
Abstract
Key message
Xylem functionality is the primary determinant of the Ca relations of developing sweet cherry. Calcium influx is positively related to transpiration and decreases as xylem functionality is progressively lost during development.
Abstract
Fruit calcium (Ca) may be associated with susceptibility to rain cracking in sweet cherry but little background information is available on the fruits’ Ca relations. The objectives of the study were to identify the key determinants of the Ca content in developing sweet cherry fruit. The masses of Ca and K in the flesh (mg per fruit) increased continuously through to maturity. However, their dry mass ratios [mg per g dry mass (DM)] decreased continuously. The decrease in Ca/dry mass ratio was to about 30% of the transition stage II/III value, and for K to about 55%. These decreases occurred in all cultivars investigated. Moreover, the Ca mass per fruit and the Ca/dry mass ratio were normally distributed and wide ranging; the maximum recorded values were about twice the minimum ones. Within a fruit, the Ca/dry mass ratio was two to three times higher in the stem end than in the stylar scar end. Fruit held under low relative humidity (26.2%) contained more Ca per fruit and had a higher Ca/dry mass ratio compared with fruit held under high humidity (91.6%). There was a positive relationship between Ca mass per fruit and cumulative transpiration. Our results indicate xylem functionality is the primary determinant of the Ca relations of sweet cherry fruit.
Similar content being viewed by others


Introduction
Calcium plays an important role in the pre- and postharvest physiology of most fruit. Functions of Ca include signal transduction as a secondary messenger (Hardingham and Bading 1999; Steinhorst and Kudla 2014), the maintenance and regulation of membrane semi-permeability (van Steveninck 1965; Poovaiah and Leopold 1973; Hepler 2005) and the structural reinforcement and cross-linking of cell-wall constituents (Demarty et al. 1984; Chan et al. 2017). The latter functions are associated with apoplastic Ca. This Ca pool constitutes by far the largest in a fruit (Demarty et al. 1984).
Because fruit firmness is an important quality attribute in many fruit crop species, pre- and postharvest applications of Ca are commonly made to improve these properties, including in apple (Conway et al. 2002; Schlegel and Schönherr 2002), tomato (Garcia et al. 1995; Islam et al. 2016) and sweet cherry (Winkler and Knoche 2019).
In sweet cherry, Ca is reported to positively affect fruit-quality attributes such as firmness and cracking susceptibility [for review see Winkler and Knoche (2019)]. However, the effects are not always consistent. Thus, cracking susceptibility is markedly reduced by Ca treatments in some studies (Bullock 1952; Glenn and Poovaiah 1989; Meheriuk et al. 1991; Michailidis et al. 2017), while in other studies no Ca effect was found (Callan 1986; Koffmann et al. 1996; Looney 1985). To properly exploit the potentially positive effects of Ca applications on fruit quality in commercial production systems requires a better understanding of the Ca relations of developing fruit. To our knowledge, there are no detailed reports on fruit Ca relations in sweet cherry. In particular, the import of Ca to the fruit and its subsequent distribution within the fruit have not been investigated on a temporospatial basis. Because Ca transport is restricted to the xylem (Marschner 1995) and because, in a sweet cherry fruit, xylem functionality decreases during development (Brüggenwirth et al. 2016; Grimm et al. 2017; Winkler et al. 2016), the import of Ca to the fruit is most likely to be slowed and changes in fruit Ca relations are predicted to occur. In particular, during the later stage of fruit development, Ca import is expected to decline. Coincidentally, this will occur at much the same time as cracking susceptibility is known to increase.
Also, studies on Ca relations indicate that Ca transport is driven by transpiration. Hence, manipulating transpiration affected Ca import as was shown in kiwi when subjected to differential transpiration (Montanaro et al. 2015) or in tomato following spray application of abscisic acid (ABA) (de Freitas et al. 2014).
In contrast to Ca, K occurs at high concentrations in the phloem. Hence, plant organs preferentially supplied by phloem sap such as fruit are typically rich in K (Mengel and Kirkby 1982). Three variables are useful in describing the changing mineral composition of a developing fruit: (a) solute mass per volume, (b) solute mass per fruit and (c) solute/dry mass ratio.
-
a
Solute mass per volume (e.g., mg/ml)—describes the concentration of each soluble moiety in the fruit per volume of solvent (water). Solute mass per volume is particularly relevant to fruit chemistry because chemical reactions are mostly concentration dependent. However, with strong tissue and cellular compartmentation of solutes, solute concentration measurements commonly offer an averaged, whole-fruit or whole-tissue, value. These values can obscure quite wide concentration differences among the various compartments, so are not very useful.
-
b
Solute mass per fruit (e.g., mg per fruit)—describes for each solute moiety the net of the influxes/effluxes through the fruit-stalk vasculature that have occurred up to that stage of fruit development. Thus, changes in solute mass per fruit (the first differential of mass per fruit with respect to time) can be taken as indicative of the rates and directions of the phloem and xylem sap flows (the only pathways through which most of these solutes can enter/leave the fruit). This is particularly useful because some solutes (e.g., sugars) are transported exclusively in the phloem, while others (e.g., Ca) are transported exclusively in the xylem (Marschner 1995). Moreover, while (it is believed) the phloem sap flows always from tree to fruit, the xylem sap can flow in either direction. That is, the speed and direction of xylem sap flow depend on the magnitude and direction of the water potential gradient between tree and fruit. With wide diurnal oscillations in tree water potential being the norm, the xylem sap can flow from fruit to tree in daytime and from tree to fruit at night (Lang 1990).
-
c
Solute/dry mass ratio (e.g., mg/g dry mass)—describes the mass of a particular solute moiety in relation to the total dry mass of a tissue. Changes with time in the solute/dry mass ratio are useful in similar ways to those of solute mass per fruit, providing current information regarding the solute influxes/effluxes in the xylem and phloem saps. However, while solute mass per fruit is subject to very wide statistical variability between apparently similar fruits, the normalization offered by the solute/dry mass ratio can greatly reduce this variability.
The objectives of this study were (1) to quantify the Ca and K contents of developing sweet cherry fruit, (2) to determine the natural variation in Ca content of sweet cherry within the canopy (fruit-to-fruit variability with position in the tree) and across a range of cultivars (fruit-to-fruit variability between cultivars), (3) to identify potential relationships between fruit transpiration and fruit Ca import and (4) to quantify the effects of abscisic acid (ABA) on the Ca contents of fruit and leaves. Ca content was quantified as the Ca mass per fruit (mg per fruit) and the Ca/dry mass ratio (mg per g DM).
Materials and methods
Plant material
Sweet cherry fruit from the cultivars Adriana, Hedelfinger, Sam, Schneiders Späte Knorpel, Staccato and Sweetheart were sampled from greenhouse-grown or field-grown trees grafted on ‘Gisela 5’ rootstocks (P. cerasus L. × P. canescens Bois) at the Horticultural Research Station of Leibniz University in Ruthe, Germany (lat. 52° 14′ N, long. 9° 49′ E). The only exception was the ‘Regina’ fruit used in the study on the developmental time course of changes in Ca, K and C content. For this, fruit was sourced from a commercial sweet cherry orchard in Gleidingen, Germany (lat. 52° 16′ N, long. 9° 50′ E). The trees were trained to a slender spindle. The soil nutrient level was optimum for potassium (18–20 mg per 100 g soil). Soil pH was slightly acidic (pH 5.5) to neutral (pH 6.0) for the loamy sand at Gleidingen and the loess at Ruthe, respectively. All fruits were cultivated according to current European regulations for integrated fruit production. No foliar fertilizers containing either Ca or K were applied to any trees used in this study.
General experimental procedure
Determination of fresh and dry mass
Fruit flesh and pit were separated mechanically and weighed individually. Pit and flesh samples were frozen at − 20 °C, freeze-dried for 3 to 4 days and then dried in an oven for 14 days at 103 °C and re-weighed.
Contents of Ca, K and C
Unless specified otherwise, the Ca content was determined only for the flesh. Flesh and pit were separated mechanically. Preliminary experiments established that destruction of the cuticle was essential for efficient lyophilization. Hence, stage III fruits were incubated in liquid nitrogen and subsequently crushed and freeze-dried for 3 to 4 days. For flesh samples of stage III fruit, an aliquot of about 8 g was dried in an oven at 103 °C for at least three to 4 days before grinding. For samples of stage II fruit, oven drying was not required, samples were ground directly. Grinding was carried out in a MM 400 mill (Retsch, Haan, Germany; 50-ml grinding jar, frequency 30 Hz, duration 10 s for flesh or 30 s for pits). A 1 g sample of the powder was taken and further dried to constant mass for at least 3 to 4 days at 103 °C. A 100 mg aliquot was taken and ashed in a muffle furnace (L24/11/B180; Nabertherm, Lilienthal, Germany) at 500 °C (heating phase: from 20 to 500 °C 2 h, holding phase: 4 h at 500 °C). Occasionally ashing was incomplete. These samples were taken up in 200 µl of 1 N HCl and re-ashed using the same settings. The ash was then taken up in 2 ml of 1 N HCl plus 8 ml of deionized water and filtered (MN 640 M; Macherey-Nagel, Dueren, Germany). LaCl3 was added to the solutions at a final concentration of 1% to eliminate interference in the Ca analyses by any P in the samples (Fishman and Downs 1966).The filtrate was diluted with deionized water as required to obtain a Ca concentration in the measuring window (0 to 4 mg l− 1). Samples were analyzed using atomic absorption spectroscopy (AAS) (AAnalayst 300; Perkin Elmer, Waltham, MA, USA) equipped with a Ca lumina hollow cathode lamp (wavelength 422.7 nm, slit 0.7 nm) using an air–acetylene flame. Following an additional dilution step, the K content of the solutions was quantified by atomic emission spectroscopy (1100B; Perkin Elmer).
The C mass of the flesh in developing ‘Regina’ fruit was analyzed following drying and grinding. An aliquot of 30.0–41.5 mg of dried powder was weighed into stannic cartridges for subsequent determination of C content using an elemental analyzer (vario EL cube; Elementar Analysensysteme, Langenselbold, Germany).
Each sampling comprised three biological replicates of 10–20 fruits each and two to three technical replicates. The only exceptions were samples taken at 15 and 23 days after full bloom (DAFB) during the developmental time course, when 30–53 fruits were used.
Experiments
Developmental time course
The developmental time courses of change in the Ca and K mass of flesh and pit, dry mass ratios of Ca and K and in C mass of the flesh were determined in developing ‘Regina’ fruit between 15 DAFB and 86 DAFB. Here, the Ca contents of flesh and pit were determined.
Effect of cultivar
Putative differences in Ca mass per fruit and Ca/dry mass ratio among cultivars were investigated in ‘Adriana’, ‘Hedelfinger’, ‘Sam’, ‘Schneiders Späte Knorpel’, ‘Staccato’ and ‘Sweetheart’. The Ca was determined in immature fruit (transition stage II/III, straw yellow) and also in mature fruit.
Fruit-to-fruit variability
The fruit-to-fruit variability in Ca mass per fruit and Ca/dry mass ratio was determined in mature ‘Sweetheart’. Here 40 randomly sampled fruit were analyzed individually.
Position in the canopy
Fruits taken from different positions within the canopy were analyzed for their Ca mass and Ca/dry mass ratio in ‘Adriana’. Row orientation was from southwest to northeast. Comparisons were made between (1) top center, (2) mid-center and (3) base-center, and on either side of the row, i.e., (4) mid-northwest, (5) mid-southeast, (6) base-northwest and (7) base-southeast. All mid positions were at a height of about 1.5 m above the ground and all base positions were at about 0.5 m above the ground. The northwest and southeast orientations faced the alleys on either side of the row. Three biological replicates of ten fruits each per position were sampled from three different trees.
Axial gradient within the fruit
Putative axial gradients in Ca/dry mass ratio from the fruits stylar ends to their stem ends were investigated in ‘Sweetheart’ and ‘Merchant’ sweet cherry fruit. Fruits were cut transversely (i.e., normal to the stem/stylar-scar axis) into four slices of equal thickness, and the pit was removed. The slices were frozen, freeze-dried and analyzed as described above. The number of replicates was ten.
Effect of transpiration
Putative relationships between fruit transpiration and Ca mass per fruit or between fruit transpiration and Ca/dry mass ratio were investigated by manipulating the transpiration of greenhouse-grown ‘Sam’ fruit beginning at 13 DAFB. Working with fruit still attached to the tree, transpiration was changed by enclosing individual fruit in transparent polypropylene pottles (volume 120 ml). To increase transpiration, relative humidity (RH) was lowered by placing 25 g of dry silica gel in each pottle. To minimize transpiration, RH was increased by fruit transpiration in empty pottles. The fruit was introduced to the pottle through a 12-mm-diameter hole cut in the screw lid. The gap between stem and lid was sealed with a foam plug. The plug arrangement significantly reduced water vapor gain/loss from the pottle through the hole in the lid. Each setup was attached to the tree using thin wire. Sensors (logger: MSR147WD, sensor: FH2.3/160; MSR Electronics, Seuzach, Switzerland) were used to monitor the temperatures and humidities inside four such pottles—two representing the low-RH treatment (silica gel) and the other two the high-RH treatment (no silica gel) (Fig. 1). In total, 150 fruits were mounted in pottles with silica gel and 150 fruits in empty pottles. In the pottles containing dry silica gel, a low RH (mean 26.2%) and thus high water vapor pressure deficit (WVPD, mean 1.8 kPa) was established. In contrast, in the pottles with fruit but no silica gel, the RH was elevated (mean 91.6%) and the WVPD was low (mean 0.3 kPa; Fig. 1). The pottles with dry silica gel were weighed and the silica gel was replaced weekly with freshly dried silica gel. Separate pottles without fruit but with dried silica gel served as controls to quantify diffusion of water vapor into the vessel. A third group of fruit remained hanging in situ in the tree (no pottle), being directly exposed to the greenhouse microclimate. For this group, the mean RH was 76.4% (corresponding to a WVPD of 0.7 kPa). Fruits were sampled at 27, 37, 48, 58 and 69 DAFB and fresh mass (FM), dry mass as well as Ca mass per fruit and Ca/dry mass ratios were quantified as described above. Three biological replications of ten fruits each were sampled for each treatment and date.

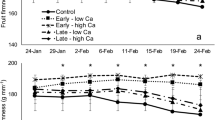
Relative humidity (a, c, e) and water vapor pressure deficit (WVPD) (b, d, f) of fruit held at low (a, b) and high relative humidity (RH) (c, d) and control fruit (e, f). The low and high-RH environments were generated by sealing still-attached fruit within plastic pottles containing dry silica gel (low RH) or not (high RH). Fruits not within pottles (i.e., directly exposed to the greenhouse atmosphere) served as controls. The silica gel was replaced periodically. Repeated weighing of the silica gel allowed fruit transpiration to be quantified. There were no differences in temperature (c inset) between the treatments. Trees were grown in a greenhouse. Numbers in the graphs are means ± SE
Effect of abscisic acid
The effects of abscisic acid (ABA) on Ca mass and Ca/dry mass ratios of ‘Sam’ sweet cherry were investigated. Whole trees were sprayed to run-off using a commercial formulation of ABA (ProTone SL; Valent BioSciences, Libertyville, IL, USA). The treatments were applied at weekly intervals starting from 14 DAFB to 81 DAFB (maturity) at a concentration of 400 mg l− 1. The non-ionic surfactant Tween 20 (Sigma-Aldrich, St. Louis, MO, USA) was added at a concentration of 0.05%. The effect of ABA on transpiration was measured 24 h after spray application using a leaf porometer (SC-1; Meter Group, Pullman, Washington, USA). The Ca mass per fruit and the Ca/dry mass ratios of fruit and leaves were analyzed at maturity as described above.
Data analysis
Except for Figs. 1 and 2 data are presented as means ± SE. Where error bars are not visible in a graph, they are smaller than the plotted symbols. The WVPD was calculated from temperature and relative humidity for every 5 min interval logged using the following equation (Allen et al. 1999):

Developmental time course of change in fresh mass and dry mass (DM) (a) and Ca mass and K mass per fruit (b) and Ca/dry mass ratio and K/dry mass ratio in sweet cherry (c). Dashed vertical lines indicate transition between stage I and II and between stage II and III development
In the next step, the cumulative transpiration of the low-humidity (high transpiration) treatment was calculated. The increase in mass of the silica in the pottle, which was determined at weekly intervals, was accumulated for the time periods from the initial mounting of the pottle at 13 DAFB until harvest at 27, 37, 48, 58 or 69 DAFB. Regressing the so-obtained cumulative transpiration against the cumulative WVPD in the pottle (calculated for the 5-min intervals as described above and then accumulated for the same time period as the cumulative transpiration was calculated) yielded a significant relationship. The equation was \({\text{Transpiration}} \left({\text{g}}\right)=2.84\times{10}^{-4}{\text{(g kPa}^{-1})} \times {\text{WVPD}} \left({\text{kPa}}\right)\), r2 = 0.97**. This regression equation allowed prediction of transpiration from the WVPD value for the free-hanging control fruit and for the fruit of the high-humidity treatment.
Regression analyses were carried out using R (function lm, version 3.5.1; R Foundation for Statistical Computing, Vienna, Austria) and SigmaPlot (version 12.5; Systat Software, San Jose, CA, USA). SigmaPlot (version 12.5; Systat Software) was used to create the figures. The significances of coefficients of determination (r² at P ≤ 0.05, 0.01 and 0.001) are indicated by *, ** and ***, respectively.
Results
As expected, fruit fresh (FM) mass and dry mass (DM) increased with time (Fig. 3a). The increases resulted primarily from increases in flesh FM and DM. Pit FM and DM increased only until about 37 DAFB (FM) and 65 DAFB (DM) and remained constant thereafter.

Relationship between K mass and the dry mass (a) and the C mass (b) per fruit of developing sweet cherry. (b inset) Relationship between C mass per fruit and dry mass per fruit
Flesh DM (%) increased only slowly up to about 58 DAFB and then increased rapidly. For the pit, DM (%) followed a sigmoidal pattern:
(\({\text{DM }} \left(\%\right)=5.70+\frac{56.73}{1+{\text{e}}^{\frac{-\left(\text{Time } \right({\text{DAFB}})-45.87)}{7.83}}}\), data not shown).
In the flesh, Ca mass per fruit and (especially) K mass per fruit increased markedly up till maturity (Fig. 3b). In contrast, Ca and K mass in the pit increased only to about 51 DAFB. Thereafter, Ca in the pit remained constant, while K in the pit decreased (Fig. 3b). On a dry mass ratio basis, Ca in the flesh decreased to about 30% during the developmental period and that in the pit to about 84%. In the flesh, the K/dry mass ratio decreased from about 15 to 37 DAFB followed by a transient increase from about 37 to 65 DAFB. Thereafter, the K/dry mass ratio remained about constant. In the pit, the K/dry mass ratio decreased from about 22 mg at 15 DAFB to about 2 mg at 65 DAFB and remained about constant thereafter (Fig. 3c).
The mass of K per fruit was linearly related to fruit dry mass (Fig. 4a) and the amount of C per fruit (Fig. 4b) indicating that the C content of the dry mass was constant (Fig. 4b inset).

Normal probability plots of Ca mass per fruit (a) and the Ca/dry mass ratio of single sweet cherry fruit (b). Insets: histograms of frequency distributions
The marked decrease of the Ca/dry mass ratio between fruits at the transition stage II/III and at the mature stage III was not unique for ‘Regina’, but also occurred to similar extents in other cultivars (Table 1). Marked differences existed between cultivars. At the transition stage II/III, the range was 1.6-fold from the lowest to the highest Ca/dry mass ratio and this increased to 2.7-fold at the fully mature stage. The larger range at the mature stage III was due in part to differences among cultivars in the increase in fruit mass that diluted the Ca present.
The mass of Ca per fruit and the Ca/dry mass ratio followed normal distributions as indicated by the linear normal probability plots. Ranges in Ca per fruit and Ca/dry mass ratio were both about twofold, with coefficients of variation of 22.3 and 18.4%, respectively (Fig. 2).
Within a mature sweet cherry fruit, the distribution of Ca was not uniform. There was a gradient in Ca along the stem/stylar scar axis. The Ca/dry mass ratios were two- (‘Sweetheart’) to threefold (‘Merchant’) higher in the stem end than in the stylar scar end (Table 2).
There were no significant effects of the position of the fruit in the canopy on fruit size, Ca per fruit or Ca/dry mass ratios (results not shown).
Manipulating the RH around individual fruit in the greenhouse resulted in significant effects on Ca per fruit and on Ca/dry mass ratio. The effects on fruit size were generally not significant. Fruits held under a low-RH environment (high transpiration) were slightly smaller than others held under a high-RH environment (low transpiration) (Fig. 5a). Free-hanging control fruit exposed to a variable and intermediate RH environment (variable and intermediate transpiration) tended to be largest. There were no differences in the temperatures between the free-hanging control fruit and the fruit enclosed in the pottles as well as between the low-RH and high-RH pottles (data not shown).

Developmental time course of change in fruit fresh mass (a), Ca mass per fruit (b) and Ca/dry mass ratio of developing sweet cherry (c). The developmental time courses were established for fruit held at low relative humidity (RH) (mean 26.2%) or high RH (mean 91.6%). The low RH was established by sealing a still-attached fruit in a plastic pottle containing dry silica gel. The high RH was established by sealing a still-attached fruit in an empty pottle. Free-hanging, still-attached fruit exposed to the greenhouse atmosphere served as controls (mean RH 76.4%). DM dry mass. The dashed vertical line indicates the transition from stage II to stage III
The low-RH fruit generally had a significantly higher Ca mass per fruit and a higher Ca/dry mass ratio compared with that of the high-RH fruit (Fig. 5b, c). Control fruit exposed to the greenhouse climate was intermediate in Ca mass per fruit and Ca/dry mass ratio. Consistent with the developmental time course study, Ca/dry mass ratios decreased during development under all treatments. The difference in Ca mass per fruit resulted from differences in cumulative transpiration. The low-RH environment resulted in high cumulative transpiration and, vice versa, the high-RH environment resulted in low cumulative transpiration (Fig. 6a). The free-hanging control fruit was intermediate (Fig. 6a). These differences were all significant. Expressing Ca mass per fruit as a function of cumulative transpiration revealed a positive, non-linear relationship (Fig. 6b). As cumulative transpiration increased, Ca mass per fruit increased at a decreasing rate, i.e., the curve tended to asymptote for values of high cumulative transpiration.

a Cumulative transpiration and b Ca mass per fruit of developing sweet cherry that remained attached to the tree and held at low relative humidity (RH) (mean 26.2%) or high RH (mean 91.6%). The low RH was established by sealing a still-attached fruit in a plastic pottle containing dry silica gel. The high RH was established by sealing a still-attached fruit in an empty plastic pottle. Free-hanging, still-attached fruit exposed to the greenhouse atmosphere served as controls (mean RH 76.4%). The dashed vertical line in a indicates the transition from stage II to stage III
Spray application of ABA decreased stomatal conductance of leaves by about 20% (Table 3). However, there was no effect on either the fruit or the leaf Ca/dry mass ratios or the Ca mass per fruit (Table 3).
Discussion
The results indicate (1) that the import of Ca into developing sweet cherries does not keep pace with the increase in fruit mass and (2) that Ca import into developing sweet cherry is positively related to changes in fruit transpiration.
Ca import does not keep pace with fruit growth
The increase in Ca mass per fruit lagged behind the increase in fruit FM and DM. As a consequence, Ca/dry mass ratio decreased. This effect is not unique to ‘Regina’, but has also been observed in ‘Sam’ (Fiedler, unpublished data) and in ‘Lambert’ (Facteau 1982). In the latter cultivar, Ca mass per fruit remained constant during the last 2 weeks before harvest. Decreases in Ca/dry mass ratio during fruit development have also been reported in apple (Amarante et al. 2013), bean, pepper, quince (Mix and Marschner 1976a, b), kiwifruit (Clark and Smith 1991; Xiloyannis et al. 2001), mango (Joyce et al. 2001), papaya (Qiu et al. 1995) and tomato (Ehret and Ho 1986).
Ca translocation is limited to the xylem (Marschner 1995). Thus, the lack of proportionality between fruit growth and fruit Ca import was likely due to a progressive loss of xylem functionality in the developing sweet cherry. During stage III, the contribution of xylem water inflow to the fruit decreases from 95% of total inflow at the transition stage II/III to about 5% of the total at the mature stage III (Brüggenwirth et al. 2016; Winkler et al. 2016). Furthermore, the decrease in xylem functionality occurs progressively, beginning at the stylar end and progressing towards the stem end (Grimm et al. 2017; Winkler et al. 2016). Accordingly, the lowest Ca/dry mass ratio was recorded in the stylar scar end, where xylem functionality was first lost; and the highest Ca/dry mass ratio was recorded in the stem end of the fruit, where the xylem remained functional through to maturity (Grimm et al. 2017; Winkler et al. 2016). Similar observations have been reported for apple (Lewis and Martin 1973), avocado (Burdon et al. 1991; Chaplin and Scott 1980), sweet pepper (Marcelis and Ho 1999) and tomato (Adams and Ho 1993; Ehret and Ho 1986; Nonami et al. 1995). Thus, xylem functionality seems to be the primary determinant of fruit Ca distribution within a sweet cherry.
In contrast to Ca, plant organs that are preferentially supplied by phloem sap such as fruit are typically rich in K (Mengel and Kirkby 1982). It is, therefore, not surprising that the K mass per fruit and the K/dry mass ratio both increased linearly with the increase in fruit dry matter and, hence, with total C import into the fruit. During stage III, water inflow via the phloem accounts for an increasing proportion of the total inflow until, at maturity, essentially all water inflow to the fruit occurs via the phloem (Brüggenwirth et al. 2016). This explains why the K/dry mass ratio changes only slightly during fruit development. The transient increase in K/dry mass ratio between 42 and 68 DAFB is difficult to explain. This effect was not due to biological variation, but it was reproducible in ‘Sam’ sweet cherry during another growing season (Fiedler, unpublished data).
Increased transpiration increases Ca import to the fruit
Our results establish a close relationship between fruit transpiration and Ca import to sweet cherry. Low-transpiration fruits had a Ca mass of about 0.6 mg per fruit at maturity, while high-transpiration fruits had a Ca mass averaging about 1.1 mg per fruit. This relationship is not linear, with the nearly twofold increase in fruit Ca being associated with a nearly 6.4-fold increase in fruit transpiration (Fig. 6b). Increases in fruit Ca with increased transpiration have also been observed in apple (Cline and Hanson 1992), kiwifruit (Montanaro et al. 2015) and snap bean (Grusak and Pomper 1999). Also, as in sweet cherry, the relationship between fruit transpiration and fruit Ca content was not linear in apple (Tromp and Vanvuure 1993) or in kiwifruit (Montanaro et al. 2015).
These observations are accounted for by the progressive change in xylem functionality in developing fruit. At the stage II/III transition, the xylem is fully functional and xylem flow and fruit transpiration are closely related (Brüggenwirth et al. 2016). After the stage II/III transition, transpiration becomes increasingly independent of xylem inflow as the latter tends to decrease (Brüggenwirth et al. 2016; Winkler et al. 2016). Thus, during early development, fruit transpiration has a strong effect on fruit Ca import, but the effect is much weaker at later stage III and at maturity.
Furthermore, convective flow within the xylem sap is not the only mechanism of Ca transport. Three additional mechanisms are involved. (1) Ca translocation occurs by movement along Ca-exchange sites on the walls of vessels and pit membranes of the xylem (Clarkson 1984; Kuhn et al. 1995). (2) The acropetal Ca transport is linked to the basipetal auxin transport. An inhibition of basipetal transport of IAA restricts the acropetal transport of Ca (Banuelos et al. 1987; Tomala and Dilley 1990). (3) Transport of Ca also occurs via Ca2+-transporters or Ca2+-binding proteins (Hirschi 2004; Park et al. 2005). The combined effects of all three mechanisms would account for the lack of a simple linear relationship between cumulative fruit transpiration and cumulative fruit Ca mass. These effects may also account for the absence of any consistent effects of position in the canopy on fruit Ca mass. We expected fruit high in the canopy to experience higher transpiration and, hence, to exhibit higher Ca mass per fruit than the more shaded fruit in the base or center of the canopy. This was not the case.
Practical implications
The influx of Ca to developing fruit is slowed by the progressive loss of xylem function. Hence, agronomic practices that may increase fruit transpiration may also increase fruit Ca—but only if these are applied early on in fruit development. These practices might include a more open canopy structure. Closing of rain shelters at (or shortly after) bloom will keep the canopy and the fruit surfaces dry and will also increase their temperature. Provided proper ventilation also occurs, these conditions (i.e., warmer and drier) will increase the water vapor pressure deficit, and hence fruit transpiration, and hence fruit Ca Whether the resulting increases in fruit Ca are sufficient to significantly reduce cracking susceptibility has yet to be demonstrated.
Author contribution statement
MK obtained the funds to support the study. MK and AW planned the experiments. AW and BF conducted the experiments. MK and AW analysed the data, wrote, revised and edited the manuscript.
References
Adams P, Ho LC (1993) Effects of environment on the uptake and distribution of calcium in tomato and on the incidence of blossom-end rot. Plant Soil 154:127–132. https://doi.org/10.1007/Bf00011081
Allen RG, Pereira LS, Raes D, Smith M (1999) Crop evapotranspiration. Guidelines for computing crop water requirements. FAO Irrigation and Drainage Paper No 56 FAO, Rome
Amarante CVT, Miqueloto A, Steffens CA, dos Santos A, Argenta LC (2013) Changes in xylem functionality during apple fruit development: Implications on calcium concentration and incidence of bitter pit. Acta Hortic 1012:135–140. https://doi.org/10.17660/ActaHortic.2013.1012.11
Banuelos GS, Bangerth F, Marschner H (1987) Relationship between polar basipetal auxin transport and acropetal Ca2+ transport into tomato fruits. Physiol Plant 71:321–327. https://doi.org/10.1111/j.1399-3054.1987.tb04350.x
Brüggenwirth M, Winkler A, Knoche M (2016) Xylem, phloem, and transpiration flows in developing sweet cherry fruit. Trees 30:1821–1830. https://doi.org/10.1007/s00468-016-1415-4
Bullock RM (1952) A study of some inorganic compounds and growth promoting chemicals in relation to fruit cracking of Bing cherries at maturity. Proc Am Soc Hortic Sci 56:243–253
Burdon JN, Moore KG, Wainwright H (1991) Mineral distribution in mango fruit susceptible to the physiological disorder soft-nose. Sci Hortic 48:329–336. https://doi.org/10.1016/0304-4238(91)90143-M
Callan NW (1986) Calcium hydroxide reduces splitting of ‘Lambert’ sweet cherry. J Am Soc Hortic Sci 111:173–175
Chan SY, Choo WS, Young DJ, Loh XJ (2017) Pectin as a rheology modifier: origin, structure, commercial production and rheology. Carbohydr Polym 161:118–139. https://doi.org/10.1016/j.carbpol.2016.12.033
Chaplin GR, Scott KJ (1980) Association of calcium in chilling injury susceptibility of stored avocados. HortScience 15:514–515
Clark CJ, Smith GS (1991) Seasonal changes in the form and distribution of calcium in fruit of kiwifruit vines. J Hortic Sci 66:747–753. https://doi.org/10.1080/00221589.1991.11516207
Clarkson DT (1984) Calcium transport between tissues and its distribution in the plant. Plant Cell Environ 7:449–456. https://doi.org/10.1111/j.1365-3040.1984.tb01435.x
Cline JA, Hanson EJ (1992) Relative humidity around apple fruit influences its accumulation of calcium. J Am Soc Hortic Sci 117:542–546. https://doi.org/10.21273/JASHS.117.4.542
Conway WS, Sams CE, Hickey KD (2002) Pre- and postharvest calcium treatment of apple fruit and its effect on quality. Acta Hortic 594:413–419. https://doi.org/10.17660/ActaHortic.2002.594.53
de Freitas ST, McElrone AJ, Shackel KA, Mitcham EJ (2014) Calcium partitioning and allocation and blossom-end rot development in tomato plants in response to whole-plant and fruit-specific abscisic acid treatments. J Exp Bot 65:235–247. https://doi.org/10.1093/jxb/ert364
Demarty M, Morvan C, Thellier M (1984) Calcium and the cell wall. Plant Cell Environ 7:441–448. https://doi.org/10.1111/j.1365-3040.1984.tb01434.x
Ehret DL, Ho LC (1986) Translocation of calcium in relation to tomato fruit growth. Ann Bot 58:679–688. https://doi.org/10.1093/oxfordjournals.aob.a087230
Facteau TJ (1982) Levels of pectic substances and calcium in gibberellic acid-treated sweet cherry fruit. J Am Soc Hortic Sci 107:148–151
Fishman MJ, Downs SC (1966) Methods for analysis of selected metals in water by atomic absorption. Geological surves water-supply paper 1540-C. U.S. Government Printing Office, Washington, DC
Garcia JM, Ballesteros JM, Albi MA (1995) Effect of foliar applications of CaCl2 on tomato stored at different temperatures. J Agric Food Chem 43:9–12. https://doi.org/10.1021/jf00049a003
Glenn GM, Poovaiah BW (1989) Cuticular properties and postharvest calcium applications influence cracking of sweet cherries. J Am Soc Hortic Sci 114:781–788
Grimm E, Pflugfelder D, van Dusschoten D, Winkler A, Knoche M (2017) Physical rupture of the xylem in developing sweet cherry fruit causes progressive decline in xylem sap inflow rate. Planta 246:659–672. https://doi.org/10.1007/s00425-017-2719-3
Grusak MA, Pomper KW (1999) Influence of pod stomatal density and pod transpiration on the calcium concentration of snap bean pods. J Am Soc Hortic Sci 124:194–198. https://doi.org/10.21273/Jashs.124.2.194
Hardingham GE, Bading H (1999) Calcium as a versatile second messenger in the control of gene expression. Microsc Res Tech 46:348–355. https://doi.org/10.1002/(Sici)1097-0029(19990915)46:6<348::Aid-Jemt3>3.3.Co;2-1
Hepler PK (2005) Calcium: A central regulator of plant growth and development. Plant Cell 17:2142–2155. https://doi.org/10.1105/tpc.105.032508
Hirschi KD (2004) The calcium conundrum. Both versatile nutrient and specific signal. Plant Physiol 136:2438–2442. https://doi.org/10.1104/pp.104.046490
Islam MZ, Mele MA, Baek JP, Kang HM (2016) Cherry tomato qualities affected by foliar spraying with boron and calcium. Hortic Environ Biotechnol 57:46–52. https://doi.org/10.1007/s13580-016-0097-6
Joyce DC, Shorter AJ, Hockings PD (2001) Mango fruit calcium levels and the effect of postharvest calcium infiltration at different maturities. Sci Hortic 91:81–99. https://doi.org/10.1016/S0304-4238(01)00247-3
Koffmann W, Wade NL, Nicol H (1996) Tree sprays and root pruning fail to control rain induced cracking of sweet cherries. Plant Prot Q 11:126–130
Kuhn AJ, Bauch J, Schröder WH (1995) Monitoring uptake and contents of Mg, Ca and K in Norway spruce as influenced by pH and Al, using microprobe analysis and stable isotope labelling. Plant Soil 168:135–150. https://doi.org/10.1007/Bf00029322
Lang A (1990) Xylem, phloem and transpiration flows in developing apple fruits. J Exp Bot 41:645–651. https://doi.org/10.1093/jxb/41.6.645
Lewis TL, Martin D (1973) Longitudinal distribution of applied calcium, and of naturally occurring calcium, magnesium, and potassium, in Merton apple fruits. Aust J Agric Res 24:363–371. https://doi.org/10.1071/Ar9730363
Looney NE (1985) Benefits of calcium sprays below expectations in B.C. tests. GoodFruit Grower 36:7–8
Marcelis LFM, Ho LC (1999) Blossom-end rot in relation to growth rate and calcium content in fruits of sweet pepper (Capsicum annuum L.). J Exp Bot 50:357–363. https://doi.org/10.1093/jxb/50.332.357
Marschner H (1995) Mineral nutrition of higher plants, 2nd edn. Academic Press, London
Meheriuk M, Neilsen GH, McKenzie D-L (1991) Incidence of rain splitting in sweet cherries treated with calcium or coating materials. Can J Plant Sci 71:231–234. https://doi.org/10.4141/cjps91-032
Mengel K, Kirkby EA (1982) Principles of plant nutrition, 3rd edn. International Potash Institute Bern, Switzerland
Michailidis M, Karagiannis E, Tanou G, Karamanoli K, Lazaridou A, Matsi T, Molassiotis A (2017) Metabolomic and physico-chemical approach unravel dynamic regulation of calcium in sweet cherry fruit physiology. Plant Physiol Biochem 116:68–79. https://doi.org/10.1016/j.plaphy.2017.05.005
Mix GP, Marschner H (1976a) Calciumgehalte in Früchten von Paprika, Bohne, Quitte und Hagebutte im Verlauf des Fruchtwachstums. Z Pflanzenernähr Bodenkd 5:537–549
Mix GP, Marschner H (1976b) Einfluß exogener und endogener Faktoren auf den Calciumgehalt von Paprika- und Bohnenfrüchten. Z Pflanzenernähr Bodenkd 5:551–563
Montanaro G, Dichio B, Lang A, Mininni AN, Xiloyannis C (2015) Fruit calcium accumulation coupled and uncoupled from its transpiration in kiwifruit. J Plant Physiol 181:67–74. https://doi.org/10.1016/j.jplph.2015.04.004
Nonami H, Fukuyama T, Yamamoto M, Yang L, Hashimoto Y (1995) Blossom-end rot of tomato plants may not be directly caused by calcium deficiency. Acta Hortic 396:107–114. https://doi.org/10.17660/ActaHortic.1995.396.11
Park S, Cheng NH, Pittman JK, Yoo KS, Park J, Smith RH, Hirschi KD (2005) Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol 139:1194–1206. https://doi.org/10.1104/pp.105.066266
Poovaiah BW, Leopold AC (1973) Deferral of leaf senescence with calcium. Plant Physiol 52:236–239. https://doi.org/10.1104/pp.52.3.236
Qiu YX, Nishina MS, Paull RE (1995) Papaya fruit growth, calcium uptake, and fruit ripening. J Am Soc Hortic Sci 120:246–253. https://doi.org/10.21273/JASHS.120.2.246
Schlegel TK, Schönherr J (2002) Stage of development affects penetration of calcium chloride into apple fruits. J Plant Soil Sci 165:738–745. https://doi.org/10.1002/jpln.200290012
Steinhorst L, Kudla J (2014) Signaling in cells and organisms - calcium holds the line. Curr Opin Plant Biol 22:14–21. https://doi.org/10.1016/j.pbi.2014.08.003
Tomala K, Dilley DR (1990) Some factors influencing the calcium level in apple fruits. Acta Hortic 274:481–488. https://doi.org/10.17660/ActaHortic.1990.274.61
Tromp J, Vanvuure J (1993) Accumulation of calcium, potassium and magnesium in apple fruits under various conditions of humidity. Physiol Plant 89:149–156. https://doi.org/10.1111/j.1399-3054.1993.tb01798.x
van Steveninck RFM (1965) The significance of calcium on the apparent permeability of cell membranes and the effects of substitution with other divalent ions. Physiol Plant 18:54–69. https://doi.org/10.1111/j.1399-3054.1965.tb06869.x
Winkler A, Brüggenwirth M, Ngo NS, Knoche M (2016) Fruit apoplast tension draws xylem water into mature sweet cherries. Sci Hortic 209:270–278. https://doi.org/10.1016/j.scienta.2016.06.041
Winkler A, Knoche M (2019) Calcium and the physiology of sweet cherries: a review. Sci Hortic 245:107–115. https://doi.org/10.1016/j.scienta.2018.10.012
Xiloyannis C, Celano G, Montanaro G, Dichio B, Sebastiani L, Minnocci A (2001) Water relations, calcium and potassium concentration in fruits and leaves during annual growth in mature kiwifruit plants. Acta Hortic. https://doi.org/10.17660/ActaHortic.2001.564.14
Acknowledgements
Open Access funding provided by Projekt DEAL. We thank Friederike Schroeder and Simon Sitzenstock for technical support, Dr. Sandy Lang for helpful comments on an earlier version of this manuscript and Dr. Peter D. Petracek, Valent BioSciences, Libertyville, for the gift of ProTone. This research was funded in part by a grant from the Deutsche Forschungsgemeinschaft (KN402/14-1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by L. Kalcsits.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Winkler, A., Fiedler, B. & Knoche, M. Calcium physiology of sweet cherry fruits. Trees 34, 1157–1167 (2020). https://doi.org/10.1007/s00468-020-01986-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-020-01986-9