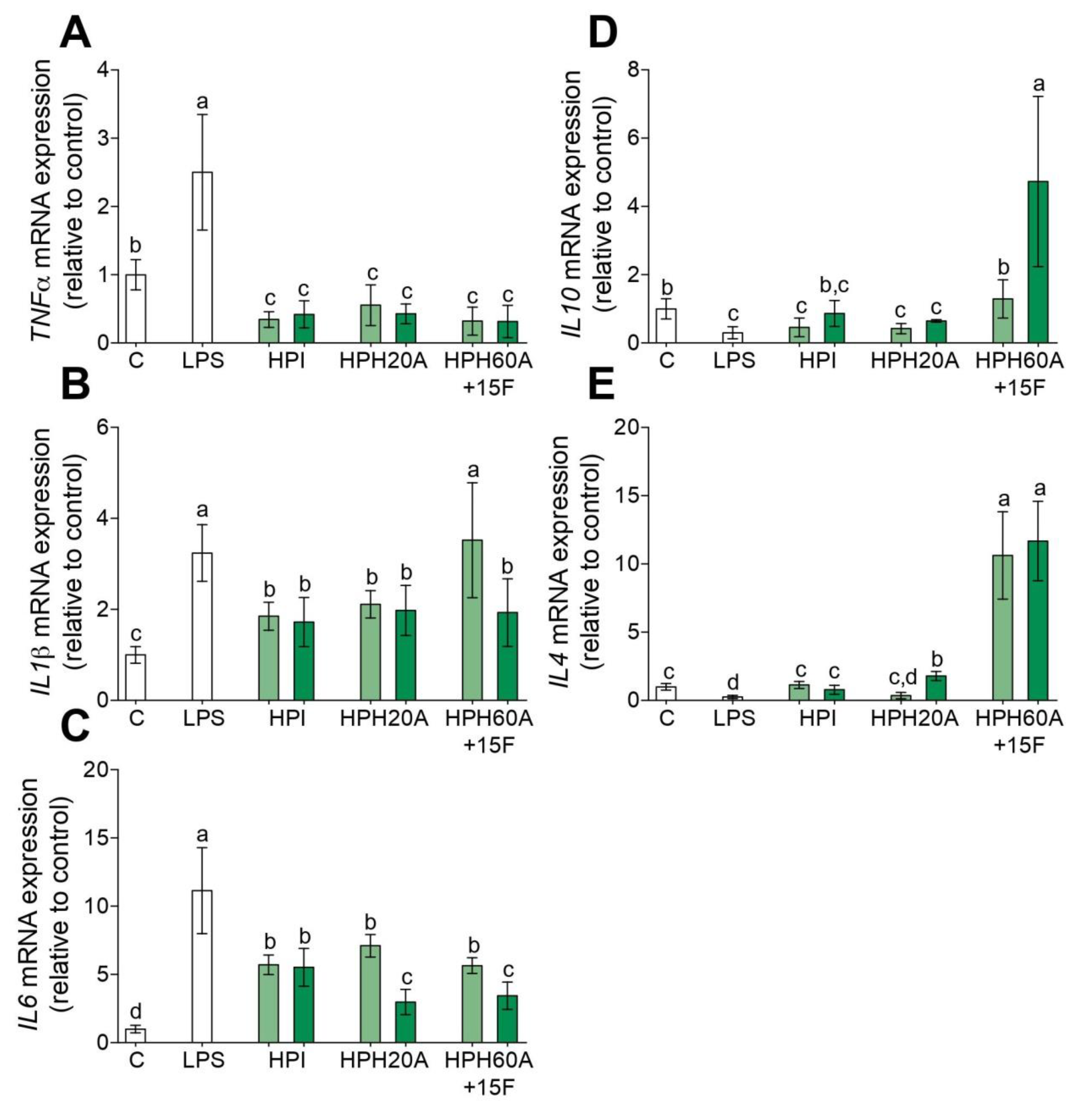
3.1. Anti-Inflammatory Properties of Hemp Protein Hydrolysates
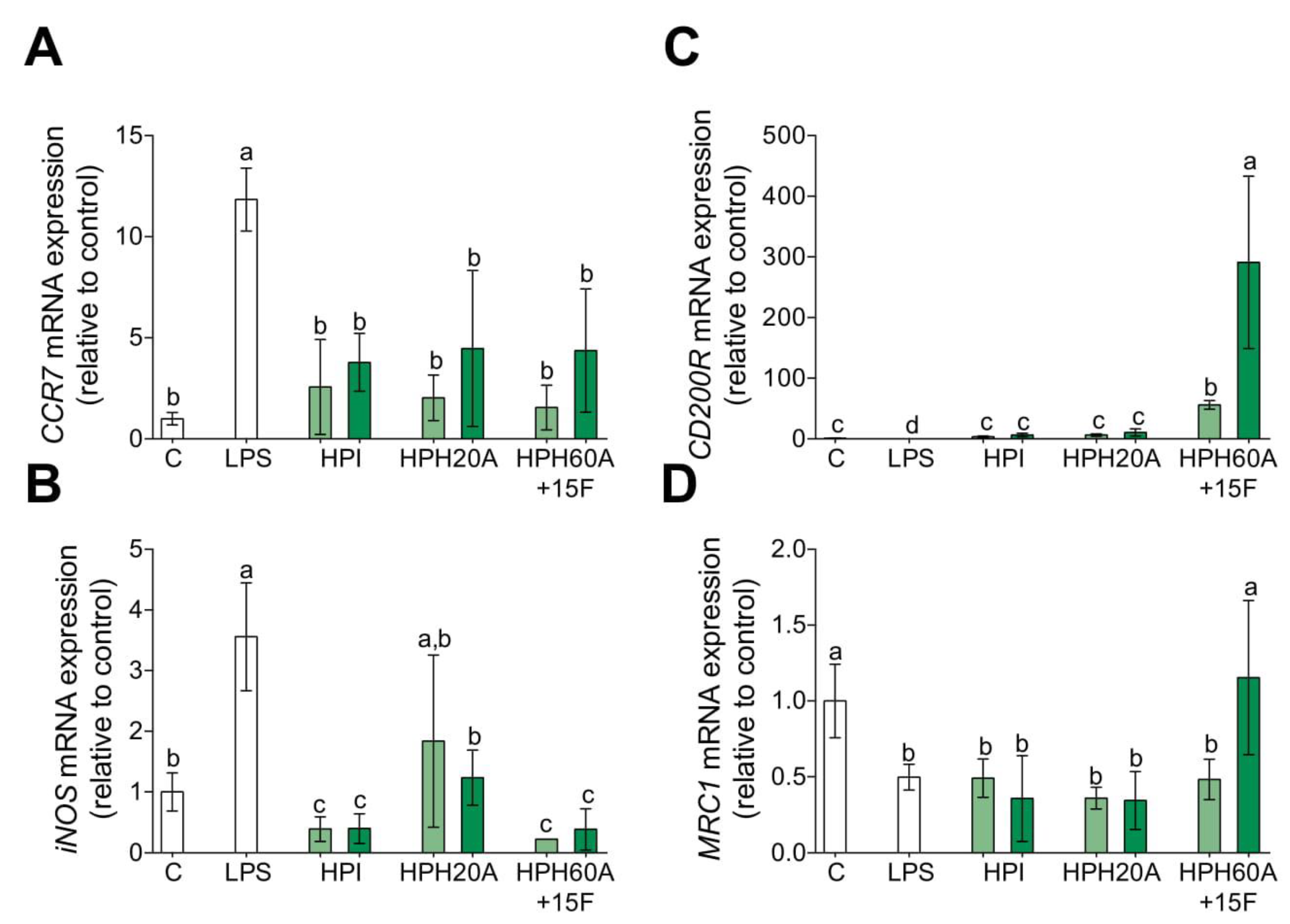
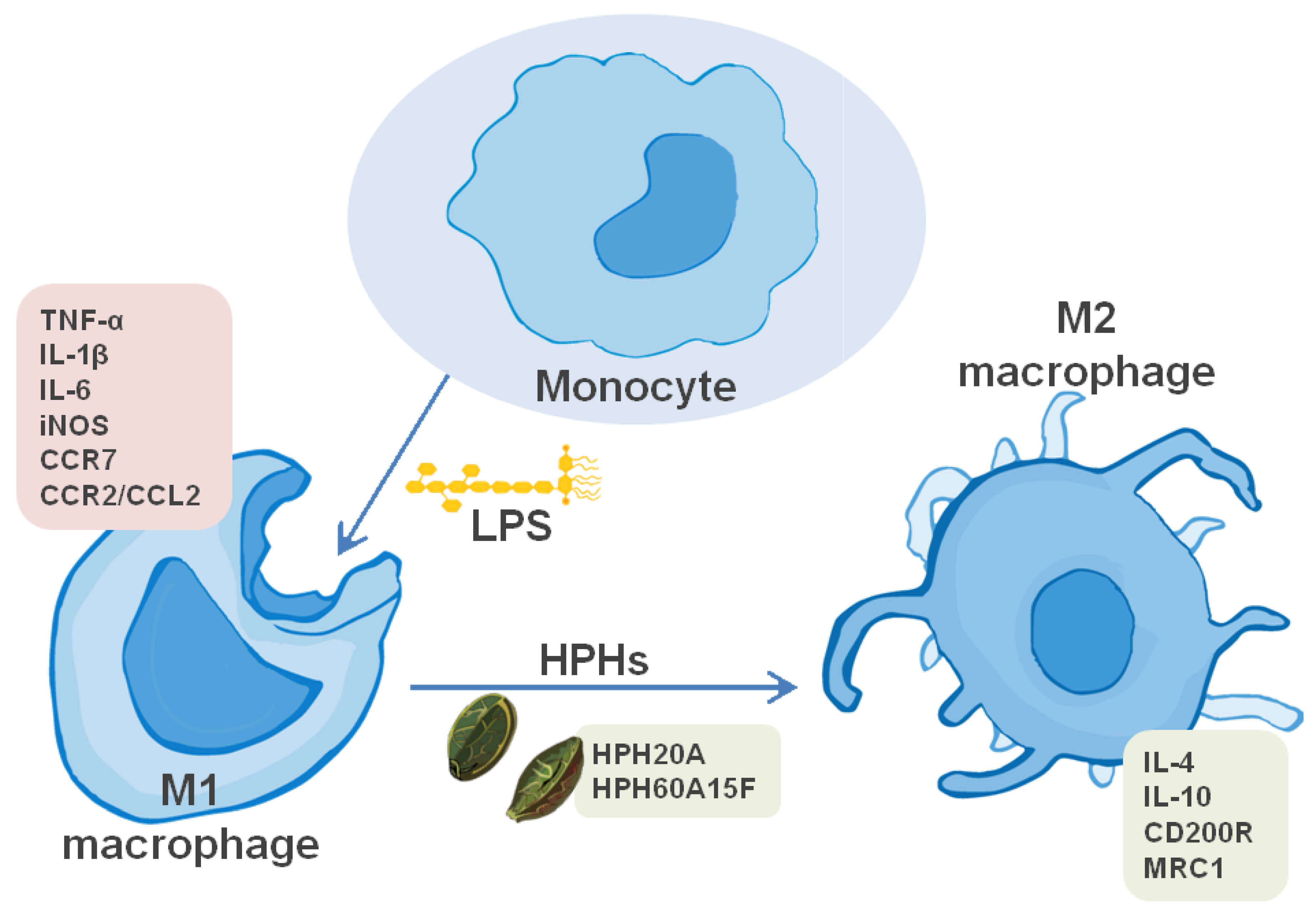
3.2. Hemp Protein Hydrolysates Shifts M1 toward M2 Macrophage Phenotype
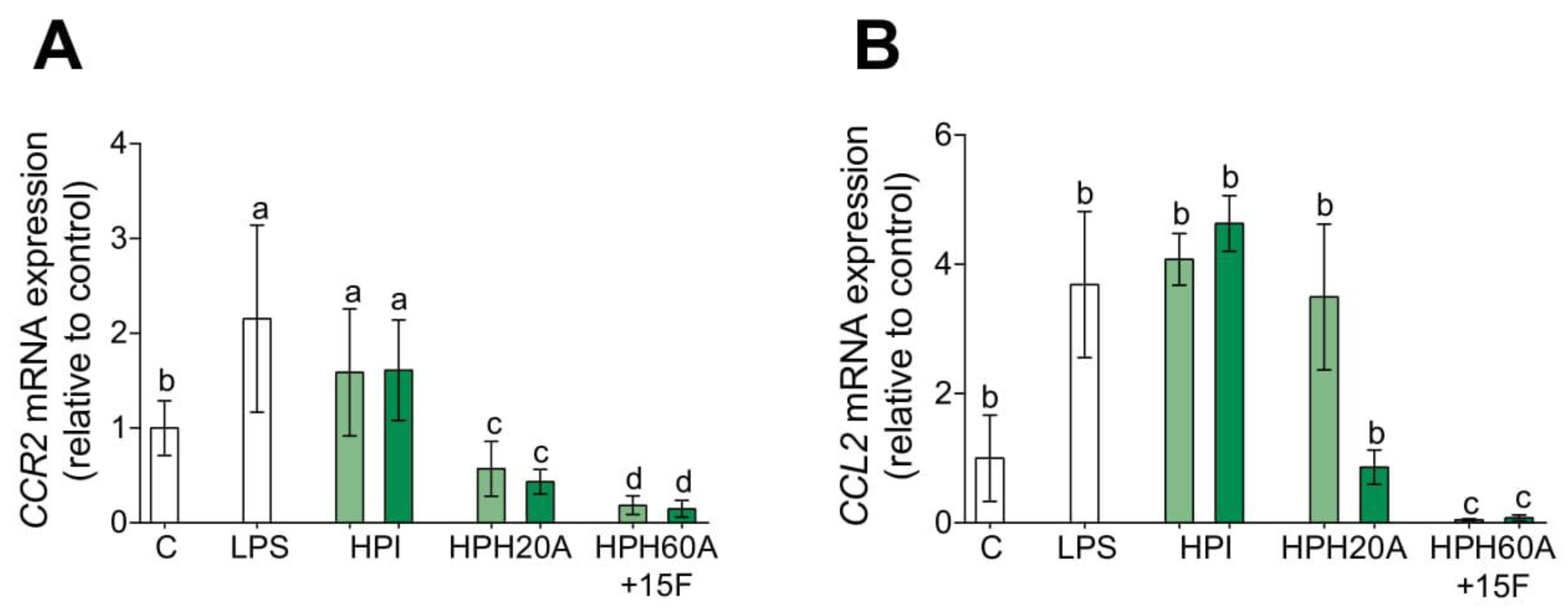
3.3. Hemp Protein Hydrolysates Modulates CCR2-Dependent Migration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient (g/100 g Product) | Hemp Seed | Hemp Protein Isolate (HPI) | Hemp Protein Hydrolysate: HPH20A | Hemp Protein Hydrolysate: HPH 60A + 15AF |
|---|---|---|---|---|
| Proteins | 24.18 ± 1.01 | 96.46 ± 0.93 | 86.56 ± 0.78 | 83.02 ± 0.29 |
| Carbs | 3.81 ± 0.76 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Fat | 32.35 ± 0.57 | --------- | --------- | --------- |
| Humidity | 4.49 ± 0.01 | 2.19 ± 0.01 | 7.34 ± 0.03 | 8.85 ± 0.04 |
| Ash | 5.09 ± 0.07 | 1.06 ± 0.10 | 6.10 ± 0.75 | 8.13 ± 0.26 |
| Fiber | 29.72 ± 1.44 | 00.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Polifenols | 0.35 ± 0.02 | 0.04 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Amino Acids (g/100 g Protein) | Defatted Hemp Flour (DHF) | Hemp Protein Isolate (HPI) | Hemp Protein Hydrolysate: HPH20A | Hemp Protein Hydrolysate: HPH 60A + 15AF |
|---|---|---|---|---|
| Aspartic acid + asparragine | 11.34 ± 0.37 | 12.35 ± 0.34 | 12.40 ± 0.29 | 12.08 ± 0.51 |
| Glutamic acid + Glutamine | 18.41 ± 0.13 | 18.86 ± 0.35 | 18.83 ± 0.25 | 18.58 ± 0.12 |
| Alanine | 4.89 ± 0.10 | 5.01 ± 0.14 | 4.99 ± 0.10 | 5.00 ± 0.11 |
| Arginine | 12.68 ± 0.21 | 13.44 ± 0.03 | 13.34 ± 0.24 | 13.78 ± 0.23 |
| Cysteine | 1.22 ± 0.02 | 1.98 ± 0.08 | 1.81 ± 0.18 | 1.65 ± 0.21 |
| Glycine | 4.74 ± 0.07 | 4.79 ± 0.23 | 4.64 ± 0.11 | 4.64 ± 0.12 |
| Histidine | 3.07 ± 0.01 | 2.85 ± 0.05 | 2.80 ± 0.02 | 2.79 ± 0.03 |
| Isoleucine | 4.23 ± 0.08 | 3.02 ± 0.40 | 3.57 ± 0.15 | 3.85 ± 0.20 |
| Leucine | 7.12 ± 0.05 | 7.13 ± 0.11 | 7.12 ± 0.13 | 7.34 ± 0.07 |
| Lysine | 3.78 ± 0.09 | 3.04 ± 0.08 | 3.10 ± 0.06 | 3.23 ± 0.04 |
| Methionine | 2.14 ± 0.02 | 0.89 ± 0.34 | 0.60 ± 0.03 | 0.64 ± 0.04 |
| Phenylalanine | 4.76 ± 0.09 | 5.12 ± 0.02 | 5.04 ± 0.05 | 5.19 ± 0.08 |
| Proline | 2.66 ± 0.69 | 3.02 ± 0.00 | 2.83 ± 0.05 | 1.29 ± 0.19 |
| Serine | 5.57 ± 0.11 | 6.31 ± 0.21 | 6.36 ± 0.19 | 6.31 ± 0.22 |
| Threonine | 3.79 ± 0.08 | 3.79 ± 0.10 | 3.89 ± 0.06 | 4.02 ± 0.07 |
| Tryptophan | 0.76 ± 0.05 | 0.55 ± 0.02 | 0.66 ± 0.01 | 0.82 ± 0.03 |
| Tyrosine | 3.57 ± 0.06 | 4.08 ± 0.06 | 3.98 ± 0.03 | 4.06 ± 0.12 |
| Valine | 5.27 ± 0.11 | 3.77 ± 0.40 | 4.05 ± 0.42 | 4.71 ± 0.19 |
References
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Schluttenhofer, C.; Yuan, L. Challenges towards revitalizing hemp: A multifaceted crop. Trends Plant Sci. 2017, 22, 917–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamone, G.; Picariello, G.; Ramondo, A.; Nicolai, M.A.; Ferranti, P. Production, digestibility and allergenicity of hemp (Cannabis sativa L.) protein isolates. Food Res. Int. 2019, 115, 562–571. [Google Scholar] [CrossRef]
- Carus, M. The European Hemp Industry: Cultivation, processing and applications for fibres, shivs and seeds. EIHA 2017. Available online: www.eiha.org (accessed on 16 April 2020).
- Industrial Hemp Market Size, Share & Trends Analysis Report By Product (Seeds, Fiber, Shives), By Application (Animal Care, Textiles, Food & Beverages, Personal Care), and Segment Forecasts, 2020–2027. Grand View Res. 2019. Available online: https://www.grandviewresearch.com (accessed on 16 April 2020).
- Wang, S.; Luo, Q.; Fan, P. Cannabisin F from hemp (Cannabis sativa) seed suppresses lipopolysaccharide-induced inflammatory responses in BV2 Microglia as SIRT1 Modulator. Int. J. Mol. Sci. 2019, 20, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Yan, X.; Bobrovskaya, L.; Ji, M.; Yuan, H.; Lou, H.; Fan, P. Anti-neuroinflammatory effects of grossamide from hemp seed via suppression of TLR-4-mediated NF-κB signaling pathways in lipopolysaccharide-stimulated BV2 microglia cells. Mol. Cell. Biochem. 2017, 428, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. Reverse-phase HPLC separation of hemp seed (Cannabis sativa L.) protein hydrolysate produced peptide fractions with enhanced antioxidant capacity. Plant Foods Hum. Nutr. 2013, 68, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef]
- Aluko, R.E. Hemp Seed (Cannabis sativa L.) Proteins. In Sustainable Protein Sources; Elsevier: Winnipeg, MB, Canada, 2017; pp. 121–132. [Google Scholar]
- Tang, C.H.; Wang, X.S.; Yang, X.Q. Enzymatic hydrolysis of hemp (Cannabis sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem. 2009, 114, 1484–1490. [Google Scholar] [CrossRef]
- Zanoni, C.; Aiello, G.; Arnoldi, A.; Lammi, C. Hempseed peptides exert hypocholesterolemic effects with a statin-like mechanism. J. Agric. Food Chem. 2017, 65, 8829–8838. [Google Scholar] [CrossRef]
- Malomo, S.A.; Onuh, J.O.; Girgih, A.T.; Aluko, R.E. Structural and antihypertensive properties of enzymatic hemp seed protein hydrolysates. Nutrients 2015, 7, 7616–7632. [Google Scholar] [CrossRef] [Green Version]
- Teh, S.S.; Bekhit, A.E.A.; Carne, A.; Birch, J. Antioxidant and ACE-inhibitory activities of hemp (Cannabis sativa L.) protein hydrolysates produced by the proteases AFP, HT, Pro-G, actinidin and zingibain. Food Chem. 2016, 203, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Mclaughlin, T.; Ackerman, S.E.; Shen, L.; Engleman, E. Role of innate and adaptive immunity in obesity-associated metabolic disease. J. Clin. Investig. 2017, 127, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Inagawa, H. Gene expression in lipopolysaccharide-treated human monocytes following interaction with hepatic cancer cells. Anticancer Res. 2016, 36, 3688–3704. [Google Scholar]
- Kratofil, R.M.; Kubes, P.; Deniset, J.F. Monocyte Conversion during inflammation and injury. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaler, B.; Hohensinner, P.J.; Krychtiuk, K.A.; Matzneller, P.; Koller, L.; Brekalo, M.; Maurer, G.; Huber, K.; Zeitlinger, M.; Jilma, B.; et al. Differential in vivo activation of monocyte subsets during low-grade inflammation through experimental endotoxemia in humans. Sci. Rep. 2016, 6, 30162. [Google Scholar] [CrossRef]
- Awad, F.; Assrawi, E.; Jumeau, C.; Georgin-Lavialle, S.; Cobret, L.; Duquesnoy, P.; Piterboth, W.; Thomas, L.; Stankovic-Stojanovic, K.; Louvrier, C.; et al. Impact of human monocyte and macrophage polarization on NLR expression and NLRP3 inflammasome activation. PLoS ONE 2017, 12, e0175336. [Google Scholar] [CrossRef]
- Röszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.L.; Tai, J.J.; Wong, W.C.; Han, H.; Sem, X.; Yeap, W.H.; Kourilsky, P.; Wong, S.C. Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2011, 12, 1035–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. World Medical Association Declaration of Helsinki Ethical Principles for Medical Research Involving Human Subjects Adopted. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan-Linares, M.C.; Bermudez, B.; Martin, M.E.; Muñoz, E.; Abia, R.; Millan, F.; Muriana, F.J.G.; Montserrat-De La Paz, S. Unsaponifiable fraction isolated from grape (Vitis vinifera L.) seed oil attenuates oxidative and inflammatory responses in human primary monocytes. Food Funct. 2018, 9, 2517–2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation pathways: A general review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Schain, M.; Kreisl, W.C. Neuroinflammation in Neurodegenerative Disorders—A Review. Curr. Neurol. Neurosci. Rep. 2017, 17, 25. [Google Scholar] [CrossRef]
- Rogero, M.M.; Calder, P.C. Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Narumiya, S. Prostaglandin-cytokine crosstalk in chronic inflammation. Br. J. Pharmacol. 2019, 176, 337–354. [Google Scholar] [CrossRef]
- Phillips, C.M.; Shivappa, N.; Hebert, J.R.; Perry, I.J. Dietary inflammatory index and biomarkers of lipoprotein metabolism, inflammation and glucose homeostasis in adults. Nutrients 2018, 10, 1033. [Google Scholar] [CrossRef] [Green Version]
- Mazidi, M.; Shivappa, N.; Wirth, M.D.; Hebert, J.R.; Mikhailidis, D.P.; Kengne, A.P.; Banach, M. Dietary inflammatory index and cardiometabolic risk in US adults. Atherosclerosis 2018, 276, 23–27. [Google Scholar] [CrossRef]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of potentially bioactive peptides generated from the enzymatic hydrolysis of hempseed proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef]
- Malomo, S.A.; He, R.; Aluko, R.E. Structural and functional properties of hemp seed protein products. J. Food Sci. 2014, 79, C1512–C1521. [Google Scholar] [CrossRef]
- Wang, X.S.; Tang, C.H.; Yang, X.Q.; Gao, W.R. Characterization, amino acid composition and in vitro digestibility of hemp (Cannabis sativa L.) proteins. Food Chem. 2008, 107, 11–18. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-inhibitory peptides from hemp seed (Cannabis sativa L.) proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; Alashi, A.; He, R.; Malomo, S.; Aluko, R.E. Preventive and treatment effects of a hemp seed (Cannabis sativa L.) meal protein hydrolysate against high blood pressure in spontaneously hypertensive rats. Eur. J. Nutr. 2014, 53, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, P.; Taka, E.; Bauer, D.; Cobourne-Duval, M.; Soliman, K.E. The attenuating effects of 1,2,3,4,6 penta-O-galloyl-β-Dglucose on inflammatory cytokines release from activated BV-2 microglial cells. J. Neuroimmunol. 2017, 305, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno-Silva, B.; Kawamoto, D.; Ando-Suguimoto, E.S.; Alencar, S.M.; Rosalen, P.L.; Mayer, M.P. Brazilian red propolis attenuates inflammatory signaling cascade in LPS activated macrophages. PLoS ONE 2015, 10, e0144954. [Google Scholar] [CrossRef] [PubMed]
- Santiago, K.B.; Conti, B.K.; Murbach Teles Andrade, B.F.; Mangabeira da Silva, J.J.; Rogez, H.L.; Crevelin, E.J.; Beraldo de Moraes, L.A.; Veneziani, R.; Ambrosio, S.R.; Bastos, J.K.; et al. Immunomodulatory action of Copaifera spp. oleoresins on cytokine production by human monocytes. Biomed. Pharmacother. 2015, 70, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Risitano, R.; Curro, M.; Cirmi, S.; Ferlazzo, N.; Campiglia, P.; Caccamo, D.; Ientile, R.; Navarra, M. Flavonoid fraction of bergamot juice reduces LPS induced inflammatory response through SIRT1-Mediated NF-kB inhibition in THP-1 monocytes. PLoS ONE 2014, 9, e107431. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kim, H.S.; Kim, Y.J.; Lee, H.K.; Kim, J.S.; Hong, J.T.; Kim, Y.; Hwang, B.Y.; Han, S.B. Effusanin C inhibits inflammatory responses via blocking NF-κB and MAPK signalingin monocytes. Int. Immunopharmacol. 2013, 15, 84–88. [Google Scholar] [CrossRef]
- Mohana, T.; Navin, A.V.; Jamuna, S.; Sakeena Sadullah, M.S.; Niranjali Devaraj, S. Inhibition of differentiation of monocyte to macrophages in atherosclerosis by oligomeric proanthocyanidins—In-vivo and in-vitro study. Food Chem. Toxicol. 2015, 82, 96–105. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, H.; Wang, X. Lunasin abrogates monocytes to endothelial cells. Mol. Immunol. 2017, 92, 146–150. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Lemus-Conejo, A.; Toscano, R.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. GPETAFLR, an octapeptide isolated from Lupinus angustifolius L. protein hydrolysate, promotes the skewing to the M2 phenotype in human primary monocytes. Food Funct. 2019, 10, 3303–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Target | GeneBank Accession Number | Forward Reverse | Sequence (5′ → 3′) |
|---|---|---|---|
| TNF-α | NM_000594 | Forward Reverse | TCCTTCAGACACCCTCAACC AGGCCCCAGTTTGAATTCTT |
| IL-1β | NM_000576 | Forward Reverse | GGGCCTCAAGGAAAAGAATC TTCTGCTTGAGAGGTGCTGA |
| IL-6 | NM_000600 | Forward Reverse | TACCCCCAGGAGAAGATTCC TTTTCTGCCAGTGCCTCTTT |
| IL-10 | NM_000572 | Forward Reverse | GCCTAACATGCTTCGAGATC TGATGTCTGGGTCTTGGTTC |
| IL-4 | NM_021283.2 | Forward Reverse | TCAACCCCCAGCTAGTTGTC TGTTCTTCGTTGCTGTGAGG |
| CCR7 | NM_007719.2 | Forward Reverse | GTGTGCTTCTGCCAAGATGA CCACGAAGCAGATGACAGAA |
| iNOS | NM_ 000625 | Forward Reverse | ACCCAGACTTACCCCTTTGG GCCTGGGGTCTAGGAGAGAC |
| CD200R | NM_138940.2 | Forward Reverse | GTTGCCCTCCTATCGCATTA TGGAAATTCCCATCAGGTGT |
| MRC1 | NM_ 138806 | Forward Reverse | GGCGGTGACCTCACAAGTAT ACGAAGCCATTTGGTAAACG |
| CCR2 | NM_001123396.1 | Forward Reverse | TGCCTGACTCACACTCAAGG GGCTTCTCAGCAACTGAACC |
| CCL2 | NM_002982.3 | Forward Reverse | CCCCAGTCACCTGCTGTTAT ACGAAGCCATTTGGTAAACG |
| HPRT | NM_001289746 | Forward Reverse | ACCCCACGAAGTGTTGGATA AAGCAGATGGCCACAGAACT |
| GADPH | NM_001289746 | Forward Reverse | CACATGGCCTCCAAGGAGTAAG CCAGCAGTGAGGGTCTCTCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Martin, N.M.; Montserrat-de la Paz, S.; Toscano, R.; Grao-Cruces, E.; Villanueva, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Hemp (Cannabis sativa L.) Protein Hydrolysates Promote Anti-Inflammatory Response in Primary Human Monocytes. Biomolecules 2020, 10, 803. https://doi.org/10.3390/biom10050803
Rodriguez-Martin NM, Montserrat-de la Paz S, Toscano R, Grao-Cruces E, Villanueva A, Pedroche J, Millan F, Millan-Linares MC. Hemp (Cannabis sativa L.) Protein Hydrolysates Promote Anti-Inflammatory Response in Primary Human Monocytes. Biomolecules. 2020; 10(5):803. https://doi.org/10.3390/biom10050803
Chicago/Turabian StyleRodriguez-Martin, Noelia M., Sergio Montserrat-de la Paz, Rocio Toscano, Elena Grao-Cruces, Alvaro Villanueva, Justo Pedroche, Francisco Millan, and Maria C Millan-Linares. 2020. "Hemp (Cannabis sativa L.) Protein Hydrolysates Promote Anti-Inflammatory Response in Primary Human Monocytes" Biomolecules 10, no. 5: 803. https://doi.org/10.3390/biom10050803