Haplotypes of the Mutated SIRT2 Promoter Contributing to Transcription Factor Binding and Type 2 Diabetes Susceptibility


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Clinical Parameters and Serum Biomarkers
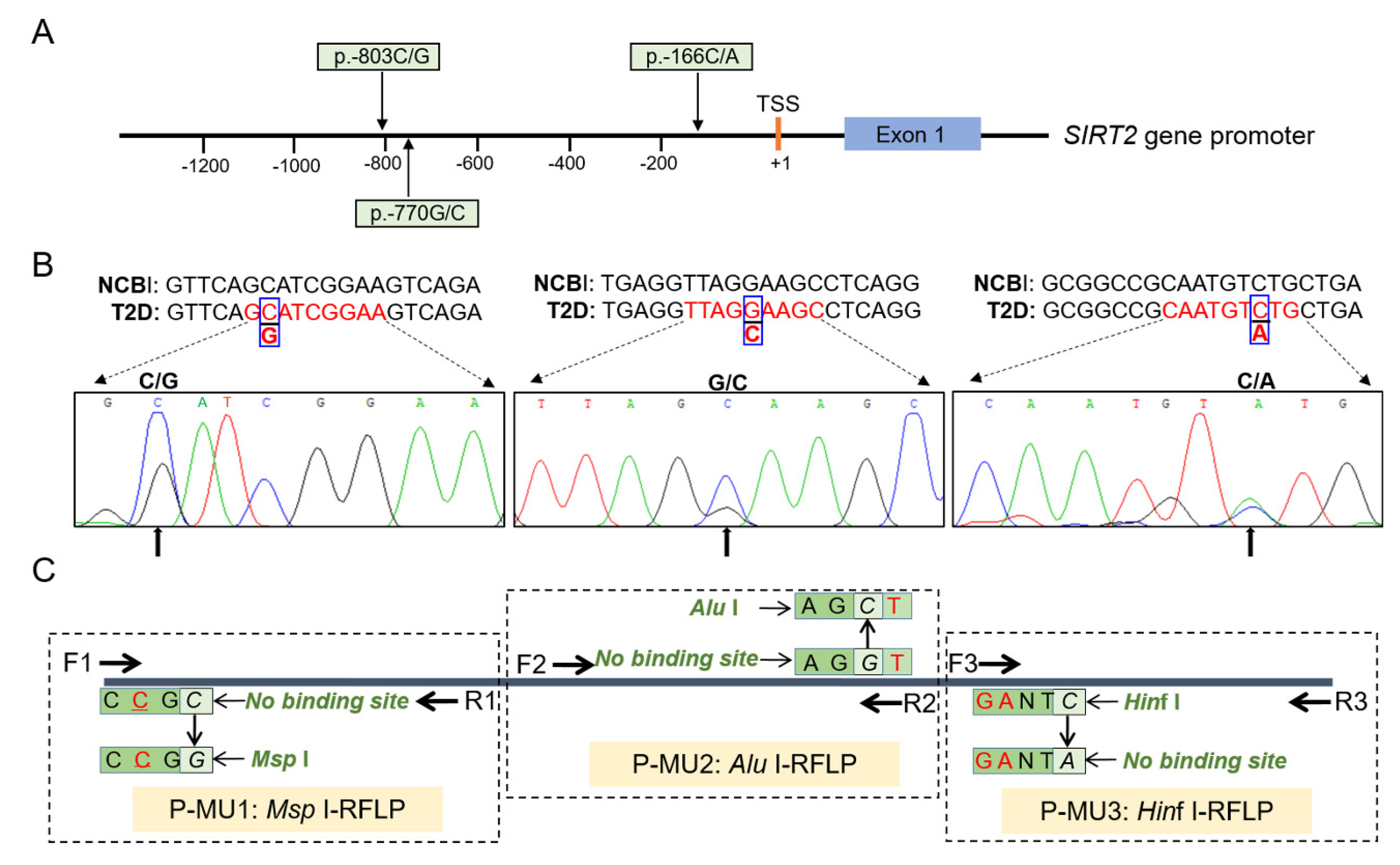
2.3. SNP Genotyping Assay
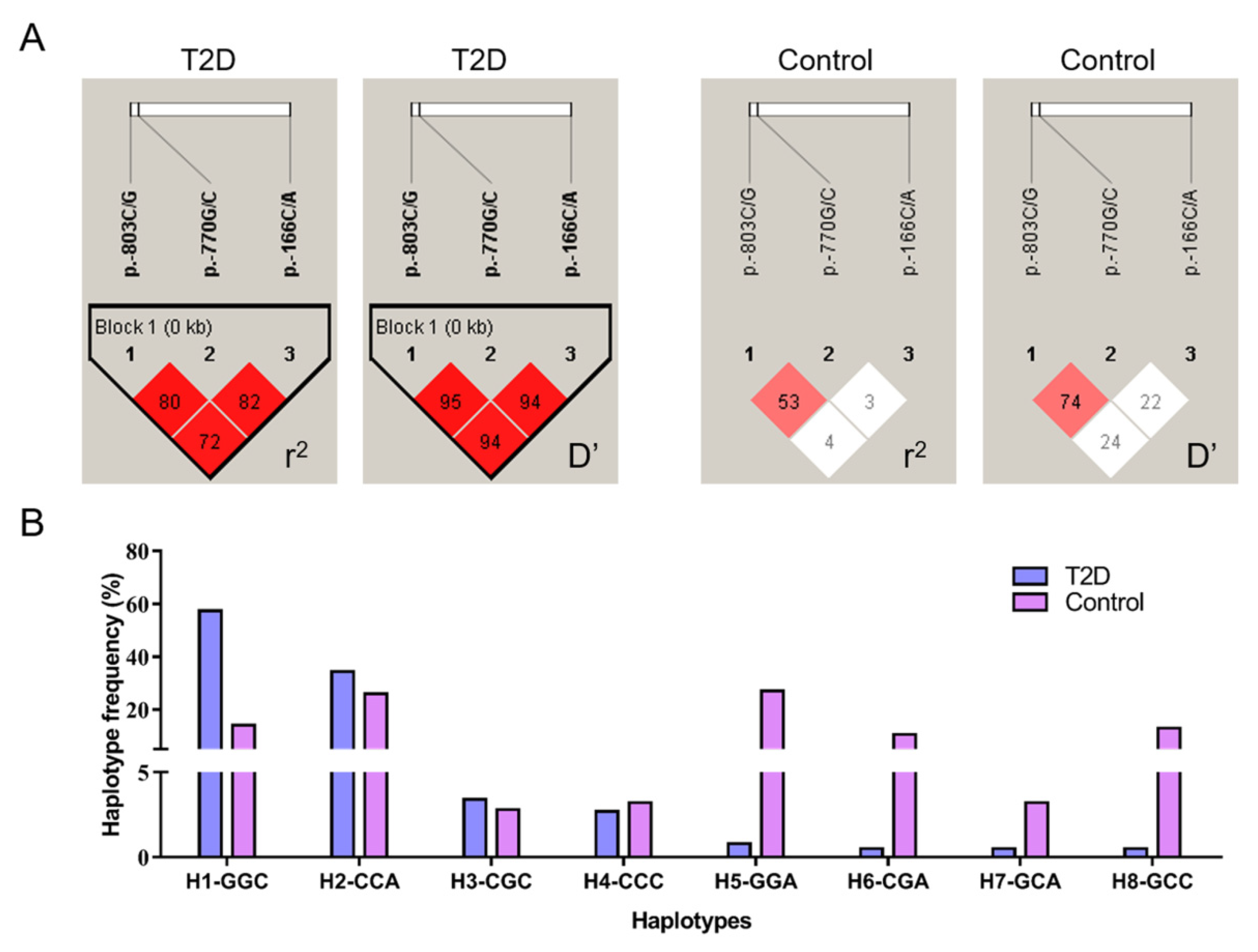
2.4. Linkage Disequilibrium and Haplotype Analysis
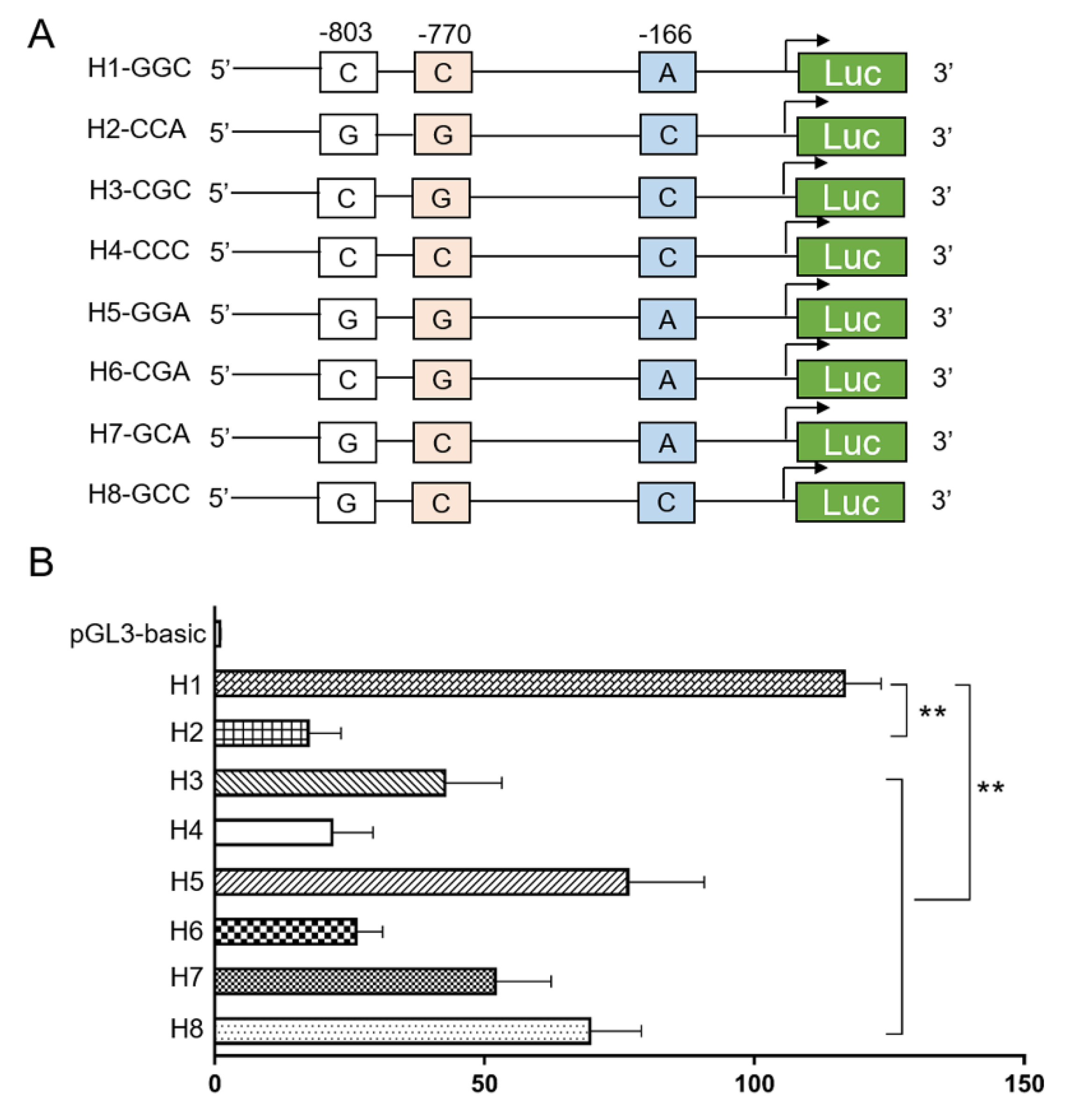
2.5. Promoter Activity Analysis with a Dual-Luciferase Reporter Assay
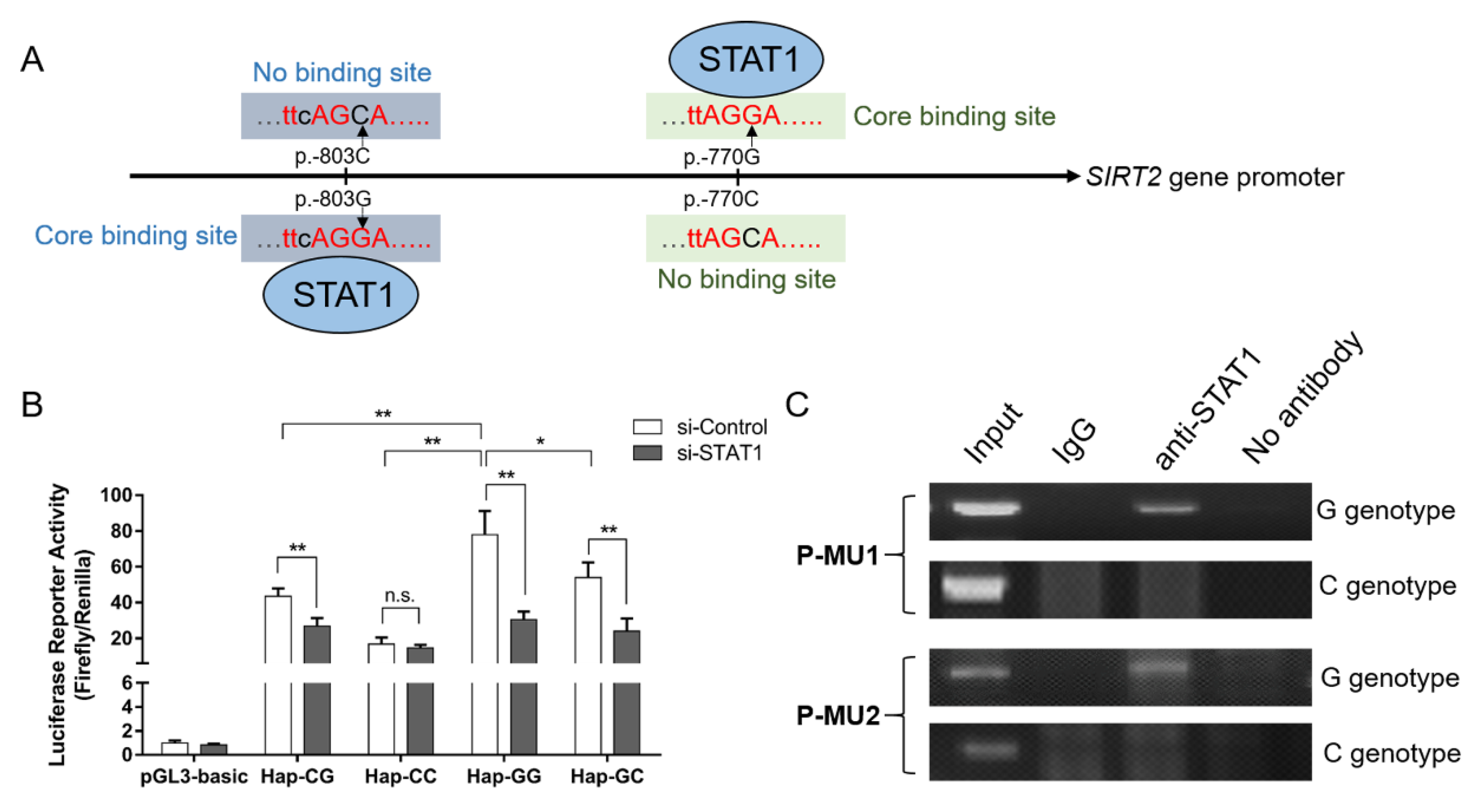
2.6. Chromatin Immunoprecipitation (ChIP) Assay
2.7. Statistical Analysis
3. Results
3.1. Discovery and Genotyping of the SIRT2 SNPs
3.2. Linkage Disequilibrium and Haplotype Combination Analysis
3.3. Population Genetic Analysis and T2D Risk Evaluation
3.4. Effects of the SIRT2 Haplotype Combinations on Diabetic Characteristics
3.5. Genetic Variation Affects the Promoter Activity by Altering Putative Binding Site of STAT1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Persaud, S.J.; Bowe, J.E. Editorial overview: Endocrine and metabolic diseases Druggable diabetes: Identification of therapeutic opportunities. Curr. Opin. Pharmacol. 2018, 43, iii. [Google Scholar] [CrossRef] [PubMed]
- Montesanto, A.; Bonfigli, A.R.; Crocco, P.; Garagnani, P.; De Luca, M.; Boemi, M.; Marasco, E.; Pirazzini, C.; Giuliani, C.; Franceschi, C.; et al. Genes associated with Type 2 Diabetes and vascular complications. Aging 2018, 10, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.H.; Chae, J.; Lee, S.; Choi, S.; Koo, B.K.; Yoon, J.W.; Park, J.H.; Cho, B.; Moon, M.K. Nonsynonymous Variants in PAX4 and GLP1R Are Associated With Type 2 Diabetes in an East Asian Population. Diabetes 2018, 67, 1892–1902. [Google Scholar] [CrossRef] [Green Version]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; Wu, Y.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Yang, B.; Jia, X.; Li, M.; Tan, W.; Ma, S.; Shi, X.; Feng, L. Distinctive Roles of Sirtuins on Diabetes, Protective or Detrimental? Front. Endocrinol. 2018, 9, 724. [Google Scholar] [CrossRef]
- Ye, X.; Li, M.; Hou, T.; Gao, T.; Zhu, W.G.; Yang, Y. Sirtuins in glucose and lipid metabolism. Oncotarget 2017, 8, 1845–1859. [Google Scholar] [CrossRef] [Green Version]
- Sarikhani, M.; Mishra, S.; Desingu, P.A.; Kotyada, C.; Wolfgeher, D.; Gupta, M.P.; Singh, M.; Sundaresan, N.R. SIRT2 regulates oxidative stress-induced cell death through deacetylation of c-Jun NH2-terminal kinase. Cell Death Differ. 2018, 25, 1638–1656. [Google Scholar] [CrossRef] [Green Version]
- Gomes, P.; Fleming Outeiro, T.; Cavadas, C. Emerging Role of Sirtuin 2 in the Regulation of Mammalian Metabolism. Trends Pharmacol. Sci. 2015, 36, 756–768. [Google Scholar] [CrossRef]
- Ramakrishnan, G.; Davaakhuu, G.; Kaplun, L.; Chung, W.C.; Rana, A.; Atfi, A.; Miele, L.; Tzivion, G. Sirt2 deacetylase is a novel AKT binding partner critical for AKT activation by insulin. J. Biol. Chem. 2014, 289, 6054–6066. [Google Scholar] [CrossRef] [Green Version]
- Arab Sadeghabadi, Z.; Nourbakhsh, M.; Pasalar, P.; Emamgholipour, S.; Golestani, A.; Larijani, B.; Razzaghy-Azar, M. Reduced gene expression of sirtuins and active AMPK levels in children and adolescents with obesity and insulin resistance. Obes. Res. Clin. Pract. 2018, 12, 167–173. [Google Scholar] [CrossRef]
- Lantier, L.; Williams, A.S.; Hughey, C.C.; Bracy, D.P.; James, F.D.; Ansari, M.A.; Gius, D.; Wasserman, D.H. SIRT2 knockout exacerbates insulin resistance in high fat-fed mice. PLoS ONE 2018, 13, e0208634. [Google Scholar] [CrossRef]
- Jia, X.; Yang, Y.; Chen, Y.; Xia, Z.; Zhang, W.; Feng, Y.; Li, Y.; Tan, J.; Xu, C.; Zhang, Q.; et al. Multivariate analysis of genome-wide data to identify potential pleiotropic genes for type 2 diabetes, obesity and coronary artery disease using MetaCCA. Int. J. Cardiol. 2019, 283, 144–150. [Google Scholar] [CrossRef]
- Xu, Y.; Song, R.; Long, W.; Guo, H.; Shi, W.; Yuan, S.; Xu, G.; Zhang, T. CREB1 functional polymorphisms modulating promoter transcriptional activity are associated with type 2 diabetes mellitus risk in Chinese population. Gene 2018, 665, 133–140. [Google Scholar] [CrossRef]
- Saini, S.; Mitra, I.; Mousavi, N.; Fotsing, S.F.; Gymrek, M. A reference haplotype panel for genome-wide imputation of short tandem repeats. Nat. Commun. 2018, 9, 4397. [Google Scholar] [CrossRef] [Green Version]
- Olokoba, A.B.; Obateru, O.A.; Olokoba, L.B. Type 2 diabetes mellitus: A review of current trends. Oman Med. J. 2012, 27, 269–273. [Google Scholar] [CrossRef]
- Deepa, M.; Pradeepa, R.; Rema, M.; Mohan, A.; Deepa, R.; Shanthirani, S.; Mohan, V. The Chennai Urban Rural Epidemiology Study (CURES)--study design and methodology (urban component) (CURES-I). J. Assoc. Physicians India 2003, 51, 863–870. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Xu, N.; Wu, H.; Li, D.; Wang, J. Diagnostic accuracy of glycated hemoglobin compared with oral glucose tolerance test for diagnosing diabetes mellitus in Chinese adults: A meta-analysis. Diabetes Res. Clin. Pract. 2014, 106, 11–18. [Google Scholar] [CrossRef]
- Garasia, S.; Samaan, Z.; Gerstein, H.C.; Engert, J.C.; Mohan, V.; Diaz, R.; Anand, S.S.; Meyre, D. Influence of depression on genetic predisposition to type 2 diabetes in a multiethnic longitudinal study. Sci. Rep. 2017, 7, 1629. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, S.K.; Raimondo, A.; Hastoy, B.; Sengupta, S.; Dai, X.Q.; Bautista, A.; Censin, J.; Payne, A.J.; Umapathysivam, M.M.; Spigelman, A.F.; et al. Type 2 diabetes risk alleles in PAM impact insulin release from human pancreatic beta-cells. Nat. Genet. 2018, 50, 1122–1131. [Google Scholar] [CrossRef]
- Watanabe, H.; Inaba, Y.; Kimura, K.; Matsumoto, M.; Kaneko, S.; Kasuga, M.; Inoue, H. Sirt2 facilitates hepatic glucose uptake by deacetylating glucokinase regulatory protein. Nat. Commun. 2018, 9, 30. [Google Scholar] [CrossRef]
- Choudhury, R.R.; Rogivue, A.; Gugerli, F.; Parisod, C. Impact of polymorphic transposable elements on linkage disequilibrium along chromosomes. BMC Bioinform. 2019, 28, 1550–1562. [Google Scholar] [CrossRef] [Green Version]
- Taliun, D.; Gamper, J.; Pattaro, C. Efficient haplotype block recognition of very long and dense genetic sequences. Mol. Ecol. 2014, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Chimusa, E.R.; Meintjies, A.; Tchanga, M.; Mulder, N.; Seoighe, C.; Soodyall, H.; Ramesar, R. A genomic portrait of haplotype diversity and signatures of selection in indigenous southern African populations. PLoS Genet. 2015, 11, e1005052. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Ding, Y.; Zhang, M.; Chen, X.; Wang, X.; Yu, H.; Liu, C.; Lv, H.; Zhang, R. Genome-wide haplotype association study identifies risk genes for non-small cell lung cancer. J. Theor. Biol. 2018, 456, 84–90. [Google Scholar] [CrossRef]
- Shen, G.Q.; Girelli, D.; Li, L.; Olivieri, O.; Martinelli, N.; Chen, Q.; Topol, E.J.; Wang, Q.K. Multi-allelic haplotype association identifies novel information different from single-SNP analysis: A new protective haplotype in the LRP8 gene is against familial and early-onset CAD and MI. Gene 2013, 521, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Nagafuchi, S.; Kamada-Hibio, Y.; Hirakawa, K.; Tsutsu, N.; Minami, M.; Okada, A.; Kai, K.; Teshima, M.; Moroishi, A.; Murakami, Y.; et al. TYK2 Promoter Variant and Diabetes Mellitus in the Japanese. EBioMedicine 2015, 2, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, A.; Furuta, M.; Totoki, Y.; Tsunoda, T.; Kato, M.; Shiraishi, Y.; Tanaka, H.; Taniguchi, H.; Kawakami, Y.; Ueno, M.; et al. Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat. Genet. 2016, 48, 500–509. [Google Scholar] [CrossRef]
- Kwon, E.Y.; Chung, J.Y.; Park, H.J.; Kim, B.M.; Kim, M.; Choi, J.H. OCT3 promoter haplotype is associated with metformin pharmacokinetics in Koreans. Sci. Rep. 2018, 8, 16965. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, Y.; Dorfman, R.G.; Yin, Y.; Zhou, Q.; Huang, S.; Liu, J.; Zhao, S. Sirtinol promotes PEPCK1 degradation and inhibits gluconeogenesis by inhibiting deacetylase SIRT2. Sci. Rep. 2017, 7, 7. [Google Scholar] [CrossRef]
- Arora, A.; Dey, C.S. SIRT2 negatively regulates insulin resistance in C2C12 skeletal muscle cells. Biochim. Biophys. Acta 2014, 1842, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cai, Y.; Huang, H.; Chen, X.; Chen, X.; Chen, X.; Mai, H.; Li, X.; Zhao, J.; Yang, J.; et al. miR-486-3p Influences the Neurotoxicity of a-Synuclein by Targeting the SIRT2 Gene and the Polymorphisms at Target Sites Contributing to Parkinson’s Disease. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 2732–2745. [Google Scholar] [CrossRef]
- Shen, Y.; Chen, L.; Zhang, S.; Xie, L. Correlation Between SIRT2 3′UTR Gene Polymorphism and the Susceptibility to Alzheimer’s Disease. J. Mol. Neurosci. 2020, 1–9. [Google Scholar] [CrossRef]
- Liu, T.; Yang, W.; Pang, S.; Yu, S.; Yan, B. Functional genetic variants within the SIRT2 gene promoter in type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2018, 137, 200–207. [Google Scholar] [CrossRef]
- Gao, X.; Zheng, P.; Yang, L.; Luo, H.; Zhang, C.; Qiu, Y.; Huang, G.; Sheng, W.; Ma, X.; Lu, C. Association of functional variant in GDF1 promoter with risk of congenital heart disease and its regulation by Nkx2.5. Clin. Sci. 2019, 133, 1281–1295. [Google Scholar] [CrossRef]
- Mitchelmore, J.; Grinberg, N.F.; Wallace, C.; Spivakov, M. Functional effects of variation in transcription factor binding highlight long-range gene regulation by epromoters. Nucleic Acids Res. 2020, 48, 2866–2879. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Qian, X.; Wang, B.; Xia, Y.; Zheng, Y.; Du, L.; Xu, D.; Xing, D.; DePinho, R.A.; Lu, Z. Programmable base editing of mutated TERT promoter inhibits brain tumour growth. Nat. Cell Biol. 2020, 22, 282–288. [Google Scholar] [CrossRef]
- Bendaya, I.; Riahi, A.; Kharat, M.; Kahla, S.; Sdiri, W.; Oueslati, R. STAT1 and STAT6 Act as Antagonistic Regulators of PPARγ in Diabetic Patients with and without Cardiovascular Diseases. Clin. Lab. 2018, 64, 287–294. [Google Scholar] [CrossRef]
- Huang, F.; Zhao, Y.; Wang, Q.; Hillebrands, J.L.; van den Born, J.; Ji, L.; An, T.; Qin, G. Dapagliflozin Attenuates Renal Tubulointerstitial Fibrosis Associated with Type 1 Diabetes by Regulating STAT1/TGFβ1 Signaling. Front. Endocrinol. 2019, 10, 441. [Google Scholar] [CrossRef] [Green Version]
- Sultan, S. The effect of maternal type 2 diabetes on fetal endothelial gene expression and function. Acta Diabetol. 2019, 56, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Sun, X.; Li, G.; Wu, Q.; Chen, Y.; Yang, X.; Luo, X.; Hu, J.; Wang, G. Inhibition of SIRT2 limits tumour angiogenesis via inactivation of the STAT3/VEGFA signalling pathway. Cell Death Dis. 2018, 10, 9. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | Group | Genotypic Frequencies Group: T2D (209), Control (223) | Allelic Frequencies | He | PIC | HWE (p) 1 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| P-MU1 (p.-803C/G) | CC | CG | GG | p-value | C | G | ||||
| T2D | 44 (21%) | 63 (30%) | 102 (49%) | 0.002 ** | 36% | 64% | 0.46 | 0.35 | <0.01 | |
| Control | 76 (34%) | 85 (38%) | 62 (28%) | 53% | 47% | 0.50 | 0.37 | <0.01 | ||
| P-MU2 (p.-770G/C) | GG | GC | CC | p-value | G | C | ||||
| T2D | 95 (46%) | 65 (31%) | 49 (23%) | 0.001 ** | 61% | 39% | 0.48 | 0.36 | <0.01 | |
| Control | 54 (24%) | 100 (45%) | 69 (31%) | 47% | 53% | 0.50 | 0.37 | >0.05 a | ||
| P-MU3 (p.-166C/A) | CC | CA | AA | p-value | C | A | ||||
| T2D | 48 (23%) | 75 (36%) | 86 (41%) | 0.194 | 41% | 59% | 0.48 | 0.37 | <0.01 | |
| Control | 49 (22%) | 103 (46%) | 71 (32%) | 45% | 55% | 0.50 | 0.37 | >0.05 a | ||
| Loci | Haplotype Combinations | T2D (n) | Control (n) | p-Value 2 | Odds Ratio | 95% CI |
|---|---|---|---|---|---|---|
| p.-803C/G p.-770G/C p.-166C/A | H2-H2-CCCCAA | 40 | 12 | 1 | ||
| H1-H1-GGGGCC | 85 | 29 | 0.02 * | 2.03 | 1.12–3.72 | |
| H1-H2-GCGCCA | 57 | 42 | 0.16 | 0.89 | 0.51–3.24 | |
| H1-H4-GCGCCC | 7 | 14 | 0.11 | 1.37 | 0.69–3.91 | |
| p-value 1 | 0.001 ** | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Li, J.; Sheng, J.; Dai, Y.; Wang, Y.; Liu, J.; Xu, Y. Haplotypes of the Mutated SIRT2 Promoter Contributing to Transcription Factor Binding and Type 2 Diabetes Susceptibility. Genes 2020, 11, 569. https://doi.org/10.3390/genes11050569
Zheng X, Li J, Sheng J, Dai Y, Wang Y, Liu J, Xu Y. Haplotypes of the Mutated SIRT2 Promoter Contributing to Transcription Factor Binding and Type 2 Diabetes Susceptibility. Genes. 2020; 11(5):569. https://doi.org/10.3390/genes11050569
Chicago/Turabian StyleZheng, Xiao, Jiajun Li, Jie Sheng, Yang Dai, Yue Wang, Jinbiao Liu, and Yao Xu. 2020. "Haplotypes of the Mutated SIRT2 Promoter Contributing to Transcription Factor Binding and Type 2 Diabetes Susceptibility" Genes 11, no. 5: 569. https://doi.org/10.3390/genes11050569