MicroRNA-1253 Regulation of WASF2 (WAVE2) and its Relevance to Racial Health Disparities

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Microarray, Target Prediction, and Pathway Analysis
2.3. Cell Culture and Transfection
2.4. The 3′ UTR Luciferase Reporter Assays
2.5. RNA Isolation and RT-qPCR
2.6. Western Blot Analysis
2.7. Immunofluorescence and Scoring of Cells with Lamellipodia and Filopodia
2.8. Statistical Analysis
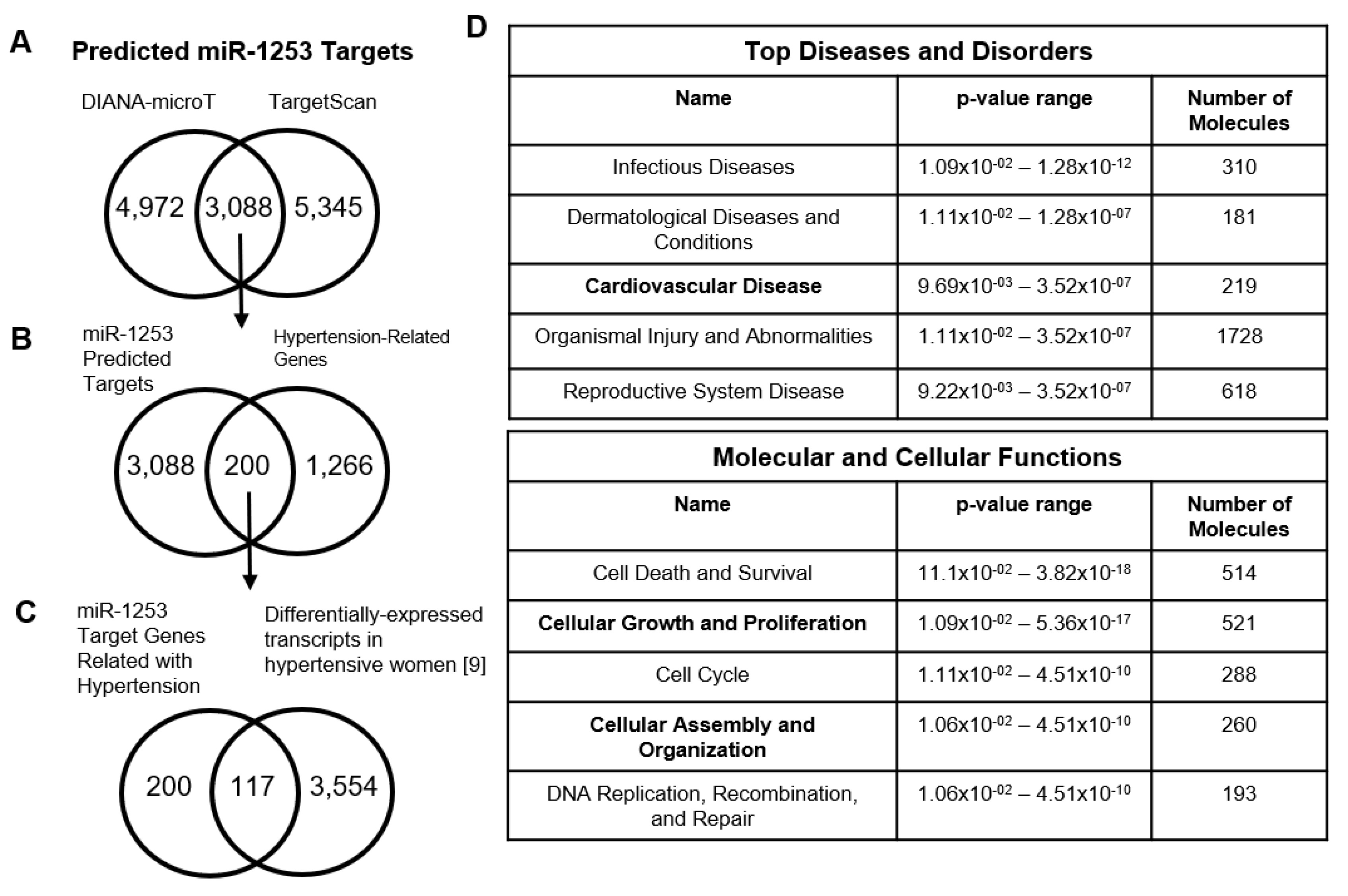
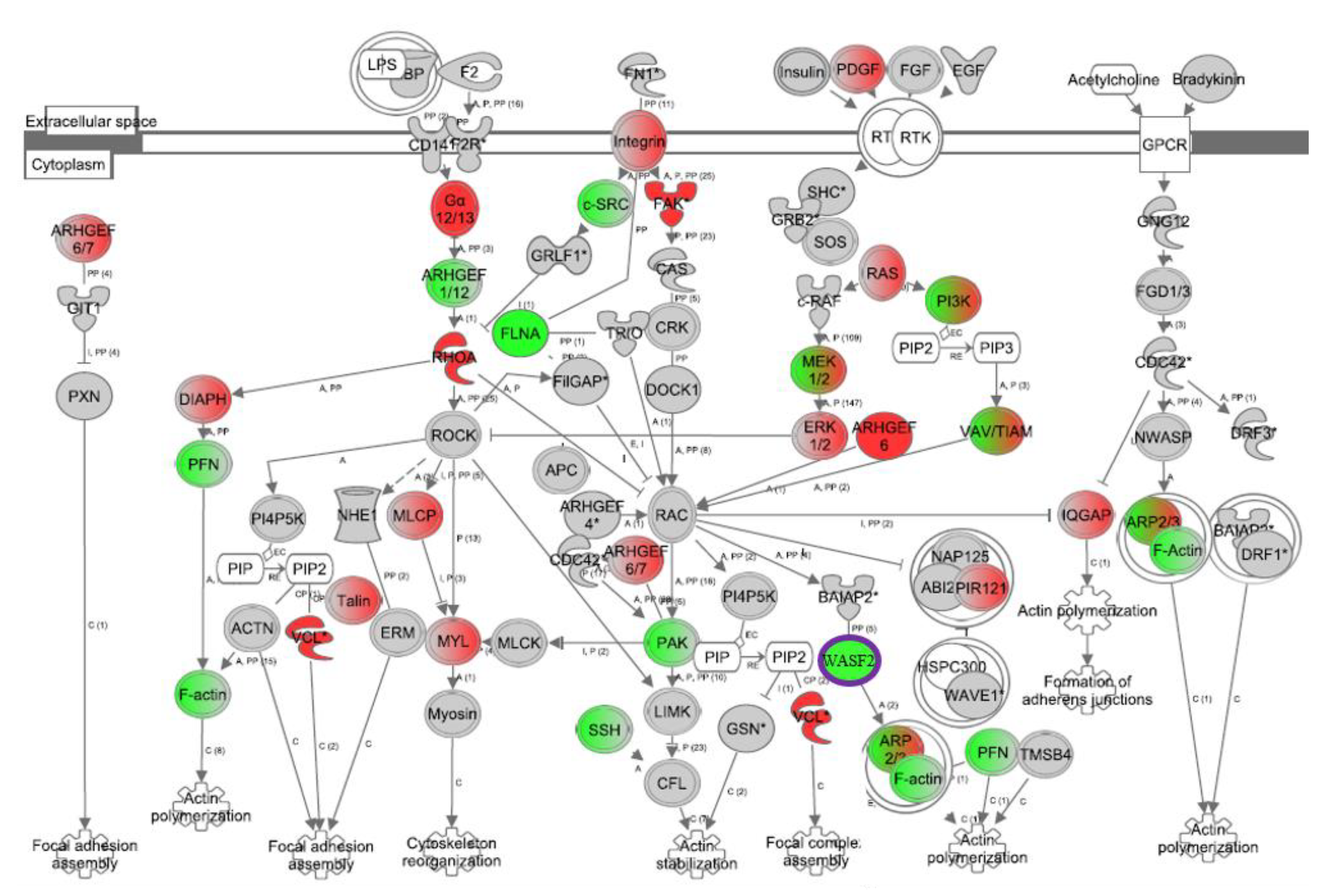
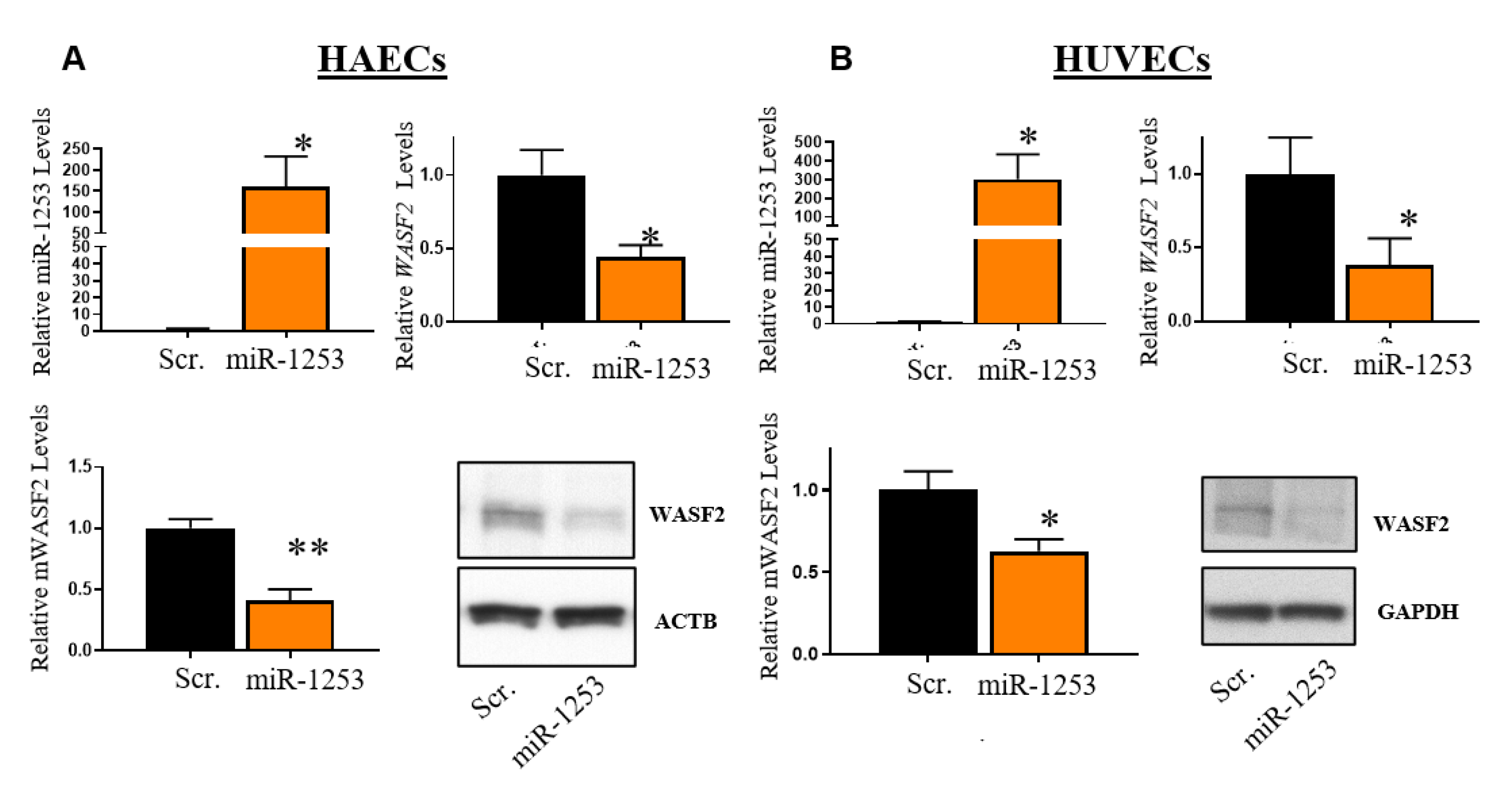
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Muntner, A.; Alonso, M.S.; Bittencourt, C.W.; Callaway, A.P.; Carson, A.M.; Chamberlain, A.R.; Chang, S.; Cheng, S.R.; Das, F.N.; et al. Virani, Epidemiology American Heart Association Council on, Committee Prevention Statistics, and Subcommittee Stroke Statistics. Heart Disease and Stroke Statistics-2019 Update: A Report from the American Heart Association. Circulation 2019, 139, E56–E528. [Google Scholar] [CrossRef]
- Patel, S.A.; Winkel, M.; Ali, M.K.; Narayan, K.V.; Mehta, N.K. Cardiovascular mortality associated with 5 leading risk factors: National and state preventable fractions estimated from survey data. Ann. Intern. Med. 2015, 163, 245. [Google Scholar] [CrossRef]
- Nédélec, Y.; Sanz, J.; Baharian, G.; Szpiech, Z.A.; Pacis, A.; Dumaine, A.; Grenier, J.-C.; Freiman, A.; Sams, A.J.; Hebert, S.; et al. Genetic Ancestry and Natural Selection Drive Population Differences in Immune Responses to Pathogens. Cell 2016, 167, 657–669.e21. [Google Scholar] [CrossRef]
- Thames, A.D.; Irwin, M.R.; Breen, E.C.; Cole, S.W. Experienced discrimination and racial differences in leukocyte gene expression. Psychoneuroendocrinology 2019, 106, 277–283. [Google Scholar] [CrossRef]
- Wang, B.-D.; Ceniccola, K.; Yang, Q.; Andrawis, R.; Patel, V.; Ji, Y.; Rhim, J.; Olender, J.; Popratiloff, A.; Latham, P.; et al. Identification and Functional Validation of Reciprocal microRNA-mRNA Pairings in African American Prostate Cancer Disparities. Clin. Cancer Res. 2015, 21, 4970–4984. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.; Parker, N.E.; Montgomery, P.S.; Sosnowska, D.; Casanegra, A.I.; Ungvari, Z.; Csiszar, A.; Sonntag, W.E. Gender and racial differences in endothelial oxidative stress and inflammation in patients with symptomatic peripheral artery disease. J. Vasc. Surg. 2014, 61, 1249–1257. [Google Scholar] [CrossRef] [Green Version]
- Moayedi, Y.; Fan, C.S.; Miller, R.J.H.; Tremblay-Gravel, M.; Posada, J.G.D.; Manlhiot, C.; Hiller, D.; Yee, J.; Woodward, R.; McCaughan, J.A.; et al. Gene Expression Profiling and Racial Disparities in Outcomes after Heart Transplantation. J. Heart Lung Transpl. 2019, 38, 820–829. [Google Scholar] [CrossRef] [Green Version]
- Dluzen, D.F.; Kim, Y.; Bastian, P.; Zhang, Y.; Lehrmann, E.; Becker, K.G.; Hooten, N.N.; Evans, M.K. MicroRNAs Modulate Oxidative Stress in Hypertension through PARP-1 Regulation. Oxid. Med. Cell. Longev. 2017, 2017, 3984280. [Google Scholar] [CrossRef] [Green Version]
- Dluzen, D.F.; Hooten, N.N.; Zhang, Y.; Kim, Y.; Glover, F.E.; Tajuddin, S.M.; Jacob, K.D.; Zonderman, A.; Evans, M.K. Racial differences in microRNA and gene expression in hypertensive women. Sci. Rep. 2016, 6, 35815. [Google Scholar] [CrossRef]
- Liang, J.; Le, T.H.; Edwards, D.R.V.; Tayo, B.O.; Gaulton, K.J.; Smith, J.A.; Lu, Y.; Jensen, R.A.; Chen, G.; Yanek, L.R.; et al. Single-Trait and Multi-Trait Genome-Wide Association Analyses Identify Novel Loci for Blood Pressure in African-Ancestry Populations. PLoS Genet. 2017, 13, e1006728. [Google Scholar] [CrossRef]
- Ness, R.B.; Haggerty, C.L.; Harger, G.; Ferrell, R. Differential Distribution of Allelic Variants in Cytokine Genes among African Americans and White Americans. Am. J. Epidemiol. 2004, 160, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, S.; Ebert, M.S.; Zheng, G.X.Y.; Tsang, J.S.; Sharp, P.A.; Van Oudenaarden, A. MicroRNAs can generate thresholds in target gene expression. Nat. Genet. 2011, 43, 854–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hergenreider, E.; Heydt, S.; Treguer, K.; Boettger, T.; Horrevoets, A.J.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nature 2012, 14, 249–256. [Google Scholar] [CrossRef]
- Schober, A.; Nazari-Jahantigh, M.; Wei, Y.; Bidzhekov, K.; Gremse, F.; Grommes, J.; Megens, R.T.A.; Heyll, K.; Noels, H.; Hristov, M.; et al. MicroRNA-126-5p promotes endothelial proliferation and limits atherosclerosis by suppressing Dlk1. Nat. Med. 2014, 20, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Fleming, I. One miR Level of Control. Hypertension 2012, 60, 1381–1382. [Google Scholar] [CrossRef] [Green Version]
- Kriegel, A.J.; Baker, M.A.; Liu, Y.; Liu, P.; Cowley, A.W.; Liang, M. Endogenous microRNAs in human microvascular endothelial cells regulate mRNAs encoded by hypertension-related genes. Hypertension 2015, 66, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Jusic, A.; Devaux, Y. (CA17129) on behalf of the E.-C.C.A. Noncoding RNAs in Hypertension. Hypertension 2019, 74, 477–492. [Google Scholar] [CrossRef]
- Evans, M.K.; Lepkowski, J.M.; Powe, N.R.; LaVeist, T.; Kuczmarski, M.F.; Zonderman, A. Healthy Aging in Neighborhoods of Diversity across the Life Span (HANDLS): Overcoming barriers to implementing a longitudinal, epidemiologic, urban study of health, race, and socioeconomic status. Ethn. Dis. 2010, 20, 267–275. [Google Scholar]
- Cheadle, C.; Vawter, M.P.; Freed, W.J.; Becker, K.G. Analysis of Microarray Data Using Z Score Transformation. J. Mol. Diagn. 2003, 5, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.S.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, B. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Isogai, T.; van der Kammen, R.; Leyton-Puig, D.; Kedziora, K.M.; Jalink, K.; Innocenti, M. Initiation of Lamellipodia and Ruffles Involves Cooperation between Mdia1 and the Arp2/3 Complex. J. Cell Sci. 2015, 128, 3796–3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaks, M.; Döring, H.; Kage, F.; Steffen, A.; Klünemann, T.; Blankenfeldt, W.; Stradal, T.E.B.; Rottner, K. RhoG and Cdc42 can contribute to Rac-dependent lamellipodia formation through WAVE regulatory complex-binding. Small GTPases 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Steffen, A.; Ladwein, M.; Dimchev, G.; Hein, A.; Schwenkmezger, L.; Arens, S.; Ladwein, K.I.; Holleboom, J.M.; Schur, F.; Small, J.V.; et al. Rac function is crucial for cell migration but is not required for spreading and focal adhesion formation. J. Cell Sci. 2013, 126, 4572–4588. [Google Scholar] [CrossRef] [Green Version]
- Beli, P.; Mascheroni, D.; Xu, D.; Innocenti, M. Wave and Arp2/3 Jointly Inhibit Filopodium Formation by Entering into a Complex with Mdia2. Nat. Cell Biol. 2008, 10, 849–857. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef]
- Davies, P.F. Hemodynamic shear stress and the endothelium in cardiovascular pathophysiology. Nat. Clin. Pract. Neurol. 2008, 6, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Iskratsch, T.; Wolfenson, H.; Sheetz, M.P. Appreciating force and shape—the rise of mechanotransduction in cell biology. Nat. Rev. Mol. Cell Boil. 2014, 15, 825–833. [Google Scholar] [CrossRef]
- Spindler, V.; Schlegel, N.; Waschke, J. Role of GTPases in control of microvascular permeability. Cardiovasc. Res. 2010, 87, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, M.; Gautreau, A.M. Steering cell migration: Lamellipodium dynamics and the regulation of directional persistence. Nat. Rev. Mol. Cell Boil. 2014, 15, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J. Life at the Leading Edge. Cell 2011, 145, 1012–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, J.-V.; Stradal, T.E.B.; Vignal, E.; Rottner, K. The lamellipodium: Where motility begins. Trends Cell Boil. 2002, 12, 112–120. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, X.; Pan, L.; Tian, X.; Dong, P. Identification of Key Pathways and Genes in Advanced Coronary Atherosclerosis Using Bioinformatics Analysis. BioMed Res. Int. 2017, 2017, 4323496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandaru, S.; Ala, C.; Salimi, R.; Akula, M.K.; Ekstrand, M.; Devarakonda, S.; Karlsson, J.; Eynden, J.V.D.; Bergström, G.; Larsson, E.; et al. Targeting Filamin A Reduces Macrophage Activity and Atherosclerosis. Circulation 2019, 140, 67–79. [Google Scholar] [CrossRef]
- Bandaru, S.; Grönros, J.; Redfors, B.; Pazooki, D.; Salimi, R.; Larsson, E.; Zhou, A.-X.; Omerovic, E.; Çil, Ç.; Akyürek, L.M. Deficiency of filamin A in endothelial cells impairs left ventricular remodelling after myocardial infarction. Cardiovasc. Res. 2014, 105, 151–159. [Google Scholar] [CrossRef] [Green Version]
- E Marketou, M.; Kontaraki, J.; Zacharis, E.; Parthenakis, F.; Maragkoudakis, S.; Gavras, I.; Gavras, H.; E Vardas, P. Differential gene expression of bradykinin receptors 1 and 2 in peripheral monocytes from patients with essential hypertension. J. Hum. Hypertens. 2014, 28, 450–455. [Google Scholar] [CrossRef]
- Byrne, K.M.; Monsefi, N.; Dawson, J.C.; Degasperi, A.; Wills, J.; Volinsky, N.; Dobrzynski, M.; Birtwistle, M.R.; Tsyganov, M.A.; Kiyatkin, A.; et al. Bistability in the Rac1, PAK, and RhoA Signaling Network Drives Actin Cytoskeleton Dynamics and Cell Motility Switches. Cell Syst. 2016, 2, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Carbone, M.L.; Chadeuf, G.; Heurtebise-Chrétien, S.; Prieur, X.; Quillard, T.; Goueffic, Y.; Vaillant, N.; Rio, M.; Castan, L.; Durand, M.; et al. Leukocyte RhoA exchange factor Arhgef1 mediates vascular inflammation and atherosclerosis. J. Clin. Investig. 2017, 127, 4516–4526. [Google Scholar] [CrossRef] [Green Version]
- Narumiya, S.; Thumkeo, D. Rho signaling research: History, current status and future directions. FEBS Lett. 2018, 592, 1763–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dee, R.A.; Mangum, K.D.; Bai, X.; Mack, C.; Taylor, J. Druggable targets in the Rho pathway and their promise for therapeutic control of blood pressure. Pharmacol. Ther. 2019, 193, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Wang, J.; Longley, S.J.; Tang, J.X.; Shaw, S.K. Relative actin nucleation promotion efficiency by WASP and WAVE proteins in endothelial cells. Biochem. Biophys. Res. Commun. 2010, 400, 661–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotty, J.D.; Wu, C.; Bear, J.E. New Insights into the Regulation and Cellular Functions of the Arp2/3 Complex. Nat. Rev. Mol. Cell Biol. 2013, 14, 7–12. [Google Scholar] [CrossRef]
- Innocenti, M.; Gerboth, S.; Rottner, K.; Lai, F.P.L.; Hertzog, M.; Stradal, T.E.B.; Frittoli, E.; Didry, M.; Polo, S.; Disanza, A.; et al. Abi1 regulates the activity of N-WASP and WAVE in distinct actin-based processes. Nature 2005, 7, 969–976. [Google Scholar] [CrossRef]
- Suarez, C.; Kovar, D.R. Internetwork competition for monomers governs actin cytoskeleton organization. Nat. Rev. Mol. Cell Boil. 2016, 17, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Babbitt, D.M.; Kim, J.-S.; Forrester, S.J.; Brown, M.D.; Park, J.-Y. Effect of Interleukin-10 and Laminar Shear Stress on Endothelial Nitric Oxide Synthase and Nitric Oxide in African American Human Umbilical Vein Endothelial Cells. Ethn. Dis. 2015, 25, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Feairheller, D.L.; Park, J.-Y.; Rizzo, V.; Kim, B.; Brown, M. Racial differences in the responses to shear stress in human umbilical vein endothelial cells. Vasc. Health Risk Manag. 2011, 7, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Buie, J.N.J.; Stanley, A.; Nietert, P.J.; Logan, A.; Adams, R.J.; Magwood, G.S. Racial Disparities in Arterial Stiffness Between Healthy Whites and African Americans in the United States: A Meta-analysis. J. Natl. Med. Assoc. 2019, 111, 7–17. [Google Scholar] [CrossRef]
- Morris, A.A.; Patel, R.S.; Binongo, J.N.G.; Poole, J.; Al Mheid, I.; Ahmed, Y.; Stoyanova, N.; Vaccarino, V.; Din-Dzietham, R.; Gibbons, G.H.; et al. Racial Differences in Arterial Stiffness and Microcirculatory Function Between Black and White Americans. J. Am. Heart Assoc. 2013, 2, e002154. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.T.; Sobey, C.G.; Lieu, M.; Ferens, D.; Kett, M.M.; Diep, H.; Kim, H.A.; Krishnan, S.M.; Lewis, C.; Salimova, E.; et al. Obligatory Role for B Cells in the Development of Angiotensin II–Dependent Hypertension. Hypertension 2015, 66, 1023–1033. [Google Scholar] [CrossRef]
- Caillon, A.; Mian, M.O.R.; Fraulob-Aquino, J.C.; Huo, K.; Barhoumi, T.; Paradis, P.; Schiffrin, E.L. Gammadelta T Cells Mediate Angiotensin Ii-Induced Hypertension and Vascular Injury. J. Hypertens. 2016, 34, e290. [Google Scholar] [CrossRef]
- Ni, X.; Wang, A.; Zhang, L.; Shan, L.-Y.; Zhang, H.-C.; Li, L.; Si, J.-Q.; Luo, J.; Li, X.-Z.; Ma, K.-T. Up-regulation of gap junction in peripheral blood T lymphocytes contributes to the inflammatory response in essential hypertension. PLoS ONE 2017, 12, e0184773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, G.R.; Vinh, A.; Guzik, T.J.; Sobey, C.G. Immune mechanisms of hypertension. Nat. Rev. Immunol. 2019, 19, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, P.; Knorr, M.; Kossmann, S.; Stratmann, J.; Hausding, M.; Schuhmacher, S.; Karbach, S.H.; Schwenk, M.; Yogev, N.; Schulz, E.; et al. Lysozyme M-Positive Monocytes Mediate Angiotensin II-Induced Arterial Hypertension and Vascular Dysfunction. Circulation 2011, 124, 1370–1381. [Google Scholar] [CrossRef] [Green Version]
- Heasman, S.J.; Ridley, A.J. Multiple roles for RhoA during T cell transendothelial migration. Small GTPases 2010, 1, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Ricker, E.; Chowdhury, L.; Yi, W.; Pernis, A. The RhoA-ROCK pathway in the regulation of T and B cell responses. F1000Research 2016, 5, 2295. [Google Scholar] [CrossRef] [Green Version]
- Königs, V.; Jennings, R.; Vogl, T.; Horsthemke, M.; Bachg, A.C.; Xu, Y.; Grobe, K.; Brakebusch, C.; Schwab, A.; Bähler, M.; et al. Mouse Macrophages Completely Lacking Rho Subfamily GTPases (RhoA, RhoB, and RhoC) Have Severe Lamellipodial Retraction Defects, but Robust Chemotactic Navigation and Altered Motility. J. Boil. Chem. 2014, 289, 30772–30784. [Google Scholar] [CrossRef] [Green Version]
- Hindle, A.G.; Thoonen, R.; Jasien, J.V.; Grange, R.M.H.; Amin, K.; Wise, J.; Ozaki, M.; Ritch, R.; Malhotra, R.; Buys, E.S. Identification of Candidate miRNA Biomarkers for Glaucoma. Investig. Opthalmol. Vis. Sci. 2019, 60, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Kanchan, R.K.; Perumal, N.; Atri, P.; Venkata, R.C.; Thapa, I.; Klinkebiel, D.L.; Donson, A.M.; Perry, D.; Punsoni, M.; Talmon, G.A.; et al. Mir-1253 Exerts Tumor-Suppressive Effects in Medulloblastoma Via Inhibition of Cdk6 and Cd276 (B7-H3). Brain Pathol. 2020. [CrossRef]
- Chen, Y.; Gu, M.; Liu, C.; Wan, X.; Shi, Q.; Chen, Q.; Wang, Z. Long Noncoding Rna Foxc2-As1 Facilitates the Proliferation and Progression of Prostate Cancer Via Targeting Mir-1253/Ezh2. Gene 2019, 686, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Y.; Zhang, J.; Cai, H.; Zhang, C.; Yang, Z.; Niu, Y.; Wang, H.; Wei, X.; Wang, W.; et al. MicroRNA-1253 suppresses cell proliferation and invasion of non-small-cell lung carcinoma by targeting WNT5A. Cell Death Dis. 2018, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yao, Y.; Gao, P.; Cui, Y. Upregulated circular RNA circ_0030235 predicts unfavorable prognosis in pancreatic ductal adenocarcinoma and facilitates cell progression by sponging miR-1253 and miR-1294. Biochem. Biophys. Res. Commun. 2018, 509, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Xu, Z.M.; Wang, X.L.; Zheng, J.K.; Du, Q.; Yang, J.X.; Zhang, H.C. Lncrna Foxc2-As1 Regulated Proliferation and Apoptosis of Vascular Smooth Muscle Cell through Targeting Mir-1253/Foxf1 Axis in Atherosclerosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3302–3314. [Google Scholar] [PubMed]
- Shang, L.; Smith, J.A.; Zhao, W.; Kho, M.; Turner, S.T.; Mosley, T.H.; Kardia, S.L.; Zhou, X. Genetic Architecture of Gene Expression in European and African Americans: An eQTL Mapping Study in GENOA. Am. J. Hum. Genet. 2020, 106, 496–512. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-X.; Zeng, D.-Y.; Li, R.-T.; Pang, R.-P.; Yang, H.; Hu, Y.-L.; Zhang, Q.; Jiang, Y.; Huang, L.-Y.; Tang, Y.-B.; et al. Essential Role of MicroRNA-155 in Regulating Endothelium-Dependent Vasorelaxation by Targeting Endothelial Nitric Oxide Synthase. Hypertension 2012, 60, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Ceolotto, G.; Papparella, I.; Bortoluzzi, A.; Strapazzon, G.; Ragazzo, F.; Bratti, P.; Fabricio, A.S.C.; Squarcina, E.; Gion, M.; Palatini, P.; et al. Interplay between miR-155, AT1R A1166C polymorphism, and AT1R expression in young untreated hypertensives. Am. J. Hypertens. 2011, 24, 241–246. [Google Scholar] [CrossRef]
- E Kontaraki, J.; E Marketou, M.; A Zacharis, E.; I Parthenakis, F.; E Vardas, P. Differential expression of vascular smooth muscle-modulating microRNAs in human peripheral blood mononuclear cells: Novel targets in essential hypertension. J. Hum. Hypertens. 2013, 28, 510–516. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted miR-1253 Targets (Compared w/Scrambled Control) | FDR | Fold Change | p-Value | Z-Ratio |
|---|---|---|---|---|
| ABCB10 | 0.0172 | −1.53 | 0.0032 | −2.83 |
| ACO1 | 0 | −7.58 | 0 | −11.94 |
| ACSL1 | 0 | −1.69 | 0 | −3.32 |
| DCUN1D5 | 0.0116 | −1.87 | 0.002 | −3.8 |
| DPYSL2 | 0 | −1.61 | 0 | −2.47 |
| DUSP14 | 0 | −2.13 | 0 | −4.46 |
| MSN | 0 | −1.83 | 0 | −3.15 |
| PARP1 | 0 | −4.4 | 0 | −8.57 |
| PDE12 | 0 | −2 | 0 | −4.36 |
| POLA1 | 0 | −1.53 | 0 | −2.82 |
| PTGER4 | 0 | −1.72 | 0 | −3.05 |
| RAB27A | 0 | −2.06 | 0 | −4.5 |
| RSU1 | 0 | −2.16 | 0 | −4.78 |
| RXRA | 0 | −1.54 | 0 | −2.61 |
| SEC62 | 0 | −1.55 | 0 | −2.96 |
| SERINC3 | 0 | −2.51 | 0 | −5.53 |
| SPARC | 0 | −1.91 | 0 | −3.55 |
| TFRC | 0 | −1.71 | 0 | −3.1 |
| TMEM127 | 0 | −1.89 | 0 | −4.07 |
| TNS3 | 0 | −1.83 | 0 | −3.68 |
| TOPBP1 | 0 | −2.09 | 0 | −4.35 |
| UBE2N | 0.0005 | −1.56 | 0.0001 | −2.67 |
| WASF2 | 0 | −1.64 | 0 | −3.09 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arkorful, M.A.; Noren Hooten, N.; Zhang, Y.; Hewitt, A.N.; Barrientos Sanchez, L.; Evans, M.K.; Dluzen, D.F. MicroRNA-1253 Regulation of WASF2 (WAVE2) and its Relevance to Racial Health Disparities. Genes 2020, 11, 572. https://doi.org/10.3390/genes11050572
Arkorful MA, Noren Hooten N, Zhang Y, Hewitt AN, Barrientos Sanchez L, Evans MK, Dluzen DF. MicroRNA-1253 Regulation of WASF2 (WAVE2) and its Relevance to Racial Health Disparities. Genes. 2020; 11(5):572. https://doi.org/10.3390/genes11050572
Chicago/Turabian StyleArkorful, Mercy A., Nicole Noren Hooten, Yongqing Zhang, Amirah N. Hewitt, Lori Barrientos Sanchez, Michele K. Evans, and Douglas F. Dluzen. 2020. "MicroRNA-1253 Regulation of WASF2 (WAVE2) and its Relevance to Racial Health Disparities" Genes 11, no. 5: 572. https://doi.org/10.3390/genes11050572