Crotoxin-Induced Mice Lung Impairment: Role of Nicotinic Acetylcholine Receptors and COX-Derived Prostanoids

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
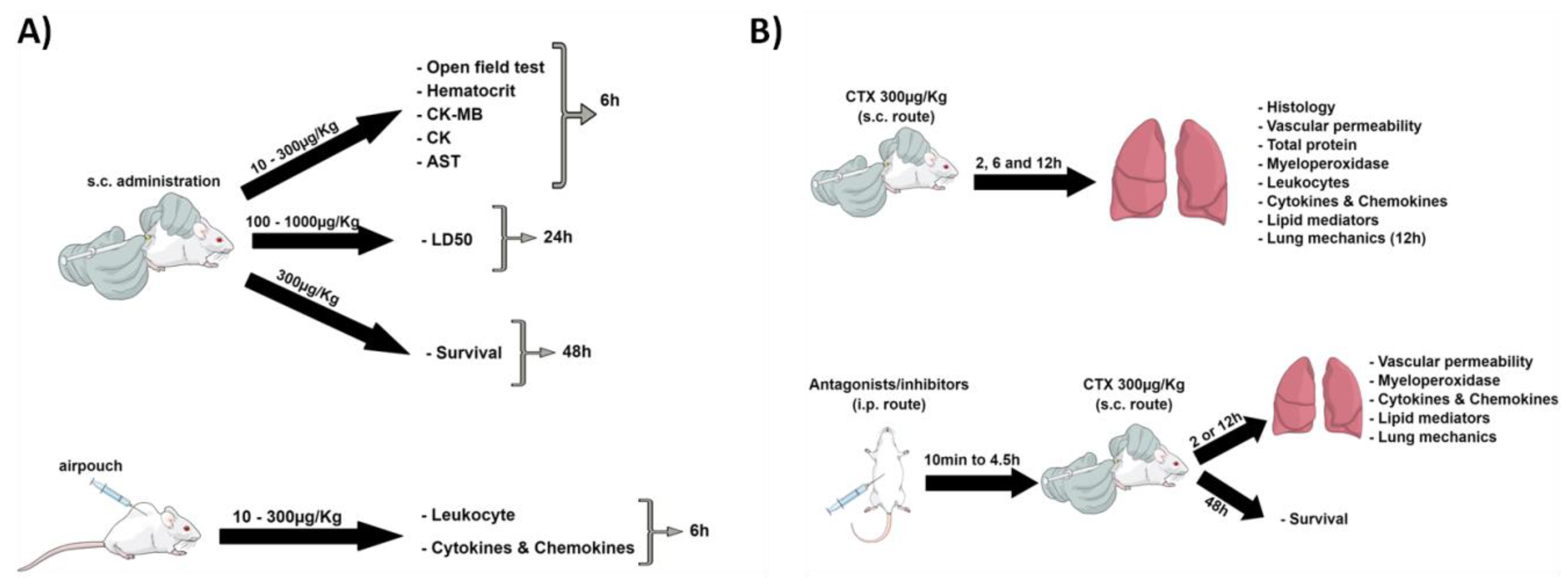
2. Materials and Methods
2.1. Animals
2.2. Crotoxin
2.3. CTX and Drug Treatments
2.3.1. CTX Dose- and Time-Response Experiments
2.3.2. Drug Treatments
2.4. In Vivo Experiments
2.4.1. Lethality
2.4.2. Open Field Test
2.4.3. Vascular Permeability
2.4.4. Air Pouch Model to Examine Local Inflammation
2.4.5. Lung Mechanics
2.5. Biological Parameters and Markers
2.5.1. Biochemical Markers
2.5.2. Pro-Inflammatory Cytokines
2.5.3. Hematocrit
2.5.4. Gene Expression
2.5.5. Eicosanoids
2.5.6. Myeloperoxidase Activity
2.6. Lung Histology
2.7. Analysis of Lung Leukocyte Population
2.8. Statistical Analysis
3. Results
3.1. CTX Working Dose
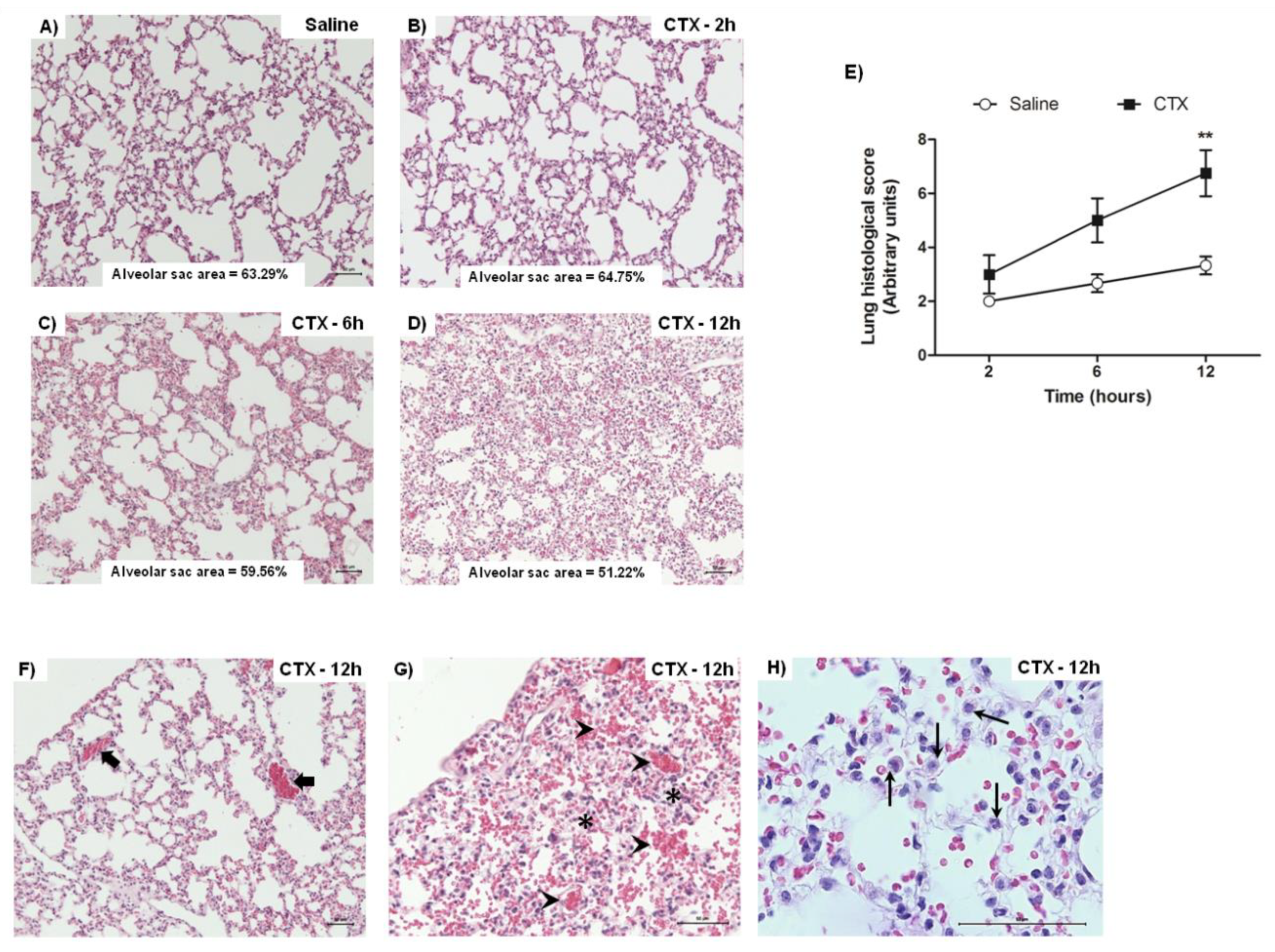
3.2. CTX Induces Lung Alterations
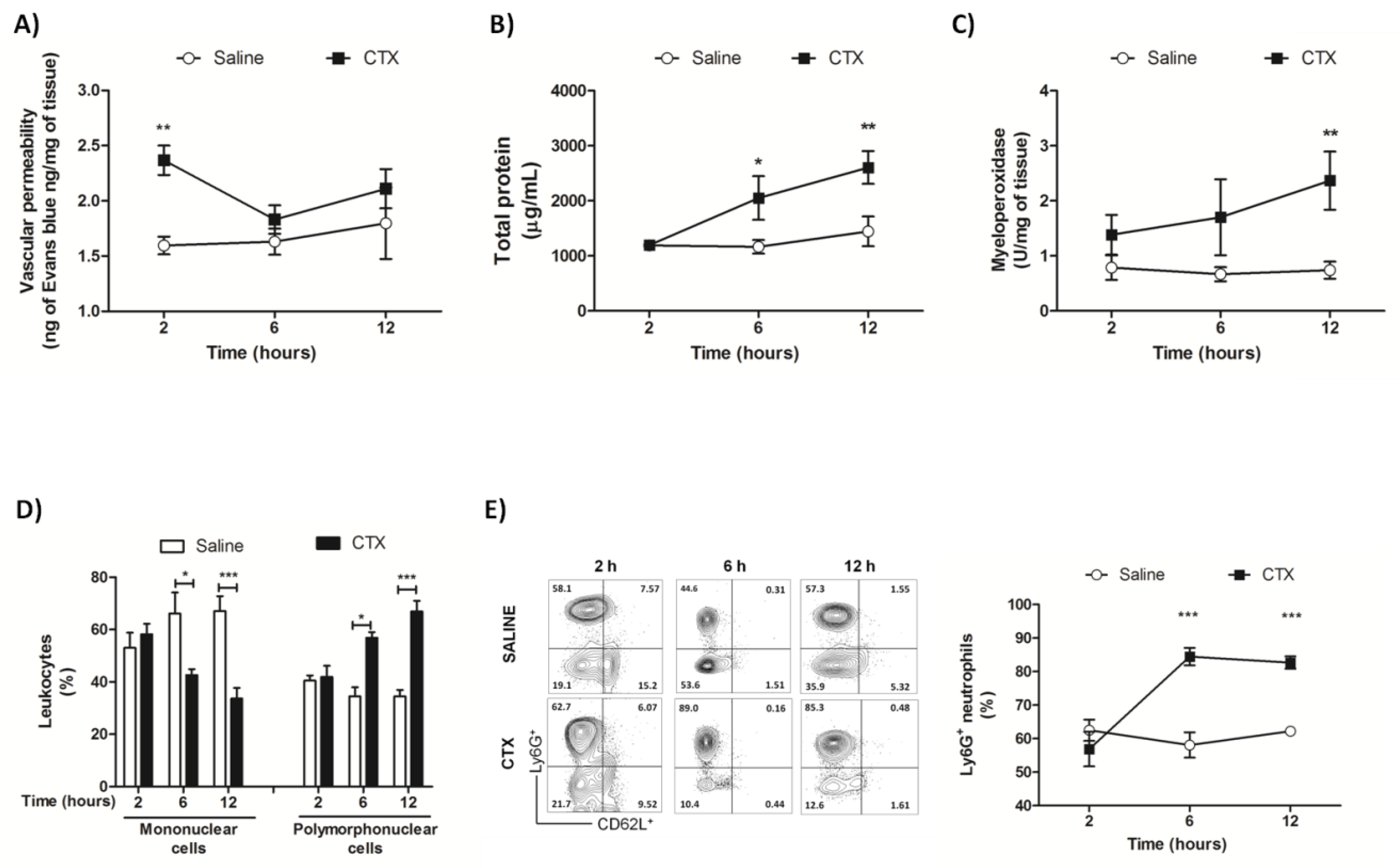
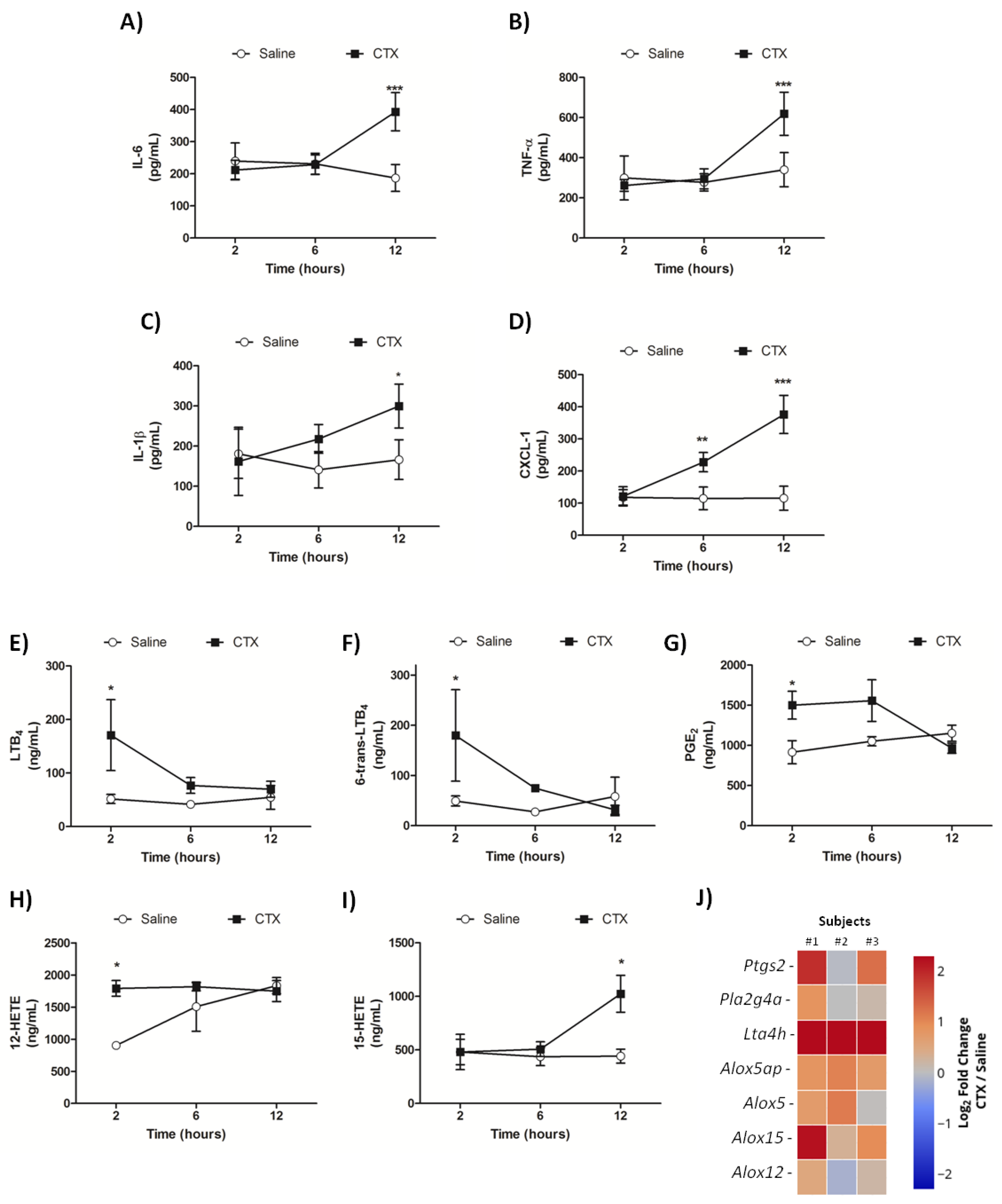
3.3. CTX Elevates the Levels of Inflammatory Mediators and Impairs Pulmonary Function
3.4. Indomethacin and Hexamethonium Mitigate CTX-Induced Lethality
3.5. Role of Hexamethonium And Indomethacin on CTX-Induced Lung Alterations
3.5.1. Early Phase
3.5.2. Late Phase
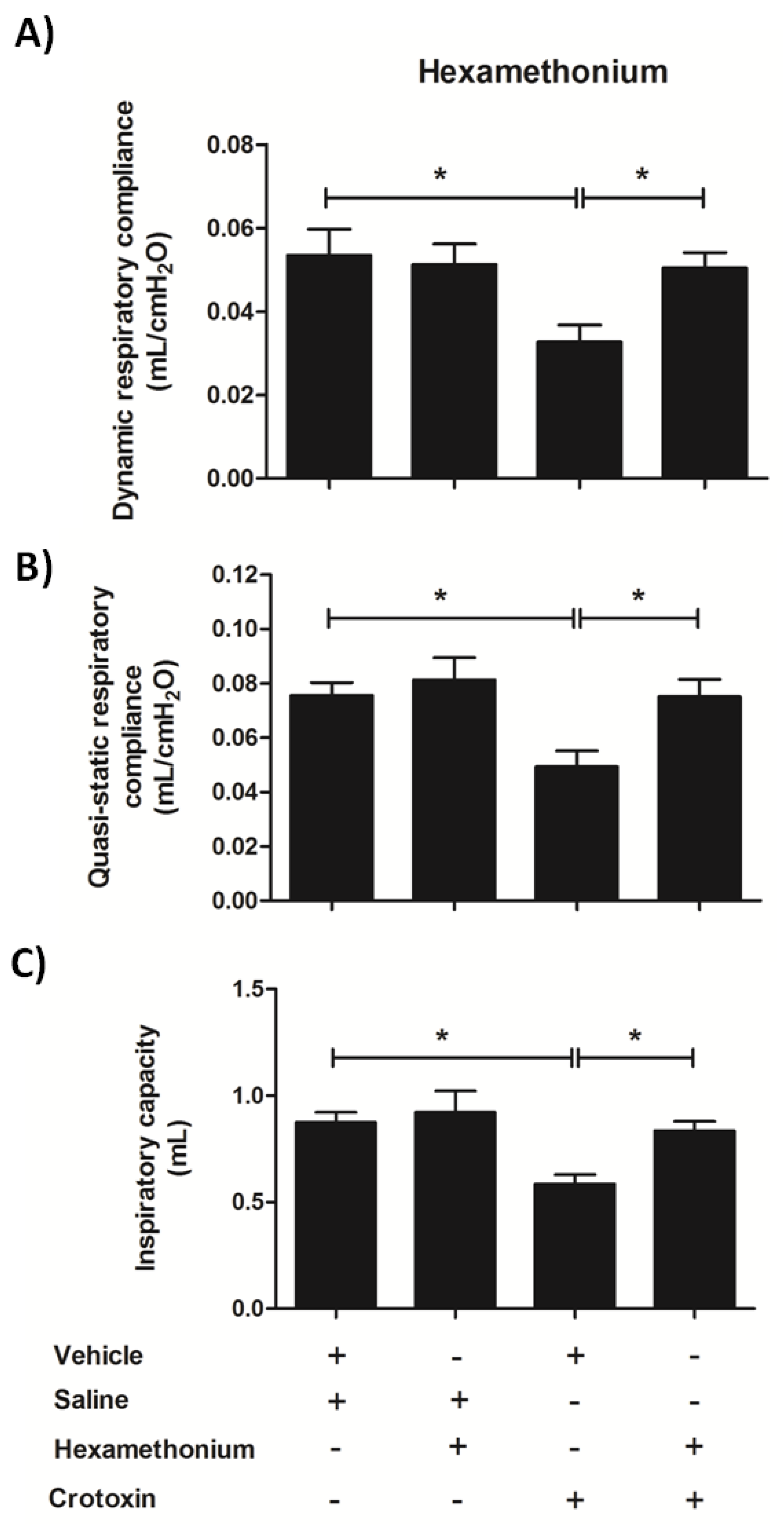
3.6. Hexamethonium Mitigates CTX-Induced Lung Mechanics Impairment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Muller, V.D.M.; Russo, R.R.; Oliveira Cintra, A.C.; Sartim, M.A.; De Melo Alves-Paiva, R.; Figueiredo, L.T.M.; Sampaio, S.V.; Aquino, V.H. Crotoxin and phospholipases A 2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon 2012, 59, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Zoccal, K.F.; Sorgi, C.A.; Hori, J.I.; Paula-Silva, F.W.G.; Arantes, E.C.; Serezani, C.H.; Zamboni, D.S.; Faccioli, L.H. Opposing roles of LTB4 and PGE2 in regulating the inflammasome-dependent scorpion venom-induced mortality. Nat. Commun. 2016, 7, 10760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, N.G.; Sampaio, S.C.; Gonçalves, L.R.C. Inhibitory effect of Crotalus durissus terrificus venom on chronic edema induced by injection of bacillus Calmette-Guérin into the footpad of mice. Toxicon 2013, 63, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Giannopoulos, P.F.; Chu, J.; Joshi, Y.B.; Sperow, M.; Li, J.G.; Kirby, L.G.; Praticò, D. 5-lipoxygenase activating protein reduction ameliorates cognitive deficit, synaptic dysfunction, and neuropathology in a mouse model of Alzheimer’s disease. Biol. Psychiatry 2013, 74, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaisakul, J.; Ahmad Rusmili, M.R.; Hodgson, W.C.; Hatthachote, P.; Suwan, K.; Inchan, A.; Chanhome, L.; Othman, I.; Chootip, K. A pharmacological examination of the cardiovascular effects of Malayan krait (Bungarus candidus) venoms. Toxins 2017, 9, E122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa Nogueira-Neto, F.; Amorim, R.L.; Brigatte, P.; Picolo, G.; Ferreira, W.A.; Gutierrez, V.P.; Conceição, I.M.; Della-Casa, M.S.; Takahira, R.K.; Nicoletti, J.L.M.; et al. The analgesic effect of crotoxin on neuropathic pain is mediated by central muscarinic receptors and 5-lipoxygenase-derived mediators. Pharmacol. Biochem. Behav. 2008, 91, 252–260. [Google Scholar] [CrossRef]
- Lee, C.Y.; Chen, Y.M.; Joubert, F.J. Protection by atropine against synergistic lethal effects of the Angusticeps-type toxin F7 from eastern green mamba venom and toxin I from black mamba venom. Toxicon 1982, 20, 665–667. [Google Scholar] [CrossRef]
- Durand, M.T.; Becari, C.; Tezini, G.C.S.V.; Fazan, R.; Oliveira, M.; Guatimosim, S.; Prado, V.F.; Prado, M.A.M.; Salgado, H.C. Autonomic cardiocirculatory control in mice with reduced expression of the vesicular acetylcholine transporter. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H655–H662. [Google Scholar] [CrossRef] [Green Version]
- Zanelati, T.V.; Biojone, C.; Moreira, F.A.; Guimarães, F.S.; Joca, S.R.L. Antidepressant-like effects of cannabidiol in mice: Possible involvement of 5-HT 1A receptors. Br. J. Pharmacol. 2010, 159, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Radu, M.; Chernoff, J. An in vivo assay to test blood vessel permeability. J. Vis. Exp. 2013, 73, e50062. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Sorgi, C.A.; Peti, A.P.F.; Petta, T.; Meirelles, A.F.G.; Fontanari, C.; Moraes, L.A.B.; Faccioli, L.H. Comprehensive high-resolution multiple-reaction monitoring mass spectrometry for targeted eicosanoid assays. Sci. Data 2018, 21, 180167. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.R.; Provinciatto, P.R.; Godoi, D.F.; Ferreira, B.R.; Teixeira, G.; Rossi, M.A.; Cunha, F.Q.; Silva, J.S. IL-4 regulates susceptibility to intestinal inflammation in murine food allergy. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G593–G600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, C.O.S.; Espíndola, M.S.; Fontanari, C.; Prado, M.K.B.; Frantz, F.G.; Rodrigues, V.; Gardinassi, L.G.; Faccioli, L.H. CD18 regulates monocyte hematopoiesis and promotes resistance to experimental schistosomiasis. Front. Immunol. 2018, 9, 1970. [Google Scholar] [CrossRef]
- Diz Filho, E.B.S.; Marangoni, S.; Toyama, D.O.; Fagundes, F.H.R.; Oliveira, S.C.B.; Fonseca, F.V.; Calgarotto, A.K.; Joazeiro, P.P.; Toyama, M.H. Enzymatic and structural characterization of new PLA2 isoform isolated from white venom of Crotalus durissus ruruima. Toxicon 2009, 53, 104–114. [Google Scholar] [CrossRef]
- Cavalcante, W.L.G.; Noronha-Matos, J.B.; Timóteo, M.A.; Fontes, M.R.M.; Gallacci, M.; Correia-de-Sá, P. Neuromuscular paralysis by the basic phospholipase A2 subunit of crotoxin from Crotalus durissus terrificus snake venom needs its acid chaperone to concurrently inhibit acetylcholine release and produce muscle blockage. Toxicol. Appl. Pharmacol. 2017, 334, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Araújo, D.A.; Beirão, P.S. Effects of crotoxin on the action potential kinetics of frog skeletal muscle. Braz. J. Med. Biol. Res. 1993, 26, 1111–1121. [Google Scholar]
- Cura, J.E.; Blanzaco, D.P.; Brisson, C.; Cura, M.A.; Cabrol, R.; Larrateguy, L.; Mendez, C.; Sechi, J.C.; Silveira, J.S.; Theiller, E.; et al. Phase I and pharmacokinetics study of crotoxin (cytotoxic PLA2, NSC-624244) in patients with advanced cancer. Clin. Cancer Res. 2002, 8, 1033–1041. [Google Scholar]
- Câmara, P.R.S.; Esquisatto, L.C.M.; Camargo, E.A.; Ribela, M.T.C.P.; Toyama, M.H.; Marangoni, S.; De Nucci, G.; Antunes, E. Inflammatory oedema induced by phospholipases A2 isolated from Crotalus durissus sp. in the rat dorsal skin: A role for mast cells and sensory C-fibers. Toxicon 2003, 41, 823–829. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Alberto Ponce-Soto, L.; Marangoni, S.; Lomonte, B. Systemic and local myotoxicity induced by snake venom group II phospholipases A2: Comparison between crotoxin, crotoxin B and a Lys49 PLA2 homologue. Toxicon 2008, 51, 80–92. [Google Scholar] [CrossRef]
- Moreira, V.; Gutiérrez, J.M.; Soares, A.M.; Zamunér, S.R.; Purgatto, E.; de Fátima Pereira Teixeira, C. Secretory phospholipases A2 isolated from Bothrops asper and from Crotalus durissus terrificus snake venoms induce distinct mechanisms for biosynthesis of prostaglandins E2 and D2 and expression of cyclooxygenases. Toxicon 2008, 52, 428–459. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Ong, W.Y.; Thwin, M.M.; Fong, C.W.; Farooqui, A.A.; Gopalakrishnakone, P.; Hong, W. Group IIA secretory phospholipase A2 stimulates exocytosis and neurotransmitter release in pheochromocytoma-12 cells and cultured rat hippocampal neurons. Neuroscience 2003, 121, 891–898. [Google Scholar] [CrossRef]
- Xavier, C.V.; da S. Setúbal, S.; Lacouth-Silva, F.; Pontes, A.S.; Nery, N.M.; de Castro, O.B.; Fernandes, C.F.C.; Soares, A.M.; Fortes-Dias, C.L.; Zuliani, J.P. Phospholipase A2 Inhibitor from Crotalus durissus terrificus rattlesnake: Effects on human peripheral blood mononuclear cells and human neutrophils cells. Int. J. Biol. Macromol. 2017, 105, 1117–1125. [Google Scholar] [PubMed]
- Azevedo-Marques, M.M.; Cupo, P.; Coimbra, T.M.; Hering, S.E.; Rossi, M.A.; Laure, C.J. Myonecrosis, myoglobinuria and acute renal failure induced by south american rattlesnake (Crotalus durissus terrificus) envenomation in brazil. Toxicon 1985, 23, 631–636. [Google Scholar] [CrossRef]
- Azevedo-Marques, M.M.; Cupo, P.; Amaral, C.F.S.; Hering, S.E. Rattlesnake bites. Clinical features and complementary tests. Mem. Inst. Butantan 1990, 52, 27–30. [Google Scholar]
- Bucaretchi, F.; De Capitani, E.M.; Branco, M.M.; Fernandes, L.C.R.; Hyslop, S. Coagulopathy as the main systemic manifestation after envenoming by a juvenile South American rattlesnake (Crotalus durissus terrificus): Case report. Clin. Toxicol. 2013, 51, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Hardy, D.L. Envenomation by the Mojave rattlesnake (Crotalus scutulatus scutulatus) in southern Arizona, USA. Toxicon 1983, 21, 11–118. [Google Scholar] [CrossRef]
- Amaral, C.F.; Magalhães, R.A.; de Rezende, N.A. Respiratory involvement secondary to crotalid ophidian bite (Crotalus durissus). Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Kerns, W.; Tomaszewski, C. Airway obstruction following canebrake rattlesnake envenomation. J. Emerg. Med. 2001, 20, 377–380. [Google Scholar] [CrossRef]
- Brooks, D.E.; Graeme, K.A.; Ruha, A.M.; Tanen, D.A. Respiratory compromise in patients with rattlesnake envenomation. J. Emerg. Med. 2002, 23, 329–332. [Google Scholar] [CrossRef]
- Baum, R.A.; Bronner, J.; Akpunonu, P.D.S.; Plott, J.; Bailey, A.M.; Keyler, D.E. Crotalus durissus terrificus (viperidae; crotalinae) envenomation: Respiratory failure and treatment with antivipmyn TRI ® antivenom. Toxicon 2019, 163, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.G.; Nascimento, N.; Rogero, J.R.; Vassilieff, V.S. Gabaergic-benzodiazepine system is involved in the crotoxin-induced anxiogenic effect. Pharmacol. Biochem. Behav. 2000, 65, 7–13. [Google Scholar] [CrossRef]
- De Barros Ribeiro, G.; De Almeida, H.C.; Velarde, D.T.; De Melo Sá, M.L.V. Study of crotoxin on the induction of paralysis in extraocular muscle in animal model. Arq. Bras. Oftalmol. 2012, 75, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Santos, A.; Dos-Santos, E.C.; Lopes-Ferreira, M.; Lima, C.; Cardoso, D.F.; Mota, I. A comparative study of biological activities of crotoxin and CB fraction of venoms from Crotalus durissus terrificus, Crotalus durissus cascavella and Crotalus durissus collilineatus. Toxicon 2004, 43, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Brazil, O.V.; Fariña, R.; Yoshida, L.; De Oliveira, V.A. Pharmacology of crystalline crotoxin. 3. Cardiovascular and respiratory effects of crotoxin and Crotalus durissus terrificus venom. Mem. Inst. Butantan 1966, 33, 1000. [Google Scholar]
- Cupo, P.; Azevedo-Marques, M.M.; Hering, S.E. Acute myocardial infarction-like enzyme profile in human victims of Crotalus durissus terrificus envenoming. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 447–451. [Google Scholar] [CrossRef]
- Sano-Martins, I.S. Coagulopathy following lethal and non-lethal envenoming of humans by the South American rattlesnake (Crotalus durissus) in Brazil. QJM 2001, 94, 551–559. [Google Scholar] [CrossRef] [Green Version]
- de Sousa-e-Silva, M.C.C.; Tomy, S.C.; Tavares, F.L.; Navajas, L.; Larsson, M.H.M.A.; Lucas, S.R.R.; Kogika, M.M.; Sano-Martins, I.S. Hematological, hemostatic and clinical chemistry disturbances induced by Crotalus durissus terrificus snake venom in dogs. Hum. Exp. Toxicol. 2003, 22, 491–500. [Google Scholar] [CrossRef]
- Brazil, O.V. Pharmacology of crystalline crotoxin. II. Neuromuscular blocking action. Mem. Inst. Butantan 1966, 33, 981–992. [Google Scholar]
- Haschek, W.M.; Rousseaux, C.G.; Wallig, M.A.; Bolon, B.; Ochoa, R. Haschek and Rousseaux’s Handbook of Toxicologic Pathology, 3rd ed.; Elsevier: Waltham, MA, USA, 2013; ISBN 9780124157590. [Google Scholar]
- Sanderson, M.J. Exploring lung physiology in health and disease with lung slices. Pulm. Pharmacol. Ther. 2011, 24, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, P.N.; Amorim, C.F.; Paneque Peres, A.C.; e Silva, C.A.M.; Zamuner, S.R.; Ribeiro, W.; Cogo, J.C.; Vieira, R.P.; Dolhnikoff, M.; de Oliveira, L.V.F. Pulmonary mechanic and lung histology injury induced by Crotalus durissus terrificus snake venom. Toxicon 2008, 51, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Neto, J.; de Moraes Silveira, J.A.; Serra, D.S.; de Araújo Viana, D.; Borges-Nojosa, D.M.; Sampaio, C.M.S.; Monteiro, H.S.A.; Cavalcante, F.S.Á.; Evangelista, J.S.A.M. Pulmonary mechanic and lung histology induced by Crotalus durissus cascavella snake venom. Toxicon 2017, 137, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.C. Acute lung injury and pulmonary vascular permeability: Use of transgenic models. Compr. Physiol. 2011, 1, 835–882. [Google Scholar] [PubMed]
- Mammoto, A.; Mammoto, T.; Kanapathipillai, M.; Yung, C.W.; Jiang, E.; Jiang, A.; Lofgren, K.; Gee, E.P.S.; Ingber, D.E. Control of lung vascular permeability and endotoxin-induced pulmonary oedema by changes in extracellular matrix mechanics. Nat. Commun. 2013, 4, 1759. [Google Scholar] [CrossRef] [Green Version]
- Peters-Golden, M.; Brock, T.G. 5-Lipoxygenase and FLAP. Prostaglandins Leukot. Essent. Fat. Acids 2003, 69, 99–109. [Google Scholar] [CrossRef]
- Wan, M.; Tang, X.; Stsiapanava, A.; Haeggström, J.Z. Biosynthesis of leukotriene B4. Semin. Immunol. 2017, 33, 3–15. [Google Scholar] [CrossRef]
- Nakamura, H.; Wakita, S.; Suganami, A.; Tamura, Y.; Hanada, K.; Murayama, T. Modulation of the activity of cytosolic phospholipase A2α (cPLA2α) by cellular sphingolipids and inhibition of cPLA2α by sphingomyelin. J. Lipid Res. 2010, 51, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Fretland, D.J.; Widomski, D.L.; Anglin, C.P.; Gaginella, T.S. The antiinflammatory agent SC-41930 inhibits granulocyte infiltration of the rodent dermis induced by 6-trans-leukotriene B4. Prostaglandins Leukot. Essent. Fat. Acids 1991, 44, 61–65. [Google Scholar] [CrossRef]
- Kalinski, P. Regulation of immune responses by prostaglandin E 2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.S.; Rokach, J. Biosynthesis, biological effects, and receptors of hydroxyeicosatetraenoic acids (HETEs) and oxoeicosatetraenoic acids (oxo-ETEs) derived from arachidonic acid. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 340–355. [Google Scholar] [CrossRef] [Green Version]
- Miyabe, Y.; Miyabe, C.; Luster, A.D. LTB4 and BLT1 in inflammatory arthritis. Semin. Immunol. 2017, 33, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Omori, K.; Kida, T.; Hori, M.; Ozaki, H.; Murata, T. Multiple roles of the PGE2-EP receptor signal in vascular permeability. Br. J. Pharmacol. 2014, 171, 4879–4889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, E.H.; Davies, B.H. Dual effect of prostaglandm E2 on normal airways smooth muscle in vivo. Thorax 1982, 37, 918–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannotti, K.C.; Leiguez, E.; Carvalho, A.E.Z.D.; Nascimento, N.G.; Matsubara, M.H.; Fortes-Dias, C.L.; Moreira, V.; Teixeira, C. A snake venom group IIA PLA2 with immunomodulatory activity induces formation of lipid droplets containing 15-d-PGJ2 in macrophages. Sci. Rep. 2017, 7, 4098. [Google Scholar] [CrossRef]
- Sampaio, S.C.; Alba-Loureiro, T.C.; Brigatte, P.; Landgraf, R.G.; Santos, E.C.D.; Curi, R.; Cury, Y. Lipoxygenase-derived eicosanoids are involved in the inhibitory effect of Crotalus durissus terrificus venom or crotoxin on rat macrophage phagocytosis. Toxicon 2006, 47, 313–321. [Google Scholar] [CrossRef]
- De Almeida, C.S.; Andrade-Oliveira, V.; Câmara, N.O.S.; Jacysyn, J.F.; Faquim-Mauro, E.L. Crotoxin from Crotalus durissus terrificus is able to down-modulate the acute intestinal inflammation in mice. PLoS ONE 2015, 10, e0121427. [Google Scholar] [CrossRef] [Green Version]
- Pulli, B.; Ali, M.; Forghani, R.; Schob, S.; Hsieh, K.L.C.; Wojtkiewicz, G.; Linnoila, J.J.; Chen, J.W. Measuring myeloperoxidase activity in biological samples. PLoS ONE 2013, 8, e67976. [Google Scholar] [CrossRef] [Green Version]
- Kouyoumdjian, J.A.; Harris, J.B.; Johnson, M.A. Muscle necrosis caused by the sub-units of crotoxin. Toxicon 1986, 24, 575–583. [Google Scholar] [CrossRef]
- Landucci, E.C.T.; Antunes, E.; Donato, J.L.; Faro, R.; Hyslop, S.; Marangoni, S.; Oliveira, B.; Cirino, G.; de Nucci, G. Inhibition of carrageenin-induced rat paw oedema by crotapotin, a polypeptide complexed with phospholipase A2. Br. J. Pharmacol. 1995, 114, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, D.F.; Lopes-Ferreira, M.; Faquim-Mauro, E.L.; Macedo, M.S.; Farsky, S.H.P. Role of crotoxin, a phospholipase A2 isolated from Crotalus durissus terrificus snake venom, on inflammatory and immune reactions. Mediat. Inflamm. 2001, 10, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Massis, L.M.; Assis-Marques, M.A.; Castanheira, F.V.S.; Capobianco, Y.J.; Balestra, A.C.; Escoll, P.; Wood, R.E.; Manin, G.Z.; Correa, V.M.A.; Alves-Filho, J.C.; et al. Legionella longbeachae is immunologically silent and highly virulent in vivo. J. Infect. Dis. 2017, 215, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Mitzner, W. Mechanics of the lung in the 20th century. Compr. Physiol. 2011, 1, 2009–2027. [Google Scholar] [PubMed] [Green Version]
- Faffe, D.S.; Zin, W.A. Lung parenchymal mechanics in health and disease. Physiol. Rev. 2009, 89, 759–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGovern, T.K.; Robichaud, A.; Fereydoonzad, L.; Schuessler, T.F.; Martin, J.G. Evaluation of respiratory system mechanics in mice using the forced oscillation technique. J. Vis. Exp. 2013, e50172. [Google Scholar] [CrossRef]
- Grinnan, D.C.; Truwit, J.D. Clinical review: Respiratory mechanics in spontaneous and assisted ventilation. Crit. Care 2005, 9, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutfi, M.F. The physiological basis and clinical significance of lung volume measurements. Multidiscip. Respir. Med. 2017, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, I.E.; Amarie, O.V.; Mutze, K.; Königshoff, M.; Yildirim, A.Ö.; Eickelberg, O. Systematic phenotyping and correlation of biomarkers with lung function and histology in lung fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L919–L927. [Google Scholar] [CrossRef]
- Vulfius, C.A.; Kasheverov, I.E.; Kryukova, E.V.; Spirova, E.N.; Shelukhina, I.V.; Starkov, V.G.; Andreeva, T.V.; Faure, G.; Zouridakis, M.; Tsetlin, V.I.; et al. Pancreatic and snake venom presynaptically active phospholipases A2 inhibit nicotinic acetylcholine receptors. PLoS ONE 2017, 12, e0186206. [Google Scholar] [CrossRef] [Green Version]
- Hawgood, B.J.; de Sa, S.S. Changes in spontaneous and evoked release of transmitter induced by the crotoxin complex and its component phospholipase A2 at the frog neuromuscular junction. Neuroscience 1979, 4, 293–303. [Google Scholar] [CrossRef]
- Brazil, O.V.; Fontana, M.D.; Heluany, N.F. Nature of the postsynaptic action of crotoxin at guinea-pig diaphragm end-plates. J. Nat. Toxins 2000, 9, 33–42. [Google Scholar]
- Santos, P.E.B.; Souza, S.D.; Freire-Maia, L.; Almeida, A.P. Effects of crotoxin on the isolated guinea pig heart. Toxicon 1990, 28, 215–224. [Google Scholar] [CrossRef]
- Muniz, Z.M.; Diniz, C.R. The effect of crotoxin on the longitudinal muscle-myenteric plexus preparation of the guinea pig ileum. Neuropharmacology 1989, 28, 741–747. [Google Scholar] [CrossRef]
- Radvanyi, F.; Keil, A.; Saliou, B.; Lembezat, M.P.; Bon, C. Binding of divalent and trivalent cations with crotoxin and with its phospholipase and its non-catalytic subunits: Effects on enzymatic activity and on the interaction of phospholipase component with phospholipids. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1989, 1006, 183–192. [Google Scholar] [CrossRef]
- Yen, C.H.; Tzeng, M.C. Identification of a new binding protein for crotoxin and other neurotoxic phospholipase A 2 s on brain synaptic membranes. Biochemistry 1991, 30, 11473–11477. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S. The Pharmacology of Indomethacin. Headache 2016, 56, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Gurney, A.M.; Rang, H.P. The channel-blocking action of methonium compounds on rat submandibular ganglion cells. Br. J. Pharmacol. 1984, 82, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Rang, H.P.; Rylett, R.J. The interaction between hexamethonium and tubocurarine on the rat neuromuscular junction. Br. J. Pharmacol. 1984, 81, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Scannone, H.; Finol, H.J.; Pineda, M.E.; Fernández, I.; Vargas, A.M.; Girón, M.E.; Aguilar, I.; Rodríguez-Acosta, A. Alterations in the ultrastructure of cardiac autonomic nervous system triggered by crotoxin from rattlesnake (Crotalus durissus cumanensis) venom. Exp. Toxicol. Pathol. 2007, 59, 129–137. [Google Scholar] [CrossRef]
- Breithaupt, H. Neurotoxic and myotoxic effects of crotalus phospholipase A and its complex with crotapotin. Naunyn Schmiedebergs Arch. Pharmacol. 1976, 292, 271–278. [Google Scholar] [CrossRef]
- Huaringa, A.J.; Leyva, F.J.; Glassman, A.B.; Haro, M.H.; Arellano-Kruse, A.; Kim, E.E. The lung permeability index: A feasible measurement of pulmonary capillary permeability. Respir. Med. 2011, 105, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Habermann, E.; Walsch, P.; Breithaupt, H. Biochemistry and pharmacology of the crotoxin complex-II. Possible interrelationships between toxicity and organ distribution of phospholipase A, crotapotin and their combination. Naunyn Schmiedebergs Arch. Pharmacol. 1972, 273, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.A.; Silveira, M.B.; Simal, C.; Forte-Dias, C.L.; dos Santos, R.G. Biodistibution and spect imaging of crotoxin on mice bearing Ehrlich tumor. In Proceedings of the International Nuclear Atlantic Conference, Aben, Associação Brasileira De Energia Nuclear, Rio de Janeiro, Brazil, 27 September–2 October 2009. [Google Scholar]
- Sartim, M.A.; Menaldo, D.L.; Sampaio, S.V. Immunotherapeutic potential of Crotoxin: Anti-inflammatory and immunosuppressive properties. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, X.M.; Feldman, J.L. Central cholinergic regulation of respiration: Nicotinic receptors. Acta Pharmacol. Sin. 2009, 30, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.D.; Fryer, A.D. Role of parasympathetic nerves and muscarinic receptors in allergy and asthma. Chem. Immunol. Allergy 2012, 98, 48–69. [Google Scholar] [PubMed] [Green Version]
- Hughes, R.A.; Bihari, D. Acute neuromuscular respiratory paralysis. J. Neurol. Neurosurg. Psychiatry 1993, 56, 334–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolls, M.R.; Voelkel, N.F. Hypoxia and the lung: Beyond hypoxic vasoconstriction. Antioxid. Redox Signal. 2007, 9, 741–743. [Google Scholar] [CrossRef] [Green Version]
- Araneda, O.F.; Tuesta, M. Lung oxidative damage by hypoxia. Oxid. Med. Cell. Longev. 2012, 2012, 856918. [Google Scholar] [CrossRef] [Green Version]
- Wolz-Richter, S.; Esser, K.H.; Hess, A. Antinociceptive activity of crotoxin in the central nervous system: A functional Magnetic Resonance Imaging study. Toxicon 2013, 74, 44–55. [Google Scholar] [CrossRef]
- Wang, J.; Qin, X.; Zhang, Z.; Chen, M.; Wang, Y.; Gao, B. Crotoxin suppresses the tumorigenic properties and enhances the antitumor activity of Iressa® (gefinitib) in human lung adenocarcinoma SPCA-1 cells. Mol. Med. Rep. 2014, 10, 3009–3014. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.P.; Silva, V.A.O.; Silvestrini, A.V.P.; de Macedo, L.H.; Caetano, G.F.; Reis, R.M.; Mazzi, M.V. Crotoxin from Crotalus durissus terrificus venom: In vitro cytotoxic activity of a heterodimeric phospholipase A2 on human cancer-derived cell lines. Toxicon 2018, 156, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Medioni, J.; Brizard, M.; Elaidi, R.; Reid, P.F.; Benlhassan, K.; Bray, D. Innovative design for a phase 1 trial with intra-patient dose escalation: The Crotoxin study. Contemp. Clin. Trials Commun. 2017, 7, 186–188. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sartim, M.A.; Souza, C.O.S.; Diniz, C.R.A.F.; da Fonseca, V.M.B.; Sousa, L.O.; Peti, A.P.F.; Costa, T.R.; Lourenço, A.G.; Borges, M.C.; Sorgi, C.A.; et al. Crotoxin-Induced Mice Lung Impairment: Role of Nicotinic Acetylcholine Receptors and COX-Derived Prostanoids. Biomolecules 2020, 10, 794. https://doi.org/10.3390/biom10050794
Sartim MA, Souza COS, Diniz CRAF, da Fonseca VMB, Sousa LO, Peti APF, Costa TR, Lourenço AG, Borges MC, Sorgi CA, et al. Crotoxin-Induced Mice Lung Impairment: Role of Nicotinic Acetylcholine Receptors and COX-Derived Prostanoids. Biomolecules. 2020; 10(5):794. https://doi.org/10.3390/biom10050794
Chicago/Turabian StyleSartim, Marco Aurelio, Camila O. S. Souza, Cassiano Ricardo A. F. Diniz, Vanessa M. B. da Fonseca, Lucas O. Sousa, Ana Paula F. Peti, Tassia Rafaella Costa, Alan G. Lourenço, Marcos C. Borges, Carlos A. Sorgi, and et al. 2020. "Crotoxin-Induced Mice Lung Impairment: Role of Nicotinic Acetylcholine Receptors and COX-Derived Prostanoids" Biomolecules 10, no. 5: 794. https://doi.org/10.3390/biom10050794