Successful Ecological Regeneration of Opencast Coal Mine Spoils through Forestation: From Cradle to Grove

 ,
,
Abstract
:
1. Introduction
1.1. The “Cradle for Nature” Strategy
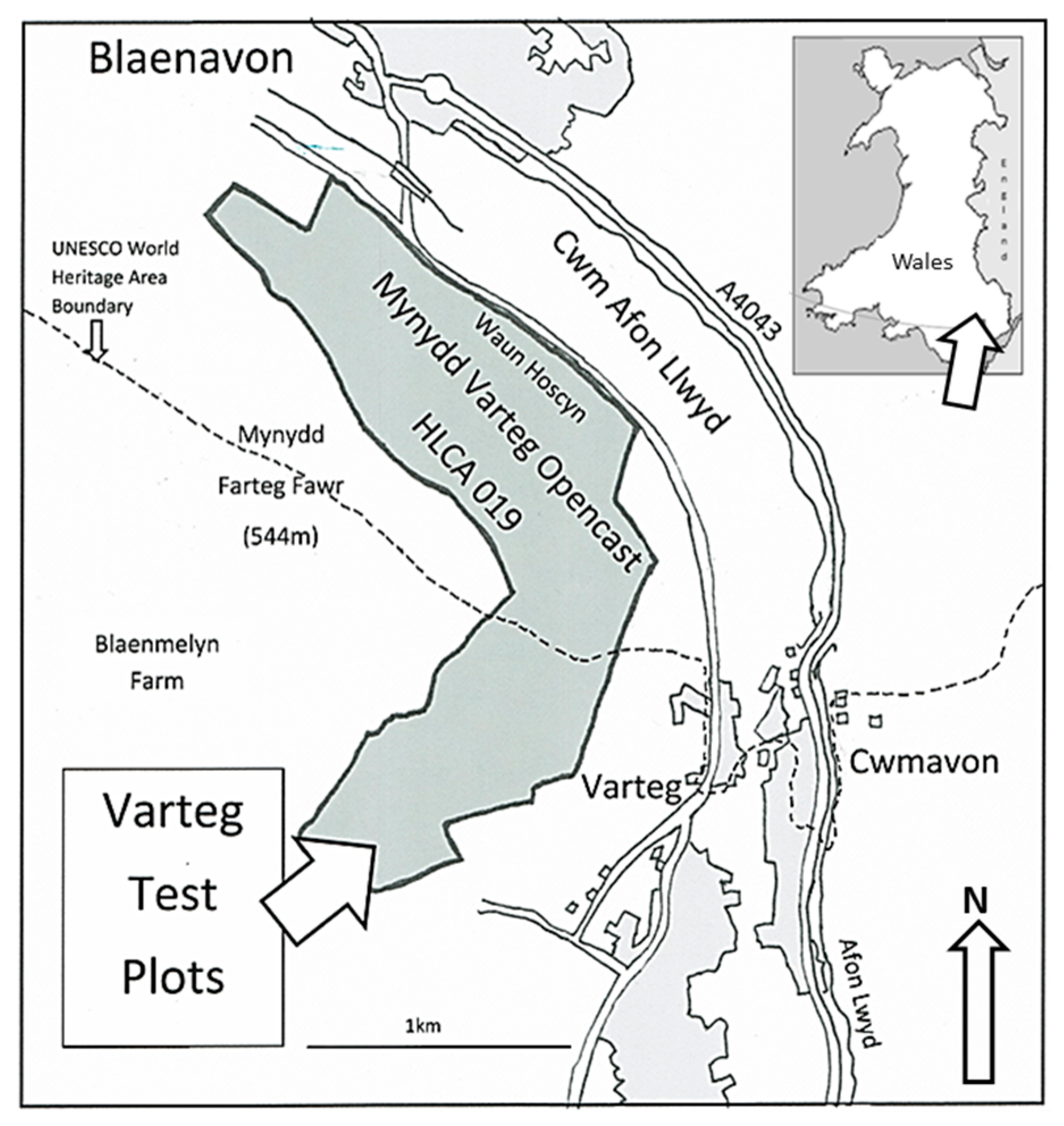
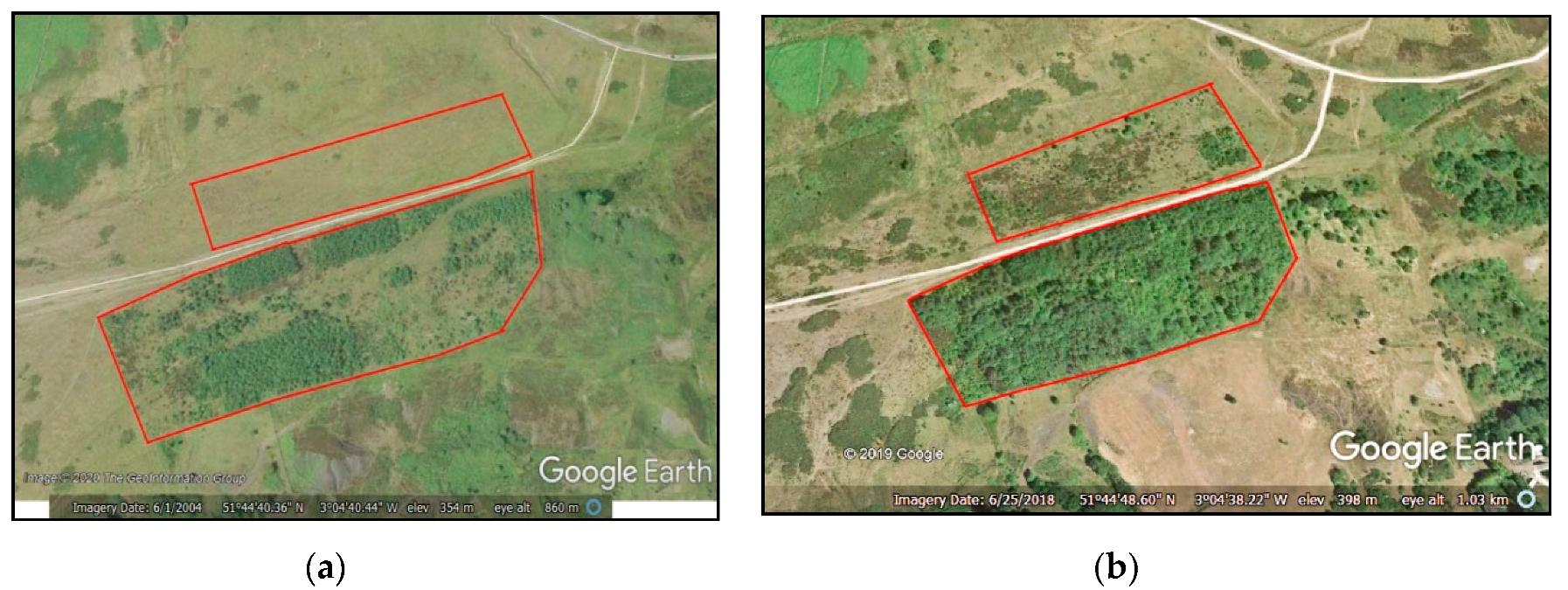
1.2. Test Site Location and Description
1.3. Method
1.4. Impact of Tree Planting on Ecology
2. Soils
2.1. Soil Quality and Physical Characteristics
2.2. Soil Density
2.3. Soil Contamination (Metals)
2.4. Soil Organic Matter and Microorganisms
2.5. Earthworms
3. Flora and Fauna
3.1. Vegetation and Ecological Succession
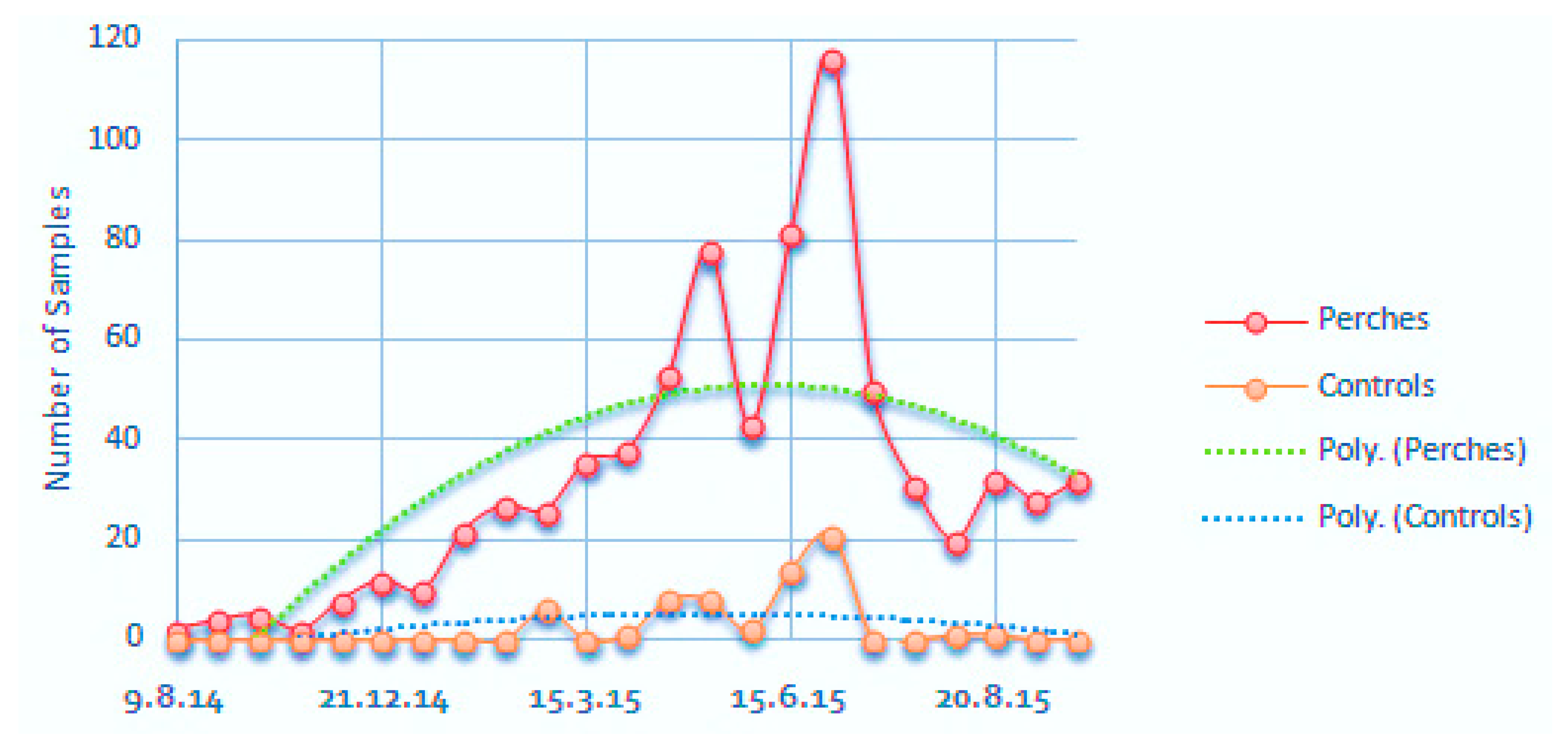
3.2. Volunteer Trees, Seed Rain and Bird Visitation
3.3. Self-Set Tree Species
3.4. Fauna
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scherer, D. Evolution, human living and the practice of ecological restoration. Environ. Ethics 1997, 17, 359–379. [Google Scholar] [CrossRef]
- Haigh, M. Chapter 32-Building a “Cradle for Nature”: A paradigm for environmental reconstruction. In Bio-Geotechnologies for Mine Site Rehabilitation; Prasad, M.N.V., de Campos, P.J., Maiti, S.K., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; pp. 593–616. [Google Scholar] [CrossRef]
- Monbiot, G. Big Coal’s Big Scam: Scar the Land for Profit and Let Others Pay; The Guardian: London, UK, 2015; p. 35. [Google Scholar]
- Milbourne, P.; Mason, K. Environmental injustice and post-colonial environmentalism: Opencast coal mining, landscape and place. Environ. Plan. Econ. Space 2017, 49, 29–46. [Google Scholar] [CrossRef]
- Welsh Government of Wales. Planning Policy Wales, 10th ed.; Government of Wales: Cardiff, UK, 2018; pp. 98–99.
- Haigh, M. The Aims of Land Reclamation. In Reclaimed Land: Erosion Control, Soils and Ecology (Land Reconstruct. Management); Haig, M., Ed.; A. A. Balkema (CRC Press): Rotterdam, The Netherlands, 2000; pp. 1–20. [Google Scholar]
- Robinson, J. Agricultural Land Classification and physical characteristics and soil forming materials survey: Pwll Du Opencast Coal Site, Blaenavon, Gwent; ADAS Land Management Services: Cardiff, UK, 1989. [Google Scholar]
- Torfaen County Borough. Blaenavon World Heritage Site Management Plan 2011–2016. 2011. Available online: https://www.visitblaenavon.co.uk/en/Publications/WorldHeritageSite/LookingAfterBlaenavon/BlaenavonWHSDraftManagementPlan(2011).pdf (accessed on 16 May 2020).
- Torfaen County Borough. Blaenavon Town Centre Conservation Area Appraisal and Management Plan 2017–2022. 2017. Available online: https://www.torfaen.gov.uk/en/Related-Documents/Planning/Conservation/Blaenavon-Town-Centre-Conservation-Area-Appraisal-and-Management-Plan-2017-2022.pdf (accessed on 16 May 2020).
- Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Angel, P.N.; Barton, C.D.; Davis, V.; Franklin, J.A. Restoring forests and associated ecosystem services on Appalachian coal surface mines. Environ. Manag. 2011, 47, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Q.; Shahrour, I.; Bai, Z.K.; Fan, W.X.; Feng, L.R.; Li, H.F. Soils development in opencast coal mine spoils reclaimed for 1–13 years in the West-Northern Loess Plateau of China. Eur. J. Soil Biol. 2013, 55, 40–46. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Bai, Z.; Reading, L. Effects of surface coal mining and land reclamation on soil properties: A review. Earth-Sci. Rev. 2019, 191, 12–25. [Google Scholar] [CrossRef]
- Haigh, M.J.; Reed, H.; D’Aucourt, M.; Flege, A.; Cullis, M.; Davis, S.; Farrugia, F.; Gentcheva-Kostadinova, S.; Zheleva, E.; Hatton, E.; et al. Effects of initial fertilizer treatment on the 10-year growth of mixed woodland on compacted surface-coal-mine spoils, S. W. ales. Land Degrad. Dev. 2018, 29, 3456–3468. [Google Scholar] [CrossRef]
- Zhang, H. Latin Square Design. In Encyclopedia of Research Design; Salkind, N.J., Ed.; SAGE: Thousand Oaks, CA, USA, 2010; pp. 99–704. [Google Scholar] [CrossRef]
- Haigh, M.; Desai, M.; Cullis, M.; D’Aucourt, M.; Sansom, B.; Wilding, G.; Alun, E.; Garate, S.; Hatton, L.; Kilmartin, M.; et al. Composted municipal green waste enhances tree success in opencast coal-land reclamation in Wales. Air Soil Water Res. 2019, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Plamping, K.; Haigh, M.; Reed, H.; Woodruffe, P.; Fitzpatrick, S.; Farrugia, F.; D’Aucourt, M.; Flege, A.; Sawyer, S.; Panhuis, W.; et al. Effects of initial planting method on the performance of mixed plantings of alder and oak on compacted opencast coal-spoils, Wales: 10-year results. Int. J. Min. Reclam. Environ. 2017, 31, 286–300. [Google Scholar] [CrossRef]
- Haigh, M.J. Soil Stewardship on reclaimed coal-lands. In Reclaimed Land: Erosion Control, Soils and Ecology; Haigh, M.J., Ed.; A. A. Balkema (CRC Press): Rotterdam, The Netherlands, 2000; pp. 165–274. [Google Scholar]
- Haigh, M.; Reed, H.; Flege, A.; D’Aucourt, M.; Plamping, K.; Cullis, M.; Woodruffe, P.; Sawyer, S.; Panhuis, W.; Wilding, G.; et al. Effect of planting method on the growth of Alnus glutinosa and Quercus petraea in compacted opencast coal-mine spoils, South Wales. Land Degrad. Dev. 2015, 26, 227–236. [Google Scholar] [CrossRef]
- Haigh, M.J.; Sansom, B. Soil compaction, runoff and erosion on reclaimed coal-lands (UK). Internat. J. Surf. Min. Reclam. Environ. 1999, 13, 135–146. [Google Scholar] [CrossRef]
- Haigh, M.J. Soil compaction on reclaimed surface coal-mine sites. J. Balk. Ecol. 1999, 2, 49–56. [Google Scholar]
- Sweigard, R.J.; Escobar, E. A field investigation into the effectiveness of reclamation equipment alternatives in reducing subsoil compaction. Min. Sci. Technol. 1989, 8, 313–320. [Google Scholar] [CrossRef]
- Ramsay, W.J. Bulk soil handling for quarry restoration. Soil Use Manag. 1986, 2, 30–39. [Google Scholar] [CrossRef]
- Baize, D. (Ed.) Referentiel Pedologique; AFES/INRA: Paris, France, 1992. [Google Scholar]
- Humphrey, M. An Investigation of the Impact of Soil Quality on Tree Growth on Land Reclaimed from Opencast Coal Mining at Varteg, South Wales. Master’s Thesis, Birmingham City University (University of Central England), Birmingham, UK, 1997. [Google Scholar]
- Desai, M. Phytoremediation: A Tool for Restoring Land Degraded due to Open Cast Coal Mining. Ph.D. Thesis, Oxford Brookes University, Oxford, UK, 2014. Available online: https://ethos.bl.uk/OrderDetails.do?uin=uk.bl.ethos.634790 (accessed on 16 May 2020).
- Haigh, M.; Kilmartin, M.P. Illustrating the water quality impacts of reclaimed opencast coal lands, SE Wales. In Management of Municipal Watersheds in Mountain Regions; Proceedings of the 31st Session, Prague, Czech Republic, 4–6 September 2017; Palán, L., Křeček, J., Eds.; EFC/FAO Working Party on the Management of Mountain Watersheds, Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2017; pp. 87–115. [Google Scholar]
- Smart, P.; Simpson, A. (Eds.) Varteg Coal Recovery and Reclamation Scheme, Blaenavon, South Wales, Hydrogeology and Contaminated Land Assessment, Final Interpretive Report; Faber-Maunsell AECOM: Birmingham, UK, 2006. [Google Scholar]
- Dang, Z.; Liu, C.Q.; Haigh, M.J. Mobility of heavy metals associated with the natural weathering of coal mine spoils. Environ. Pollut. 2002, 118, 419–426. [Google Scholar] [CrossRef]
- Desai, M.; Haigh, M.; Walkington, H. Phytoremediation: Metal decontamination of soils after the sequential forestation of former opencast coal land. Sci. Total Environ. 2019, 656, 670–680. [Google Scholar] [CrossRef]
- MAFF. Towards Sustainable Agriculture: A Pilot Set of Indicators; Ministry of Agriculture, Fisheries and Food (MAFF), Her Majesty’s Stationery Office: London, UK, 2000.
- Harris, J. Measurements of the soil microbial community for estimating the success of restoration. Eur. J. Soil Sci. 2003, 4, 801–808. [Google Scholar] [CrossRef]
- Filcheva, E.; Noustorova, M.; Gentcheva-Kostadinova, S.V.; Haigh, M.J. Impact of forestation on organic accumulation and microbial action in surface coal-mine spoils, Pernik, Bulgaria. Ecol. Eng. 2000, 15, 1–15. [Google Scholar] [CrossRef]
- Filcheva, E.; Hristova, M.; Noustorova, M.; Malcheva, B.; Haigh, M. Soil Microbiological Activity and Organic Matter in Technosols of Wales; Institute of Soil Science, Agrotechnology and Plant Protection “N. Poushkarov”and University of Forestry: Sofia, Bulgaria, 2019; paper in preparation. [Google Scholar]
- Noustorova, M.; Filcheva, E.; Gencheva, S.; Zheleva, E. Microbiological characteristics and composition of organic matter in materials from coal mining in Wales, United Kingdom. J. Balk. Ecol. 1999, 2, 57–61. [Google Scholar]
- Scullion, J.; Malik, A. Earthworm activity affecting organic matter, aggregation and microbial activity in soils restored after opencast mining for coal. Soil Biol. Biochem. 2000, 32, 119–126. [Google Scholar] [CrossRef]
- Butt, K.R. Inoculation of earthworms into reclaimed soils: The UK experience. Land Degrad. Dev. 1999, 10, 565–575. [Google Scholar] [CrossRef]
- Butt, K.R. Earthworms in soil restoration: Lessons learned from United Kingdom case studies of land reclamation. Restor. Ecol. 2008, 16, 637–641. [Google Scholar] [CrossRef]
- Drinkwater, E.; Robinson, E.J.; Hart, A.G. Keeping invertebrate research ethical in a landscape of shifting public opinion. Methods Ecol. Evol. 2019, 10, 1265–1273. [Google Scholar] [CrossRef]
- Lawrence, A.P.; Bowers, M.A. A test of the “hot”mustard extraction method of sampling earthworms. Soil Biol. Biochem. 2002, 34, 549–552. [Google Scholar] [CrossRef]
- Bartlett, M.D.; Harris, J.A.; James, I.T.; Ritz, K. Inefficiency of mustard extraction technique for assessing size and structure of earthworm communities in UK pasture. Soil Biol. Biochem. 2006, 38, 2990–2992. [Google Scholar] [CrossRef]
- Bonham, C.D. Measurements for Terrestrial Vegetation, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2013; pp. 19–40. [Google Scholar]
- Rodwell, J.S. British Plant Communities Volume 3: Grasslands and Montane Communities; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Evans, T. Flora of Monmouthshire; Chepstow Society; Chepstow Museum: Chepstow, UK, 2007; ISBN 0 900278 49 8. [Google Scholar]
- ACW. Wye Valley Surveys. In Land Reclamation/Coal Recovery Scheme, Varteg Hill, Torfaen: Environmental Statement; Glamorgan Power for Torfaen County Borough Council: Cwmbran, UK, 2004. [Google Scholar]
- Rodwell, J.S. British Plant Communities Volume 2: Heathlands and Mires; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Coulson, C.; Spooner, P.G.; Lunt, I.D.; Watson, S.J. From the matrix to roadsides and beyond: The role of isolated paddock trees as dispersal points for invasion. Divers. Distrib. 2014, 20, 137–148. [Google Scholar] [CrossRef]
- Toh, I.; Gillespie, M.; Lamb, D. The role of isolated trees in facilitating tree seedling recruitment at a degraded sub-tropical rainforest site. Restor. Ecol. 1999, 7, 288–297. [Google Scholar] [CrossRef]
- Zweiner, V.P.; Cardoso, F.C.G.; Padial, A.A.; Marques, M.C.M. Disentangling the effects of facilitation on restoration of the Atlantic Forest. Basic Appl. Ecol. 2014, 15, 34–41. [Google Scholar] [CrossRef]
- Bustamante-Sánchez, M.A.; Armesto, J.J.; Landis, D. Seed limitation during early forest succession in a rural landscape on Chiloé Island, Chile: Implications for temperate forest restoration. J. Appl. Ecol. 2012, 49, 1103–1112. [Google Scholar] [CrossRef] [Green Version]
- Graham, L.L.B.; Page, S.E. Artificial bird perches for the regeneration of degraded tropical peat swamp forest: A restoration tool with limited potential. Restor. Ecol. 2012, 20, 631–637. [Google Scholar] [CrossRef]
- Heeleman, S.; Krug, C.B.; Esler, K.J.; Reisch, C.; Poschlod, P. Pioneers and perches: Promising restoration methods for degraded renosterveld habitats? Restor. Ecol. 2012, 20, 18–23. [Google Scholar] [CrossRef]
- Snow, B.; Snow, D. Birds and Berries: A Study of an Ecological Interaction; T & AD Poyser: London, UK, 1988. [Google Scholar]
- Dement, W.T.; Hackworth, Z.J.; Lhotka, J.M.; Barton, C.D. Plantation development and colonization of woody species in response to post-mining spoil preparation methods. New For. 2020, 1–20. [Google Scholar] [CrossRef]
- Glamorgan Power. Chapter12: Ecology. In Environmental Statement: Proposed Land Reclamation/Coal Recovery Scheme at Varteg Hill, Torfaen; Glamorgan Power for Torfaen County Borough Council: Cwmbran, UK, 2014. [Google Scholar]
- Mathews, F.; Kubasiewicz, L.M.; Gurnell, J.; Harrower, C.A.; McDonald, R.A.; Shore, R.F. A Review of the Population and Conservation Status of British Mammals; 2018 A review of the population and conservation status of British mammals. (Natural England Joint Publication JP025, CEH Project no. C06029, C06940); Natural England: Peterborough, UK, 2018; Available online: http://nora.nerc.ac.uk/id/eprint/520584/ (accessed on 16 May 2020).
- Poland Bowen, C. Mammals UK: An Introduction to the Nation’s Wild Mammals; People’s Trust for Endangered Species: London, UK, 2001. [Google Scholar]
- Aderyn. Website Home; LERC Wales’ Biodiversity Information and Reporting Database: Brecon, UK. Available online: https://aderyn.lercwales.org.uk/ (accessed on 25 March 2020).
- D’Aucourt, M. Small Mammal Surveys.pdf. Available online: http://cradlefornature.org.uk/documents/ (accessed on 16 May 2020).
- D’Aucourt, M. Survey using Hedgehog Tunnel and Trail Cameras on Varteg plots 2015 & 2016. Available online: http://cradlefornature.org.uk/documents/ (accessed on 16 May 2020).
- D’Aucourt, M. Report of Mammal Survey at Varteg in July 2018. Available online: http://cradlefornature.org.uk/documents/ (accessed on 16 May 2020).
- Strachan, R. Mammal Detective; Whittet Books: London, UK, 1995. [Google Scholar]
- Anon. What Animal Poo Is this? The Wildlife Trusts: Newark, UK; Available online: https://www.wildlifetrusts.org/wildlife/how-identify/identify-poo (accessed on 16 May 2020).
- Gurnell, J.; Flowerdew, J.R. Live Trapping Small Mammals; A Practical Guide, 4th ed.; The Mammal Society: London, UK, 2019; Available online: https://www.mammal.org.uk/wp-content/uploads/2019/07/Trapping-Guide-Additional-Information-2019.pdf (accessed on 16 May 2020).
- Thomas, E.; Wilson, E. Guidance-for-Detecting-Hedgehogs-Using-Tracking-Tunnels; People’s Trust for Endangered Species: London, UK, 2019; Available online: https://ptes.org/wp-content/uploads/2015/06/Guidance-for-detecting-hedgehogs-using-tracking-tunnels.pdf (accessed on 16 May 2020).
- Middleton-Burke, A. Feasibility of footprint tracking tubes to detect presence of the hazel dormouse. Mammal News 2017, 178, 1–2. [Google Scholar]
- Cornally, A.; Lawton, C.A. Guide to the identification of Irish mammal hair. National Parks and Wildlife Service. In Irish Wildlife Manuals; National Parks and Wildlife Service: Dublin, Ireland, 2016; p. 92. [Google Scholar]
- Styles, H.T. A Guide to British Mammal Tracks and Signs; Wildlife World: London, UK, 2001. [Google Scholar]
- Bending, N.A.D.; Moffat, A.J. Tree performance on minespoils in the South Wales coalfield. J. Appl. Ecol. 1999, 36, 784–797. [Google Scholar] [CrossRef]
- Hale, K.; Spencer, M.; Peterken, G.F.; Mountford, E.P.; Bradshaw, R.H. Rapid carbon accumulation within an unmanaged, mixed, temperate woodland. Scand. J. For. Res. 2019, 34, 208–217. [Google Scholar] [CrossRef]
- Margenau, E.L.; Wood, P.B.; Weakland, C.A.; Brown, D.J. Trade-offs relating to grassland and forest mine reclamation approaches in the central Appalachian region and implications for the songbird community. Avian Conserv. Ecol. 2019, 14. [Google Scholar] [CrossRef]
- López-Marcos, D.; Turrión, M.B.; Martínez-Ruiz, C. Linking soil variability with plant community composition along a mine-slope topographic gradient: Implications for restoration. Ambio 2020, 49, 337–349. [Google Scholar] [CrossRef]
- Alday, J.G.; Marrs, R.H.; Martínez-Ruiz, C. Soil and vegetation development during early succession on restored coal wastes: A six-year permanent plot study. Plant Soil 2012, 353, 305–320. [Google Scholar] [CrossRef]
- Hayhow, D.B.; Eaton, M.A.; Stanbury, A.J.; Burns, F.; Kirby, W.B.; Bailey, N.; Beckmann, B.; Bedford, J.; Boersch-Supan, P.H.; Coomber, F.; et al. State of Nature 2019; National Biodiversity Network (State of Nature Partnership): Nottingham, UK, 2019; Available online: https://nbn.org.uk/wp-content/uploads/2019/09/State-of-Nature-2019-UK-full-report.pdf (accessed on 16 May 2020).
- Walley, C. Carving out a Future; Rural Wales Magazine CPRW (Campaign for the Protection of Rural Wales): Welshpool, Powys, UK, 1994; pp. 22–24. [Google Scholar]
- Flege, A. Recovering the Surface-Man-Made Soil at a Reclaimed Site in the South Wales Coalfield. Ph.D. Thesis, Department of Geography, McMicken College, University of Cincinnati, Cincinnati, OH, USA, 2003. [Google Scholar]
- Shaxson, T.F. New Concepts and Approaches to Land Management in the Tropics with Emphasis on Steep Lands; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 1999; Available online: http://www.fao.org/family-farming/detail/en/c/329766/ (accessed on 16 May 2020).
- Frouz, J.; Pižl, V.; Tajovský, K. The effect of earthworms and other saprophagous macrofauna on soil microstructure in reclaimed and un-reclaimed post-mining sites in Central Europe. Eur. J. Soil Biol. 2007, 43, S184–S189. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years Since Disturbance | <1 | 15–25 | 35 | 50 |
|---|---|---|---|---|
| Reclaimed Opencast Site | Varteg (New Trench < 1 year) | Blaenant (1974–1982) | Varteg (1963) | Pwll Du (1948) |
| Depth 0–5 cm | n.a. | 1.35* (0.21) | 1.41** (0.19) | 1.51* (0.18) |
| Depth 5–10 cm | 1.41* | 1.82* (0.21) | 1.69* (0.19) | 1.69* (0.22) |
| Depth 10–20 cm | 0.81 | 1.81 (0.20) | 1.73 (0.30) | 1.75 (0.18) |
| Depth 20–30 cm | 1.14 | 1.79 (0.21) | 1.73 (0.18) | 1.74 (0.11) |
| Depth 30–40 cm | 1.48 | 1.82 (0.22) | 1.79 (0.19) | 1.79 (0.19) |
| Depth 40–50 cm | 1.21 | 1.62** | 1.81** (0.19) | 1.81** (0.19) |
| Depth 50–60 cm | 1.61 | 1.81 (0.22) | 1.76 (0.19) | 1.76 (0.19) |
| Mean Bulk Density (g·cm3) 0–50 cm | 1.16 | 1.81 (0.22) | 1.76 (0.19) | 1.74 (0.17) |
| Sample (n =) | 2 (*1) | 42 (21*, 2**) | 22 (11*, 7**) | 58 (*26;**14) |
| Comparison | Probability | Interpretation |
|---|---|---|
| alder vs. open grassland | <0.01 | greater worm biomass under alder |
| under pit- vs. notch-planted alder | 0.02 | greater worm biomass under pit plantings |
| under trench- vs. notch-planted alder | 0.02 | greater worm biomass under trench plantings |
| under pit- vs. trench-planted alder | ns | ns: no significant difference |
| Earthworms | IT vs. C | OT vs. C | IT vs. OT |
|---|---|---|---|
| Total number | T = 10 N = 11 p = 0.05 | T = 6 N = 11 p < 0.02 | T = 10.5 N = 11 ns |
| Total biomass | T = 2 N = 11 p < 0.005 | T = 1 N = 11 p <0.005 | T = 22 N = 11 ns |
| Number of large worms | T = 2 N = 11 p < 0.005 | T = 0 N = 7 p = 0.02 | T = 5 N = 6 ns |
| Biomass of large worms | T = 2 N = 11 p < 0.005 | T = 1 N = 8 p < 0.02 | T = 30 N = 11 ns |
| Number of mature worms | T = 5.5 N = 10 p < 0.05 | T = 4 N = 11 p < 0.01 | T = 9 N = 9 ns |
| Mean weight of all worms | T = 0 N = 11 p < 0.001 | T = 1 N = 11 p < 0.005 | T = 19 N = 11 ns |
| Direction of difference | IT > C | OT > C | IT = OT |
| Species | Site A | Site B | Site C | ||||
|---|---|---|---|---|---|---|---|
| Unfenced, Grazed Grassland, Undisturbed/No Tree Planting | Fenced, Ungrazed Forest Plantations with Associated Disturbance | ||||||
| Year 1 | Year 14 | Year 21 | Year 1 | Year 14 | Year 21 | Year 14 | |
| Species characteristic of U4 grassland | |||||||
| Agrostis species | 29 | 20 | 26 | 30 | 27 | 15 | |
| Anthoxanthum odoratum | 12 | 17 | 2 | ||||
| Festuca species | 29 | 27 | 27 | 21 | 8 | 6 | 8 |
| Nardus stricta | 16 | 27 | 25 | 0 | 0 | 0 | 16 |
| Galium saxatile | 23 | 26 | 12 | 6 | 8 | 6 | 8 |
| Potentialla erecta | 12 | 15 | 22 | ||||
| Scrub species | |||||||
| Ulex species | 6 | 16 | 21 | ||||
| Calluna vulgaris | 3 | 3 | 14 | 24 | |||
| Vaccinium myrtilis | 4 | 22 | 17 | 9 | |||
| Woodland species excluding planted trees: | |||||||
| Hedera helix | 24 | ||||||
| Crataegus monogyna | 3 | 12 | |||||
| Quercus species | 3 | 9 | |||||
| Betula pendula | 12 | ||||||
| Alnus species | 15 | ||||||
| Rubus fruticosus | 14 | 8 | |||||
| Moss species | 29 | 30 | 27 | 30 | 30 | 30 | |
| Lichen species | 18 | 14 | 15 | ||||
| Bare Ground | 0% | 0% | <5% | 65% | NDA | 9% | 17% |
| Totals of all species recorded during the surveys: | |||||||
| Grasses, rushes and sedges | 10 | 11 | 11 | 7 | 5 | 3 | 13 |
| Herbs | 13 | 15 | 10 | 9 | 8 | 6 | 25 |
| Woody species | 2 | 3 | 2 | 1 | 1 | 4 | 8 |
| Total All Species | 25 | 29 | 23 | 17 | 14 | 13 | 46 |
| Species | Local Records Exist/Expected to Occur in South Wales | Evidence |
|---|---|---|
| Insectivores | ||
| Hedgehog Erinaceus europaeus | [54,55,56,57] | incidental sighting of dead one [58] |
| Mole Talpa europaea | [54,55,56,57] | incidental sighting of half-eaten one under trees [59] |
| Common Shrew Sorex araneus | [55,56,57] | in Longworth traps [58]; probable prints in track tubes on the ground [59] |
| Pygmy Shrew Sorex minutus | [55,56] | in Longworth traps [58] |
| Water Shrew Neomys fodiens | [55,56] Riparian, so unlikely to be on site | |
| Rodents | ||
| Grey Squirrel Sciurus carolinensis | [55,56,57] | signs: nibbled pine cones, opened acorn shell and oak debarking [60] |
| Bank Vole Clethrionomys glareolus | [55,56] | in Longworth traps [58]; possible droppings in track tubes on the ground [60,61,62] |
| Field Vole Microtus agrestis | [55,56,57] | in Longworth traps [58]; live in open control area [60]; signs of runs and holes along fencing [60] |
| Water Vole Arvicola terrestris | [56] Riparian, so unlikely to be on site | |
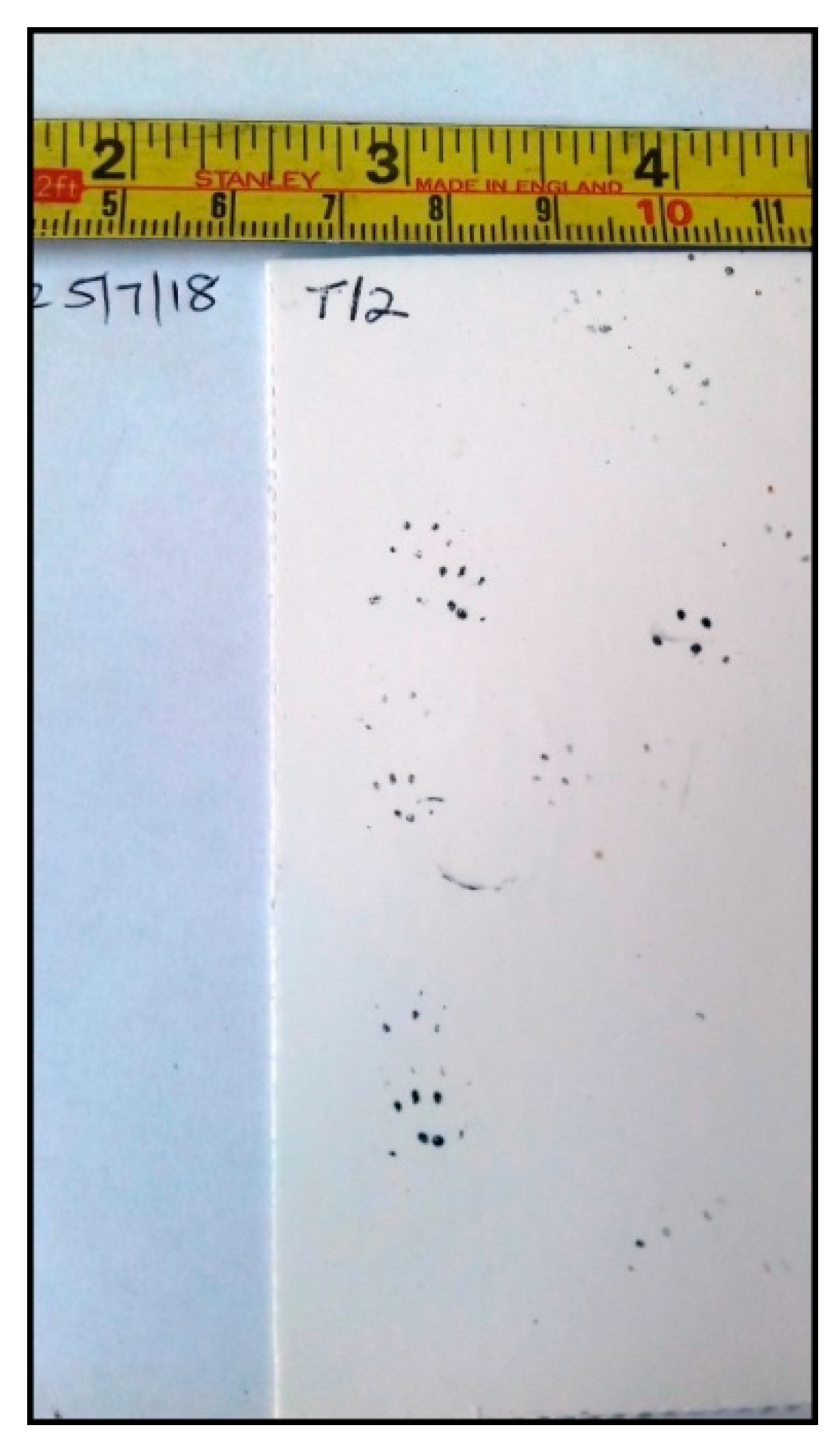
| Wood Mouse Apodemus sylvaticus | [55,56,57] | in Longworth traps [58]; camera trap photos [59]; prints in tubes and tunnel [59,60,61] (Figure 9, Figure 10 and Figure 11)/possible hair in hair tube [60], possible droppings in track tubes on the ground [60,61,62] |
| House Mouse Mus musculus | [55,56] Associated with dwellings, so unlikely here | |
| Brown Rat Rattus norvegicus | [55,56,57] | |
| Hazel Dormouse Muscardinus avellanarius | [55] Less likely, as fragmented habitat | |
| Yellow-necked Mouse Apodemus flavicollis | [55] Less likely, as edge of known range | |
| Carnivores | ||
| Red Fox Vulpes vulpes | [55,56,57] | probable droppings near tubes on the ground [60,61,62]; probable droppings on open ground near white downy feathers [60,61,62]; possible predator of the mole, corvid and the Guinea fowl [60] |
| Pine Marten Martes martes | [55] Less likely, as edge of known range | |
| Stoat Mustela erminea | [55,56,57] | |
| Weasel Mustela nivalis | [55,56,57] | |
| Possible small mustelid | blurred camera trap photo [59] | |
| Polecat Mustela putorius | [55,56] | |
| Badger Meles meles | [54,55,56] Local road-kills | |
| Otter Lutra lutra | [54,56] Riparian, so unlikely to be on site (but in Cwm Afon Llwyd and local road-kill) | |
| American Mink Mustela vison | [55,56] Riparian, so unlikely to be on site | |
| Deer | ||
| Roe Capreolus capreolus | [55,57] | |
| Fallow Dama dama | [55] | |
| Reeves’ Muntjac Muntiacus reevesi | [55] | |
| Lagomorphs | ||
| European Rabbit Oryctolagus cuniculus | [55,56,57] | probable fur and pieces of bone extracted from bird pellets [60]; incidental droppings [60,61,62]; incidental skeleton at base of tree [59] |
| Brown Hare Lepus europaeus | [54,55,56,57] Local records | dead, hanging in a tree [60] |
| Other animals | ||
| Blackbird Turdus merula | camera trap photo [59] | |
| European robin Erithacus rubecula | camera trap video of it on track tunnel roof [59] | |
| Helmeted Guineafowl Numida meleagris | Non-native | incidental articulated wing in open area [60] fairly whole, articulated leg on the ground under the trees on same day [60] |
| Corvid | incidental articulated black primary feathers on the ground under the trees [60] | |
| Owl or bird of prey | fur balls under white-splashed post in mixed tree/open area [60,62]; white downy feathers on the open ground [60] | |
| Common Frog Rana temporaria | in a Longworth trap [58] | |
| Common lizard Lacerta vivipara | live in the open control area [58] | |
| Wasp | old nest low in hawthorn [60] | |
| Attribute | Degraded Land in Theory | Rehabilitated Land in Theory | Evidence from the Varteg |
|---|---|---|---|
| Biomass | Low | High | Increasing |
| Carbon sequestration | Low | High | Increasing (0–15 cm depth) |
| Soil depth | Decreasing | Increasing | Development of humus horizon |
| Soil density | Extreme (usually high) | Equable | Less extreme (>1.7 down to <1.5 g·cm3) |
| Soil chemistry | Extreme | Equable | Reduction in metal contaminant loadings (by>35% over 14 years for 5 key metals) |
| Soil biological activity | Low | Higher | Increasing earthworm biomass and trees especially under older trees; expanding microbial system, with new emphasis on bacilli |
| Hydrological state | More variable | More equable | Improving—less surface runoff, more infiltration and more soil moisture storage [16] |
| Plant-available water | Low | Higher | Improving due to increased organic matter in soil and better soil structure |
| Ecological structure | Simple | Complex | Transition from grassland to woodland |
| Undecomposed debris as proportion of organic matter | High | Low | Very high initially, now lower due to increasing activity and recycling in the soil ecological system |
| Ecosystem niches | Low number of ecological niches | High number of ecological niches | Number of ecological niches increased greatly during the phased, annual planting, transition from grassland to woodland and consequent creation of habitat mosaic |
| Biodiversity | Usually lower | Usually higher | Increased due to the creation of the habitat mosaic on site, but probably reducing as the site matures to dense forest |
| Life cycles | Short | Long | Grass to trees |
| Population regulation | Mainly by physical environmental factors—heat drought, etc. | More by biologically mediated factors | Forest microclimate ameliorates the impact of physical environment, while deeper, more organically enriched, soils buffer moisture change. Forest mosaic increases opportunities for interspecific competition |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haigh, M.; Woodruffe, P.; D’Aucourt, M.; Alun, E.; Wilding, G.; Fitzpatrick, S.; Filcheva, E.; Noustorova, M. Successful Ecological Regeneration of Opencast Coal Mine Spoils through Forestation: From Cradle to Grove. Minerals 2020, 10, 461. https://doi.org/10.3390/min10050461
Haigh M, Woodruffe P, D’Aucourt M, Alun E, Wilding G, Fitzpatrick S, Filcheva E, Noustorova M. Successful Ecological Regeneration of Opencast Coal Mine Spoils through Forestation: From Cradle to Grove. Minerals. 2020; 10(5):461. https://doi.org/10.3390/min10050461
Chicago/Turabian StyleHaigh, Martin, Patricia Woodruffe, Margaret D’Aucourt, Elanor Alun, Gillian Wilding, Susan Fitzpatrick, Ekaterina Filcheva, and Maya Noustorova. 2020. "Successful Ecological Regeneration of Opencast Coal Mine Spoils through Forestation: From Cradle to Grove" Minerals 10, no. 5: 461. https://doi.org/10.3390/min10050461