


Abstract
Human ether-a-go-go–related gene (hERG) encodes the pore-forming subunit of the rapidly activating delayed rectifier potassium current (IKr) important for repolarization of cardiac action potentials. Drug-induced disruption of hERG channel function is a main cause of acquired long QT syndrome, which can lead to ventricular arrhythmias and sudden death. Illicit fentanyl use is associated with sudden death. We have demonstrated that fentanyl blocks hERG current (IhERG) at concentrations that overlap with the upper range of postmortem blood concentrations in fentanyl-related deaths. Since fentanyl can cause respiratory depression and electrolyte imbalances, in the present study we investigated whether certain pathologic circumstances exacerbate fentanyl-induced block of IhERG. Our results show that chronic hypoxia or hypokalemia additively reduced IhERG with fentanyl. As well, high pH potentiated the fentanyl-mediated block of hERG channels, with an IC50 at pH 8.4 being 7-fold lower than that at pH 7.4. Furthermore, although the full-length hERG variant, hERG1a, has been widely used to study hERG channels, coexpression with the short variant, hERG1b (which does not produce current when expressed alone), produces functional hERG1a/1b channels, which gate more closely resembling native IKr. Our results showed that fentanyl blocked hERG1a/1b channels with a 3-fold greater potency than hERG1a channels. Thus, in addition to a greater susceptibility due to the presence of hERG1b in the human heart, hERG channel block by fentanyl can be exacerbated by certain conditions, such as hypoxia, hypokalemia, or alkalosis, which may increase the risk of fentanyl-induced ventricular arrhythmias and sudden death.
SIGNIFICANCE STATEMENT This work demonstrates that heterologously expressed human ether a-go-go–related gene (hERG) 1a/1b channels, which more closely resemble rapidly activating delayed rectifier potassium current in the human heart, are blocked by fentanyl with a 3-fold greater potency than the previously studied hERG1a expressed alone. Additionally, chronic hypoxia, hypokalemia, and alkalosis can increase the block of hERG current by fentanyl, potentially increasing the risk of cardiac arrhythmias and sudden death.
Introduction
In recent years, fentanyl-associated death has become a serious issue in North America (Alexander et al., 2016; Torralva and Janowsky, 2019). Fentanyl is a synthetic μ-opioid receptor agonist (Raynor et al., 1994). It is used clinically for management of severe pain and combined with other drugs for anesthesia (Stanley, 2014). However, when used illicitly, fentanyl has a propensity to cause sudden death. It is known that fentanyl acts at opioid receptors in the brainstem (Wamsley, 1983), resulting in respiratory depression (McQueen, 1983), which is believed to underlie fentanyl-induced sudden death. However, we have found that fentanyl blocks human ether-a-go-go–related gene (hERG) potassium channels heterologously expressed in human embryonic kidney (HEK) 293 cells (Tschirhart et al., 2019) at submicromolar concentrations that are within the upper range of blood concentrations in fentanyl-related deaths (3–383 μg/l or 8.9 nM to 1.14 μM) (Martin et al., 2006).
hERG encodes the pore-forming subunit of the rapidly activating delayed rectifier potassium current (IKr), which is important for repolarization of cardiac action potentials (Sanguinetti et al., 1995; Trudeau et al., 1995). A decrease in hERG current (IhERG) caused by mutations or drugs can prolong cardiac action potential duration (APD), leading to long QT syndrome (LQTS) (Curran et al., 1995; Sanguinetti et al., 1995; Keating and Sanguinetti, 2001). Notably, LQTS predisposes affected individuals to a polymorphic ventricular tachycardia known as Torsade de Pointes (Keating and Sanguinetti, 2001), which can lead to sudden death (Chiang and Roden, 2000). In fact, many cases of drug-induced sudden death are caused by blockade of hERG channels, and several drugs have been removed from market because of this reason (Fenichel et al., 2004; Roden, 2004). Fentanyl can prolong cardiac action potentials recorded from isolated canine Purkinje fibers (Blair et al., 1989) and neonatal rat ventricular myocytes (Tschirhart et al., 2019). Thus, we proposed that blockade of hERG channels may be involved in fentanyl-induced death of individuals after overdose or in the context of compromised cardiac repolarization, which can occur in various conditions. For example, a reduction in serum K+ concentration, known as hypokalemia, causes LQTS (Curry et al., 1976; Roden et al., 1986). We previously demonstrated that a reduction in extracellular K+ concentration decreases hERG protein density in the plasma membrane and causes LQTS in a rabbit model (Guo et al., 2009). As well, we have shown that chronic hypoxia reduces IhERG (Lamothe et al., 2017). In addition, acidosis accelerates channel deactivation and decreases amplitude of IhERG (Anumonwo et al., 1999; Terai et al., 2000; Van Slyke et al., 2012; Shi et al., 2019). Since fentanyl may cause respiratory depression and electrolyte imbalances (e.g., hypokalemia and altered pH), we thus examined the effects of fentanyl on hERG currents under conditions such as hypoxia, hypokalemia, and altered pH.
Like other potassium channels, a single hERG channel is composed of four α-subunits that assemble to form a tetramer (MacKinnon, 1991; Jiang et al., 2003). Two variants of hERG1 have been identified in myocardium of humans and animals: hERG1a and hERG1b (Lees-Miller et al., 1997; London et al., 1997; Jones et al., 2004, 2014; Phartiyal et al., 2007). Since hERG1b (shorter form, containing 820 amino acids) alone does not effectively produce current in expression systems (Phartiyal et al., 2008; McNally et al., 2017), full-length hERG1a, containing 1159 amino acids, has generally been used to characterize the biophysical and pharmacological properties of the channel defined as the hERG channel. However, when coexpressed, hERG1b and hERG1a can effectively form heterotetramic channels that behave more like native cardiac IKr (London et al., 1997; Phartiyal et al., 2007, 2008; Sale et al., 2008; Larsen and Olesen, 2010) and display altered sensitivity to certain drugs compared with hERG1a alone (Abi-Gerges et al., 2011). Thus, we examined the effects of fentanyl on hERG1a/1b heterotetramic channels.
The present study revealed that chronic hypoxia, hypokalemia, and increased pH exacerbated fentanyl-induced block of hERG channels. Furthermore, heterotetrameric hERG1a/1b channels were blocked with a greater potency than hERG1a (generally referred to as hERG) homotetrameric channels previously studied. By utilizing several hERG N-deletion mutants that accelerate hERG deactivation, we found a correlation between fentanyl block potency and the rate of hERG channel deactivation. Thus, we propose that several conditions may enhance the block of hERG channels by fentanyl, thereby increasing the risk of QT interval prolongation and sudden death.
Materials and Methods
Molecular Biology.
hERG1a (defined as hERG) cDNA was obtained from Dr. Gail Robertson (University of Wisconsin-Madison, Madison, WI). Point mutations Y652A and F656T and the Δ2–354 deletion were constructed using polymerase chain reaction overlap extension technique, as described previously (Guo et al., 2006; Massaeli et al., 2010). The Δ2–9 and Δ2–25 hERG mutants were also constructed using the polymerase chain reaction method and confirmed by DNA sequencing (GENEWIZ, South Plainfield, NJ). The HEK293 cell line stably expressing wild-type (WT) hERG (hERG1a) channels (hERG-HEK cell line) was provided by Dr. Craig January (University of Wisconsin-Madison). hERG1b cDNA and the HEK 293 cell line stably expressing hERG1a plus pTet-One controlled hERG1b expression (hERG1a/1b-HEK cell line) were obtained from Dr. Gail Robertson (University of Wisconsin-Madison). G418 (5 μg/ml) and puromycin (0.25 μg/ml) were used to maintain expression of hERG1a and hERG1b, respectively. Expression of hERG1b was induced by treating cells with 200 ng/ml doxycycline in media for 48 hours before being used for experiments. In addition, transfection of HEK293 cells stably expressing hERG1a with hERG1b plasmid was performed to study hERG1a/1b channels. The HEK293 cell line stably expressing Δ2–354 hERG was created using transfection followed by G418 selection (1 mg/ml) and maintenance (0.4 mg/ml). Mutant hERG Y652A, Δ2–9, and Δ2–25 were transiently expressed in HEK293 cells. For coexpression of WT and Δ2–354 hERG, 2 µg of Δ2–354 hERG plasmid was mixed with 0.5 µg GFP plasmid and transfected into hERG-HEK293 cells cultured in 35-mm plates. For transient transfection, GFP plasmid (pIRES2-EGFP, Clontech, CA) was cotransfected with the plasmid of interest at a 1:4 ratio for the selection of transfected cells during patch-clamp experiments. HEK293 cells were cultured in Gibco minimum essential medium (MEM) supplemented with 10% FBS, nonessential amino acids (glycine, L-alainine, L-asparagine, L-aspartic acid, L-glutamic acid, L-proline, and L-serine, 100 μM each), and 1 mM sodium pyruvate (Thermo Fisher Scientific, Waltham, MA). Cells were passaged every 2 to 3 days and were used for experiments 16–24 hours after passage. During passages, cell culture media were removed, and then the cells were rinsed with phosphate-buffered saline, treated with 0.25% trypsin for 1 minute, suspended in MEM supplemented with 10% FBS, collected for experiments, or plated for culture. Cells were maintained in a 37°C/5% CO2 incubator. For electrophysiological recordings, the cells were collected during passaging and kept in MEM with 10% FBS at room temperature until used (within 4 hours).
Electrophysiological Recordings.
Currents were recorded from HEK cells using the whole-cell voltage-clamp method. Pipettes were pulled from thin-walled (1.5-mm) borosilicate glass (TW150-6; World Precision Instruments, Sarasota, CA) with a P-1000 micropipette puller (Sutter Instrument, Novato, CA) and polished with heat to a resistance of ∼2.0 MΩ when filled with solution. An Axopatch 200B Amplifier, Digidata 1440A Digitizer, and Clampex version 10.7 software (Molecular Devices, San Jose, CA) were used for data acquisition and analysis. The pipette solution contained the following (in millimolars): 135 KCl, 5 EGTA, 5 MgATP, and 10 HEPES (pH 7.2 with KOH). The bath solution contained the following (in millimolars): 5 KCl, 135 NaCl, 2 CaCl2, 1 MgCl2, 10 glucose, and 10 HEPES (pH 7.4 with NaOH). To create pH 6.4 or 8.4 bath solution, concentrated HCl or NaOH was added to the pH 7.4 solution. For acute concentration-response relationships, control current was first recorded in the absence of drug. Increasing concentrations of drug were then added, and the currents after steady state of block (within 1 minute after drug application) were normalized to control and plotted as relative currents against concentrations to construct concentration-response relationships. Recordings were carried out at room temperature (22 ± 1°C).
Peak tail currents upon repolarizing step to −50 mV after the 50-mV depolarization were used to analyze the amplitude of IhERG. To calculate the IC50 values, currents at individual fentanyl concentrations were normalized to the control current (in the absence of drug), summarized, and plotted against concentrations in a log scale. The data were fitted to the Hill equation (y = (A1 − A2) / (1 + (x/xo)p) + A2) in Microcal Origin (OriginLab, Northampton, MA) to obtain IC50 values and Hill coefficients. The deactivation time course was analyzed in Clampfit. The decay of tail currents was fitted to a double exponential function to obtain the fast time constant of hERG deactivation (τf-deact) and slow time constant of hERG deactivation (τs-deact). Each cell was individually fitted, and the values are shown in the scatter plots.
Hypoxic Culture.
For culture in hypoxic conditions, a C-Chamber incubator subchamber with a ProOx 110 oxygen controller (BioSpherix, Parish, NY) was used. 0.5% O2 was maintained with an influx of 95% N2/5% CO2 gas mixture, and temperature was maintained at 37°C. Cells were passaged and incubated for 6 hours. Control cells were maintained in the same incubator but outside of the hypoxic subchamber. Cells were collected for electrophysiological studies by trypsinization, resuspended in media, and used within 1 hour.
Drugs and Reagents.
Fentanyl citrate was purchased from Toronto Research Chemicals (North York, ON, Canada). The powder form of fentanyl citrate was handled in a biologic safety cabinet with proper protective equipment to prevent exposure. Doxycycline hyclate was purchased from BioShop Canada Inc. (Burlington, ON, Canada). Drugs were dissolved in double distilled water, aliquoted, and stored at −20°C. For patch-clamp experiments, fentanyl was diluted in bath solution and used within 8 hours. When investigating mutant channels or different conditions, data were always paired with WT or control (CTL) from the same days with the same drug stock solutions. For acute block, control solution was drained from the bath with a suction pump, and drug-containing solution was applied through a gravity-fed perfusion system. MEM, FBS, Lipofestamine 2000, G418 (Geneticin), trypsin, nonessential amino acids, and sodium pyruvate were purchased from Thermo Fisher Scientific (Waltham, MA). Electrolytes, EGTA, HEPES, and glucose were purchased from Sigma-Aldrich (St. Louis, MO).
Statistical Analysis.
Data are expressed as the mean ± S.D. or scatter plots. Data were analyzed using a one-way ANOVA with Tukey’s post hoc test or a two-tailed Student’s t test with GraphPad Prism (GraphPad Software, San Diego, CA). A P value of 0.05 or less was considered to be statistically significant.
Results
Chronic Hypoxia and Acute Fentanyl Decrease IhERG in an Additive Manner.
Chronic hypoxia impairs hERG function (Lamothe et al., 2017) and prolongs QT intervals in patients (Kenigsberg et al., 2007; Mehra et al., 2009). To investigate the concurrent effects of chronic hypoxia and fentanyl on hERG channels, we cultured hERG-HEK cells in hypoxic (0.5% O2) or normoxic (21% O2, control) conditions for 6 hours and then examined the block effect of 1 μM fentanyl on IhERG. As we reported previously (Lamothe et al., 2017; Tschirhart et al., 2019), compared with control culture, 6-hour culture in 0.5% O2 or acute application of 1 μM fentanyl significantly decreased IhERG. The combination of hypoxic culture and 1 μM fentanyl resulted in IhERG that was significantly less than either condition alone. These results are shown in Fig. 1 and analyzed with one-way ANOVA with Tukey’s post hoc test (**P < 0.01, comparisons as indicated). In addition, to determine whether chronic hypoxic culture alters channel sensitivity to fentanyl, we compared the extents of fentanyl-induced block in normoxic and hypoxic cultured cells. In hypoxic cultured cells, 1 μM fentanyl blocked IhERG by 59% ± 12%, which was not significantly different from 1 μM fentanyl-induced block in normoxic cultured cells (by 52% ± 11%, P > 0.05, unpaired Student's t test). Thus, chronic hypoxia and fentanyl have an additive effect in reducing IhERG.

Combined effects of chronic hypoxia and acute fentanyl on IhERG. hERG-HEK cells were cultured in normoxia (CTL, 21% O2) or hypoxia (Hx, 0.5% O2) for 6 hours. For each cell, IhERG was recorded in the absence or presence of 1 μM fentanyl (FENT). Representative current traces elicited with the voltage protocol are shown (top). The start-to-start interval was 15 seconds. Peak tail currents upon −50-mV repolarization after the 50-mV depolarizing step were used for analysis and are summarized (bottom). Lines represent the mean for each group. CTL, n = 12; FENT, n = 12; Hx, n = 10; Hx+FENT, n = 10 from three independent experiments. **P < 0.01, one-way ANOVA with Tukey’s post hoc test.
Hypokalemia Augments Fentanyl-Induced Blockade of hERG.
To study the effects of hypokalemia and fentanyl on IhERG, we cultured hERG-HEK cells in normal medium (CTL, 5 mM K+) or 2 mM K+ for 72 hours. In hypokalemic patients, serum potassium as low as 1.2 mM K+ was reported (Garcia et al., 2008). Thus, investigating the effects of 2 mM K+o plus fentanyl is of clinical relevance. After culture, we examined the blocking effects of fentanyl on IhERG recorded in standard, 5 mM K+-containing bath solution. Compared with CTL, culture in 2 mM K+ medium for 72 hours or acute application of 1 μM fentanyl significantly decreased IhERG. The combination of 2 mM K+ culture and 1 μM fentanyl resulted in IhERG that was significantly less than either condition alone. These results are shown in Fig. 2A and analyzed with one-way ANOVA with Tukey’s post hoc test (*P < 0.05, **P < 0.01, comparisons as indicated). When recorded in standard 5 mM bath solution, 1 μM fentanyl blocked IhERG from cells cultured in 2 mM K+ medium by 59% ± 8%, which was not significantly different than block from cells cultured in normal, 5 mM K+ medium (59% ± 10%, P > 0.05). Therefore, chronic hypokalemia and fentanyl additively reduce IhERG.

Combined effects of 2 mM K+o and fentanyl on IhERG. (A) hERG-HEK cells were cultured in normal, 5 mM K+o (CTL) or 2 mM K+o (2K) for 72 hours. IhERG was then recorded in the absence or presence of 1 μM fentanyl (FENT) in normal, 5-mM K+ bath solution. Representative currents and summarized IhERG amplitudes (peak tail currents after 50-mV depolarization) are shown. Lines represent the mean. CTL, n = 21, and FENT, n = 21, from six independent experiments; 2K+, n = 12, and 2K+ + FENT, n = 12, from three independent experiments. (B) hERG-HEK cells were cultured in normal media (5 mM K+o). IhERG elicited with the protocol above the current traces was recorded in 5 mM K+o (5K+) or 2 mM K+o solution (2K+) in the absence or presence of 1 μM fentanyl. The start-to-start interval was 15 seconds. Currents in the presence of fentanyl were normalized to the currents before drug and summarized. Lines represent the mean. 5K+, n = 18; 2K+, n = 18. *P < 0.05; **P < 0.01, one-way ANOVA with Tukey’s post hoc test (A) or two-tailed unpaired t test (B).
To further study the effects of acute extracellular K+ concentration on fentanyl-induced block, we examined fentanyl block of IhERG recorded in 5- or 2-mM K+-containing bath solution from cells cultured in normal medium. To clearly demonstrate the degree of block, we normalized current after fentanyl block to control current in each recording condition (Fig. 2B). Although 1 μM fentanyl blocked IhERG by 54% ± 9% in 5 K+ bath solution, it blocked IhERG by 60% ± 6% in 2 mM K+ bath solution (P < 0.05, unpaired Student's t test). Thus, hypokalemia augmented fentanyl-induced reduction of IhERG in additive (due to chronic hypokalemia) and synergistic (due to acute hypokalemia) manners.
Fentanyl Block Is Modified by Extracellular pH.
Altered extracellular pH affects hERG channel properties (Anumonwo et al., 1999; Terai et al., 2000; Van Slyke et al., 2012; Shi et al., 2019) and drug-channel interactions (Zhang et al., 1999; Wang et al., 2016). In addition, like most hERG channel blockers, such as verapamil (Zhang et al., 1999), fentanyl binds to the internal cavity of the channel from the intracellular side of the membrane (Tschirhart et al., 2019). Fentanyl has a pKa of 8.12, thus it is mostly positively charged at pH 7.4 (Thurlkill et al., 2005). Since the neutral form but not the charged form can freely cross the cell membrane, alterations in pH would change the proportion of fentanyl molecules in the neutral form and thus affect accessibility of fentanyl to the binding sites to achieve block.
Our results showed that hERG channels were blocked with a lesser potency at pH 6.4 and a greater potency at pH 8.4 compared with pH 7.4 (Fig. 3, A and B). The IC50 values were 3220 ± 314 nM with a Hill coefficient of 0.8 ± 0.1 for pH 6.4, 832 ± 53 nM with a Hill coefficient of 0.9 ± 0.1 for pH 7.4, and 127 ± 4 nM with Hill coefficient of 0.8 ± 0.1 for pH 8.4. Our results further showed that the reversibility of fentanyl block was affected by pH. At pH 7.4, 3 μM fentanyl blocked IhERG by 74% ± 5%. After 2 minutes of washout, current recovered to 86% ± 7% of control (Fig. 3C). At pH 8.4, 3 μM decreased IhERG by 90% ± 4% (P < 0.01 compared with pH 7.4). However, the block was only partially reversible. After 2 minutes of washout, the current recovered and stabilized at 50% ± 12% of the control current (P < 0.01 compared with pH 7.4 washout) (Fig. 3C). The delayed current recovery upon washout is not due to the extent of block; when the solution was subsequently changed to pH 7.4, IhERG recovered to near control levels (unpublished data).
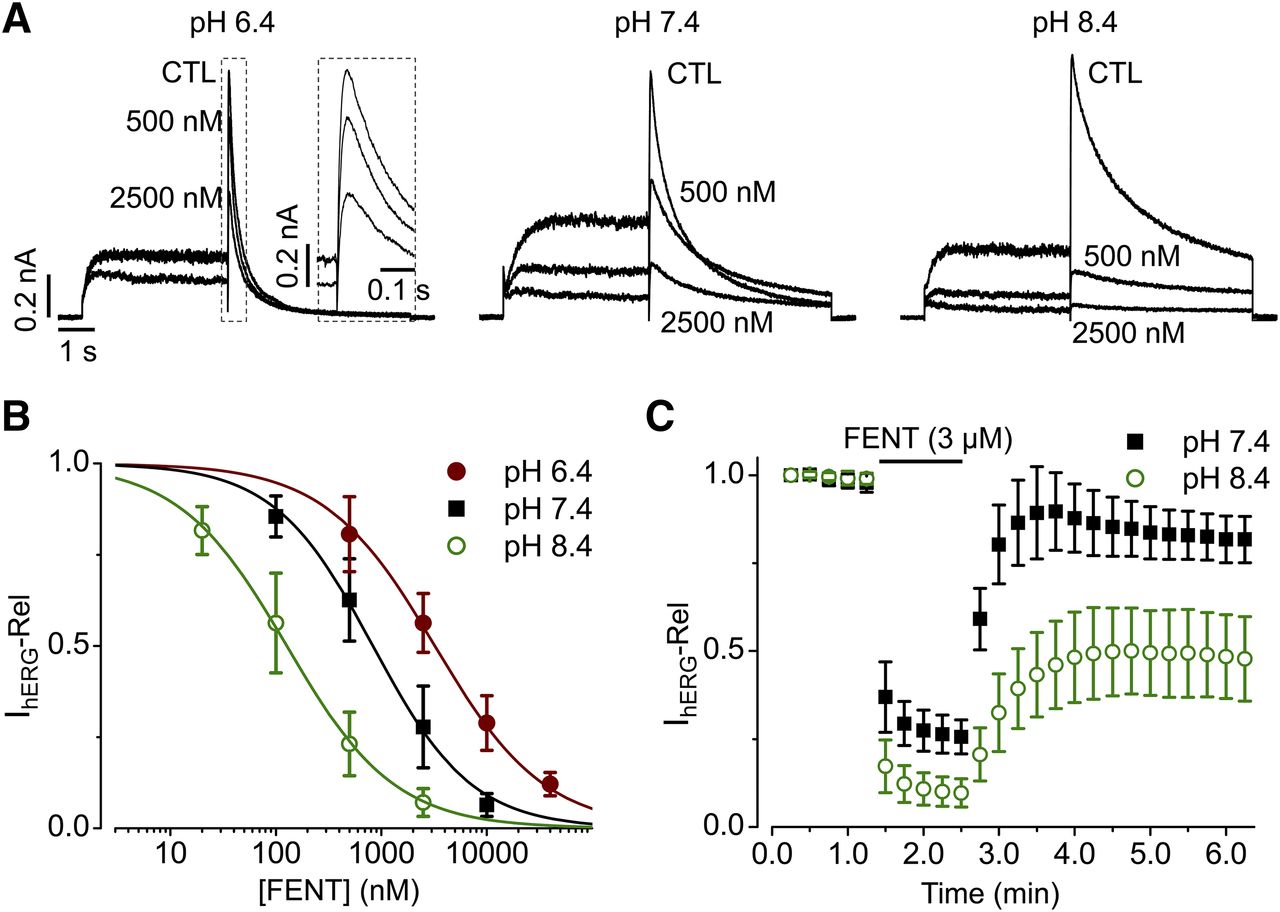
Effects of extracellular pH on fentanyl (FENT)-mediated block of hERG channels. (A) Representative hERG currents recorded in extracellular solution with a pH of 6.4, 7.4, or 8.4 in the absence (CTL) or presence of 500 or 2500 nM fentanyl. The voltage protocol is the same as shown in Fig. 2B. (B) Concentration-response relationships for fentanyl-mediated block of IhERG. Peak tail currents upon −50-mV repolarization at each concentration of fentanyl were normalized to CTL and plotted as relative IhERG (IhERG-Rel) against fentanyl concentrations. The data were fitted to the Hill equation to determine IC50 values. pH 6.4, n = 5; pH 7.4, n = 7; and pH 8.4, n = 5, from three independent experiments. (C) IhERG recorded during control, 3 μM of fentanyl, and upon washout in bath solutions with pH of 7.4 or 8.4. The voltage protocol is the same as shown in Fig. 2B with a start-to-start interval of 15 seconds. IhERG from each cell upon repetitive pulses was normalized to that upon the first pulse and summarized. pH 7.4, n = 7, and pH 8.4, n = 8, from three independent experiments. Error bars represent S.D.
hERG Drug-Binding Site Mutant Channels Display Altered pH Dependence Compared with WT Channels.
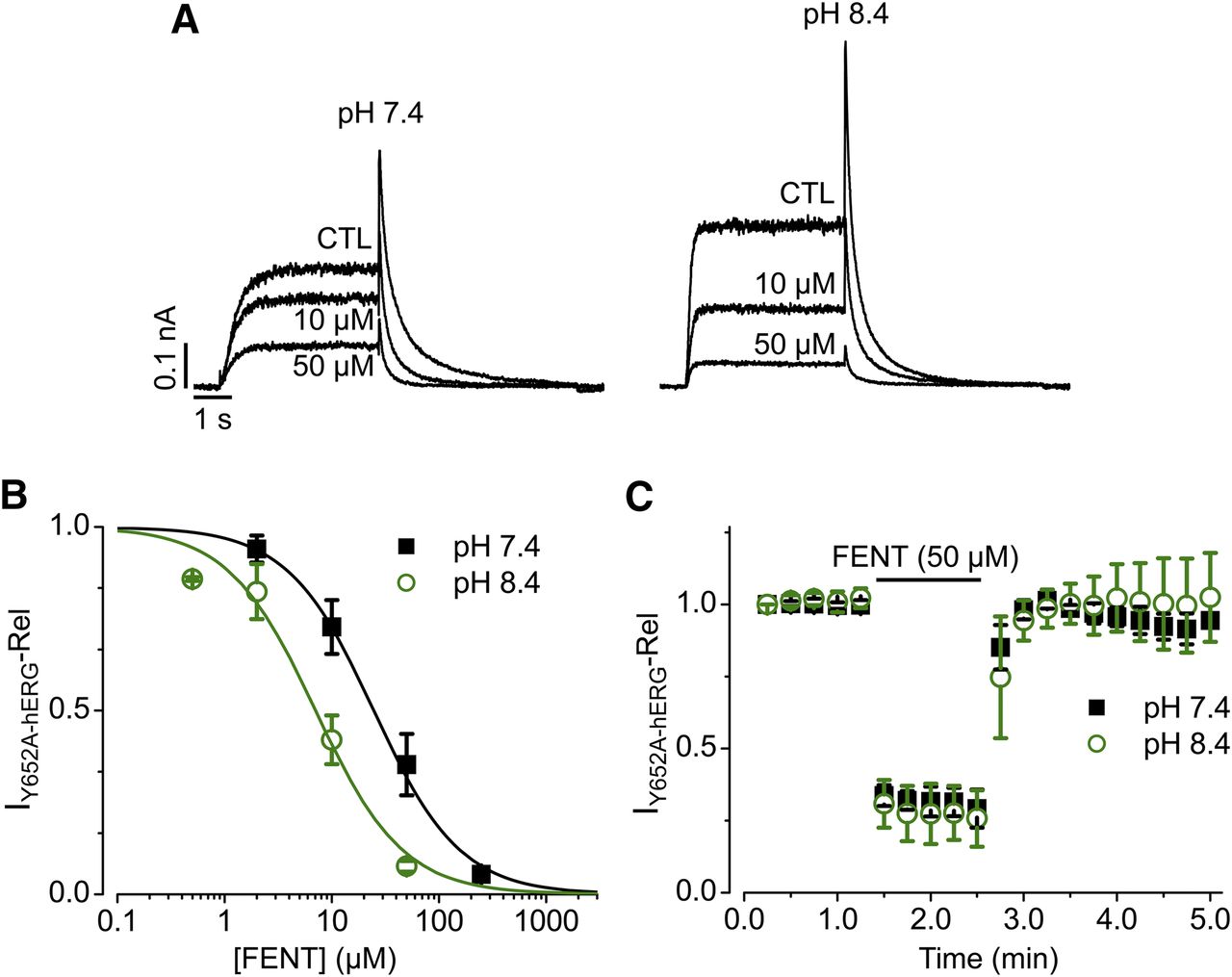
We previously showed that two aromatic residues Tyr652 and Phe656 are involved in fentanyl block of hERG channels (Tschirhart et al., 2019). Our results indicate that these residues also played a role in high pH-potentiated channel block. As shown in Fig. 4, A and B, Y652A hERG channels were also blocked with a greater potency at extracellular pH 8.4. The IC50 values were 24.7 ± 1.5 μM with a Hill coefficient of 1.1 ± 0.1 for pH 7.4 and 7.1 ± 1.0 μM with a Hill coefficient of 1.1 ± 0.2 for pH 8.4. However, in contrast to WT hERG channels, IY652A-hERG completely recovered upon fentanyl washout. At pH 7.4, 50 μM fentanyl decreased IY652A-hERG by 70% ± 7%. After 2 minutes of washout, current recovered to 93% ± 3% of control (Fig. 4C). At pH 8.4, 50 μM decreased IY652A-hERG by 75% ± 10% (P > 0.05 compared with pH 7.4). After 2 minutes of washout, current recovered to 103% ± 14% compared with control current (P > 0.05 compared with pH 7.4 washout).

Effects of extracellular pH on fentanyl (FENT)-mediated block of Y652A hERG channels. Representative Y652A hERG currents recorded in extracellular solution with a pH of 7.4 or 8.4 in the absence (CTL) or presence of 10 or 50 µM fentanyl. The voltage protocol is the same as shown in Fig. 2B. (B) Concentration-response relationships for fentanyl-mediated block of IY652A-hERG. Peak tail currents upon −50-mV repolarization at each concentration of fentanyl were normalized to CTL and plotted against fentanyl concentrations. The data were fitted to the Hill equation to determine IC50 values. pH 7.4, n = 7, and pH 8.4, n = 5, from three independent experiments. (C) IY652A-hERG recorded during control, 50 μM of fentanyl, and upon washout in bath solutions with pH of 7.4 or 8.4. The voltage protocol is the same as shown in Fig. 2B with a start-to-start interval of 15 seconds. IY652A-hERG upon repetitive pulses from each cell was normalized to CTL (IY652A-hERG upon the first pulse). pH 7.4, n = 3, from two independent experiments; pH 8.4, n = 6, from three independent experiments. Error bars represent S.D. Rel, relative.
Deletion of the hERG N Terminus Enhances Fentanyl Block Potency.
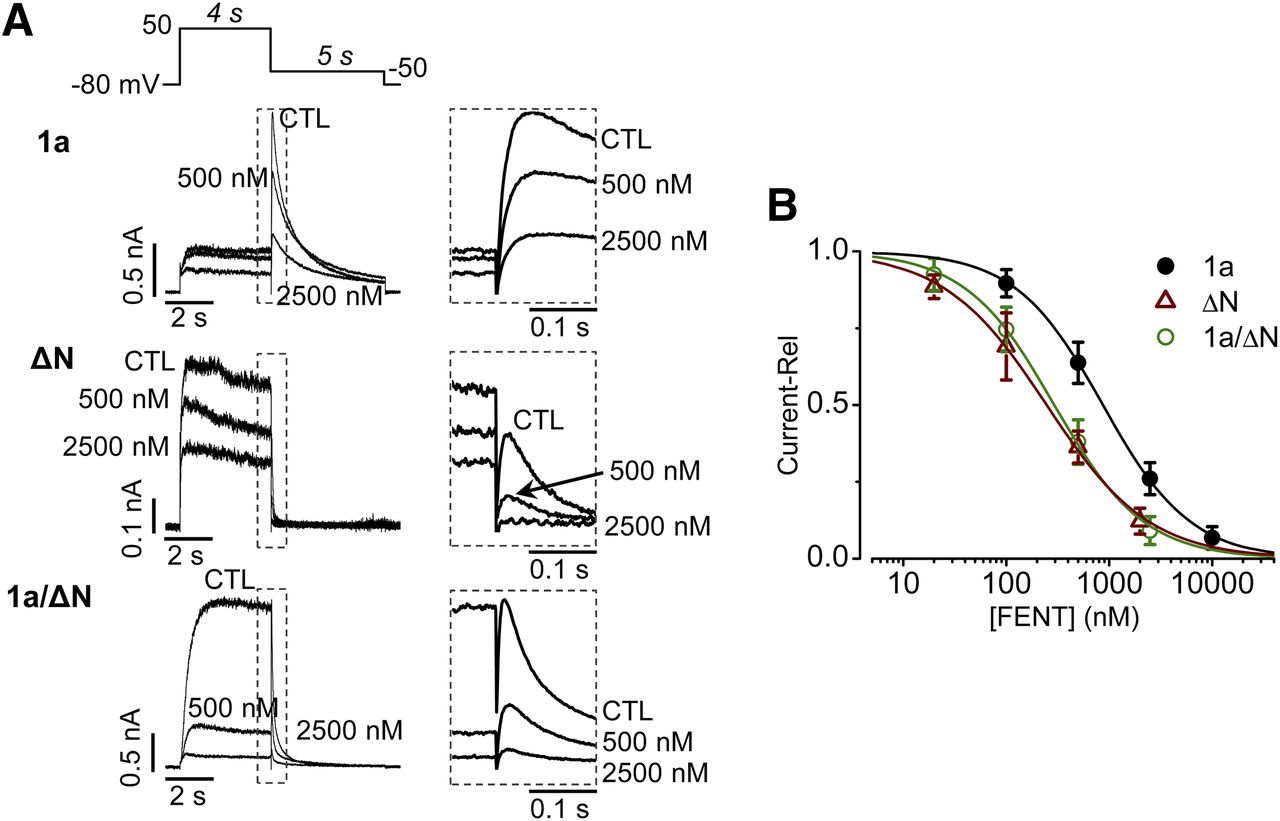
Because there is evidence suggesting that native hERG channels are composed of 1a and 1b isoforms (Lees-Miller et al., 1997; London et al., 1997; Jones et al., 2004, 2014; Phartiyal et al., 2007), we were interested in examining whether these channels are blocked by fentanyl with a different potency than the homotetrameric 1a channels (hERG) previously studied. Since hERG1b alone does not produce current (Phartiyal et al., 2008), we examined fentanyl block of homotetrameric Δ2–354 hERG channels stably expressed in HEK cells (Δ2–354 hERG-HEK cells). These channels resemble hERG1b because the 1b isoform lacks the first 376 amino acids of 1a (Lees-Miller et al., 1997; London et al., 1997). Δ2–354 hERG channels were blocked with a greater potency than WT hERG channels (Fig. 5). The IC50 values were 869 ± 17 nM (Hill coefficient of 1.0 ± 0.1) for WT hERG, and 247 ± 10 nM (Hill coefficient of 0.9 ± 0.1) for Δ2–354 hERG channels (P < 0.01).

Block of N-terminal deletion hERG mutant by fentanyl. (A) Voltage protocol and representative currents recorded from WT hERG1a-HEK cells (1a), Δ2–354 hERG-HEK cells (ΔN), and HEK cells expressing hERG1a and Δ2–354 hERG (1a/ΔN) in the absence (CTL) or presence of 500 or 2500 nM of fentanyl. The start-to-start interval was 15 seconds. Peak tail currents upon −50-mV repolarization were used for analysis. (B) Concentration-response relationships for fentanyl-mediated block of hERG1a (1a), Δ2–354 hERG (ΔN), and hERG1a/Δ2–354 hERG (1a/ΔN) currents. The currents in the presence of each concentration of fentanyl were normalized relative to CTL and plotted as relative current (Current-Rel) against fentanyl concentrations. Data were fitted to the Hill equation to determine IC50 values. hERG1a, n = 12, from five independent experiments; Δ2–354 hERG, n = 12, from three independent experiments; hERG1a/Δ2–354, n = 12, from three independent experiments. Error bars represent S.D.
To establish the physiologic significance of this increased block potency, we transiently transfected Δ2–354 hERG into hERG-HEK cells to produce hERG1a/Δ2–354 hERG channels to resemble native hERG1a/1b channels. Interestingly, these channels were also more sensitive to fentanyl than hERG1a (Fig. 5). The IC50 was 294 ± 10 nM (Hill coefficient of 1.0 ± 0.1).
Heterotetrameric hERG1a/1b Channels Are Blocked by Fentanyl with a Greater Potency.
To ensure that the enhanced block observed with Δ2–354 hERG channels also applied to the physiologic hERG1a/1b tetramers, we examined fentanyl block of hERG1a/1b channels expressed in HEK cells. hERG1a/1b channels were blocked with a greater potency than hERG1a (generally referred to as hERG) (Fig. 6). The IC50 values were 862 ± 28 nM (Hill coefficient of 1.0 ± 0.1) for hERG1a and 332 ± 5 nM (Hill coefficient of 1.0 ± 0.1) for hERG1a/1b.

Block of hERG1a/1b currents by fentanyl (FENT). Representative hERG1a and hERG1a/1b currents in the absence (CTL) or presence of 500 or 2500 nM fentanyl. The voltage protocol was the same as in Fig. 5A. Peak tail currents upon −50-mV repolarization at each concentration of fentanyl were normalized to CTL and plotted against fentanyl concentrations. Data were fitted to the Hill equation to determine IC50 values. hERG1a, n = 7, from three independent experiments; hERG1a/1b, n = 6, from two independent experiments. Rel, relative.
Fentanyl Block Potency Correlates with Channel Deactivation Time Constants.
Since fentanyl is trapped by channel closure (Tschirhart et al., 2019), we posited that accelerated deactivation may contribute to the increased sensitivity of Δ2–354 and hERG1a/1b channels to fentanyl block. To this end, we examined Δ2–9 and Δ2–25 hERG channels transiently expressed in HEK cells. Like Δ2–354 hERG, Δ2–9 and Δ2–25 hERG mutants also display accelerated deactivation (Ng et al., 2011). Since smaller deletions may have less impact on channel structure, they would therefore allow us to investigate the role of accelerated deactivation more directly. Our results showed that Δ2–9 hERG channels were blocked with a greater potency than WT hERG, with an IC50 of 386 ± 10 nM and a Hill coefficient of 1.1 ± 0.1 (Fig. 7A). As well, Δ2–25 hERG current was blocked with an IC50 of 203 ± 2 nM and a Hill coefficient of 1.0 ± 0.1 (Fig. 7B). We then plotted the IC50 values for each cell in all channel types (hERG1a, Δ2–9, Δ2–25, Δ2–354, hERG1a/Δ2–354, and hERG1a/1b studied in Figs. 5⇑–7) against the corresponding fast (τf-deact, Fig. 7C) and slow deactivation time constants (τs-deact, Fig. 7D) of tail currents upon −50-mV repolarizing step. There was a correlation between each deactivation time constant and IC50. The correlation coefficient (r) determined by linear regression analysis was 0.76 for τf-deact (P < 0.01) and 0.68 for τs-deact (P < 0.01). These data indicate that accelerated deactivation increases the block of hERG by fentanyl.

Higher fentanyl (FENT) block potency is associated with accelerated deactivation of hERG channels. Representative Δ2–9 hERG (A) and Δ2–25 hERG (B) currents in the absence (CTL) or presence of 500 or 2500 nM fentanyl. The voltage protocol was the same as in Fig. 5A. Tail currents upon −50-mV repolarization were used for analysis. The concentration-response curves are shown beneath the currents for Δ2–9 hERG (A) and Δ2–25 hERG (B), respectively. Data were fitted to the Hill equation to determine IC50 values. For Δ2–9 hERG, n = 5 from two independent experiments; for Δ2–25 hERG, n = 8 from three independent experiments. Error bars represent S.D. (C and D) IC50 values plotted against the respective fast [τf-deact, (C)] and slow time constant of deactivation [τs-deact, (D)] from data obtained in (A and B), as well as in Figs. 5 and 6. The plotted data were fitted with linear regression analysis. For Δ2–9 hERG, n = 5, from two independent experiments; Δ2–25 hERG, n = 8, from three independent experiments; Δ2–354 hERG, n = 12, from three independent experiments; WT hERG1a/Δ2–354 hERG, n = 12, from three independent experiments; hERG1a/1b, n = 6, from two independent experiments; WT hERG1a, n = 19, from seven independent experiments. Rel, relative.
Hypoxia, Hypokalemia, and pH Affect Fentanyl Block of hERG 1a/1b Channels in a Manner Similar to hERG1a.
To investigate whether hERG 1a/1b channels respond to hypoxia, hypokalemia, and pH along with fentanyl in a manner similar to hERG1a channels, we cultured hERG 1a/1b–expressing HEK cells in normoxic (CTL) or hypoxic (0.5% O2) conditions for 6 hours and recorded currents in the absence and presence of 0.5 µM fentanyl. Compared with control, 6-hour culture in 0.5% O2 or acute application of 0.5 μM fentanyl significantly decreased IhERG1a/1b. The combination of hypoxic culture and 0.5 μM fentanyl resulted in IhERG1a/1b that was significantly less than either treatment alone (Fig. 8A).

Effects of chronic hypoxia (Hx), hypokalemia, and external pH on fentanyl-induced block of IhERG1a/1b. (A) hERG1a/1b-HEK cells were cultured in normoxia (CTL, 21% O2) or hypoxia (Hx, 0.5% O2) for 6 hours. For each cell, IhERG1a/1b was recorded in the absence or presence of 0.5 μM fentanyl (FENT). Representative current traces elicited with the voltage protocol shown in Fig. 1 are shown above scatter plots of data from each condition. CTL, n = 14, and FENT, n = 14, from four independent experiments; Hx, n = 13, and Hx + FENT, n = 13, from three independent experiments. (B) hERG1a/1b-HEK cells were cultured in normal (5 mM K+, CTL) or 2 mM K+ (2K+) medium for 72 hours. IhERG1a/1b was then recorded in the absence or presence of 0.5 μM fentanyl in standard bath solution. Representative currents and scatter plot of IhERG1a/1b amplitudes at −50-mV repolarization after 50-mV depolarization are shown. CTL, n = 11; FENT, n = 11; 2K+, n = 16; 2K++FENT, n = 16 from three independent experiments. (C). Representative hERG1a/1b currents and scatter plots of the current amplitudes recorded in bath solutions with a pH of 6.4, 7.4, or 8.4 in the absence (CTL) or presence of 0.5 µM fentanyl. pH 6.4, n = 7, from two independent experiments; pH 7.4, n = 7, from three independent experiments; pH 8.4, n = 7, from two independent experiments. In (A–C), *P < 0.05; **P < 0.01, one-way ANOVA with Tukey’s post hoc test.
To study the effects of hypokalemia and fentanyl on IhERG1a/1b, we cultured channel-expressing HEK cells in normal medium (CTL, 5 mM K+) or 2 mM K+ for 72 hours. After culture, we examined the blocking effects of fentanyl on IhERG1a/1b recorded in standard, 5-mM K+-containing bath solution in the absence or presence of 0.5 μM fentanyl. Compared with CTL, culture in 2 mM K+ medium for 72 hours or acute application of 0.5 μM fentanyl significantly decreased IhERG1a/1b. The combination of 2-mM K+ culture and 0.5 μM fentanyl resulted in IhERG1a/1b that was significantly less than either condition alone (Fig. 8B).
To study the effects of pH on fentanyl-induced block of IhERG1a/1b, we examined 0.5-μM fentanyl-mediated block of currents recorded in standard bath solutions with pH 6.4, 7.4, or 8.4. IhERG1a/1b was blocked with a lesser potency at pH 6.4 and a greater potency at pH 8.4 compared with pH 7.4 (P < 0.01 for each, one-way ANOVA with Tukey’s post hoc test). Although 0.5 μM fentanyl blocked IhERG1a/1b by 58% ± 13% at pH 7.4, it blocked IhERG1a/1b by 19% ± 10% at pH 6.4 and by 84% ± 4% at pH 8.4 (Fig. 8C).
Discussion
Fentanyl-related death is a serious issue in North America (Alexander et al., 2016; Torralva and Janowsky, 2019). After finding that fentanyl blocks the hERG potassium channel and reduces IKr (Tschirhart et al., 2019), the present work examined conditions that may interact with or exacerbate fentanyl-induced channel block. First, chronic hypoxia reduces mature hERG density and IhERG (Lamothe et al., 2017). We found that chronic hypoxia and acute fentanyl reduced IhERG in an additive manner (Fig. 1). Since fentanyl-mediated respiratory depression (McQueen, 1983) may result in hypoxia, the additive impact of fentanyl-induced channel block and hypoxia-associated channel density reduction would further impair the function of hERG channels.
Fentanyl abusers may experience conditions, such as diarrhea, vomiting, or malnourishment, that can lead to hypokalemia (a reduced serum K+ concentration), which decreases cell-surface hERG expression (Guo et al., 2009; Massaeli et al., 2010) and causes LQTS (Curry et al., 1976; Roden et al., 1986). Indeed, our results showed that chronic hypokalemia and acute fentanyl additively reduced IhERG (Fig. 2A). In addition, we found that hERG channels were more sensitive to fentanyl when IhERG was recorded in 2-mM K+-containing bath solution compared with fentanyl-induced block of IhERG recorded in standard 5-mM K+-containing bath solution (Fig. 2B). These findings are consistent with previous reports that reduced extracellular K+ concentration potentiates drug-mediated block of hERG channels (Yang and Roden, 1996; Wang et al., 1997; Lin et al., 2007) and drug-induced LQTS (Zehender et al., 1991; Wenzel-Seifert et al., 2011). Thus, low K+ levels (hypokalemia) may decrease IhERG through chronically decreasing hERG protein expression as well as acutely potentiating fentanyl-induced block.
Drug abuse can cause electrolyte and acid-base disturbances. In particular, hypokalemia is often associated with metabolic alkalosis. Our results demonstrate that hERG channels were blocked with 3.9-fold less potency at extracellular pH 6.4 (Fig. 3, A and B) and 6.6-fold greater potency at extracellular pH 8.4 compared with pH 7.4 (Fig. 3, A and B). There are a couple of potential explanations for this phenomenon. First, high pH may increase the membrane permeability of fentanyl. Fentanyl has a pKa of 8.12 (Thurlkill et al., 2005) and thus is predominately positively charged at a pH of 7.4. At a pH of 8.4, the portion of fentanyl in the neutral form would be more than half. Since the neutral form of fentanyl may more readily cross the cell membrane to block hERG channels from the internal side of the cell (Tschirhart et al., 2019), enhanced membrane permeability would be expected to increase block potency.
Another possible explanation for the extracellular pH dependence of fentanyl block is that the neutral form of fentanyl has a higher affinity for the hERG channel than the positively charged form. This possibility has merit because fentanyl interacts with hydrophobic residues lining the pore of the channel (Tschirhart et al., 2019). Indeed, at pH 8.4, fentanyl-mediated block of WT channels was only partially reversed upon washout of the drug (Fig. 3C); however, fentanyl-mediated block of Y652A channels was completely reversed upon washout of the drug (Fig. 4C). Therefore, high-affinity binding to aromatic residues Tyr652 and Phe656 may explain the enhanced potency of fentanyl at increased extracellular pH. A potential flaw in this theory is the pipette solution remained at pH 7.2 for these experiments; therefore, once inside the cell, the majority of fentanyl would be positively charged. However, the pore of hERG is open to the extracellular side of the membrane (Wang and MacKinnon, 2017). Thus, it is possible that the high pH of the extracellular solution is conferred to the pore of the channel. Once fentanyl molecules are in the pore, they may become un-ionized and bind to aromatic residues with the proposed high affinity. This scenario is supported by evidence suggesting that the pore of hERG can become blocked by protons at lower pH values (Bett and Rasmusson, 2003; Van Slyke et al., 2012). The alteration of pH within the channel pore presents another possibility. Such a change in pH may alter channel structure and ionization state of residues, which could create a higher-affinity binding site for fentanyl. Although future studies would be required to elucidate the molecular mechanisms for pH dependence of fentanyl-mediated block, our finding that alkalosis increases the block of hERG by fentanyl has clinical implications. Conditions that cause alkalosis, such as repetitive vomiting, dehydration, or endocrine disorders, may increase the risk of fentanyl-associated arrhythmias. However, it should be noted that in the context of fentanyl-induced respiratory depression, hypercapnia and resulting acidosis (Agrafiotis et al., 2015) may decrease hERG block potency. Nonetheless, alterations in drug-block potency during acid-base disturbances should be considered when assessing the cardiac safety of drugs.
Two transcript variants of hERG exist, hERG1a and the shorter hERG1b (Lees-Miller et al., 1997; London et al., 1997). Early investigations of native hERG channels from adult rat, mouse, and human heart tissues only resulted in the detection of hERG1a protein, and therefore it was proposed that hERG1b does not contribute to cardiac IKr (Pond et al., 2000). However, further research has detected hERG1b proteins in rat, canine, human (Jones et al., 2004), and equine (Finley et al., 2002) heart tissue, suggesting that native IKr contains hERG1a and hERG1b isoforms. Furthermore, electrophysiological evidence suggests the presence of hERG1b in native channels. First, when hERG1a and hERG1b are coexpressed in heterologous expression systems, the currents resemble IKr more closely than hERG1a alone (London et al., 1997; Sale et al., 2008; Larsen and Olesen, 2010). Second, the functional role of hERG1b was elucidated using human-induced pluripotent stem-cell cardiomyocytes, in which knockdown of hERG1b with short hairpin RNA decreases IKr, prolongs APD, and increases APD variability (Jones et al., 2014). Third, mutations in hERG1b have been linked to LQTS in patients (Sale et al., 2008).
To investigate fentanyl blockade of hERG1a/1b channels, we first examined fentanyl block of homotetrameric Δ2–354 hERG channels as an alternative to hERG1b since hERG1b does not produce current when expressed alone in HEK cells (unpublished data). Δ2–354 hERG channels resemble hERG1b because the 1b isoform lacks the first 376 amino acids of 1a (Lees-Miller et al., 1997; London et al., 1997). Our results show that Δ2–354 hERG channels were blocked by fentanyl with a 3.5-fold greater potency than WT (hERG1a) channels (Fig. 5). When Δ2–354 hERG was expressed in hERG-HEK cells, currents were blocked with a 3.0-fold greater potency than WT channels (Fig. 5).
After this proof of concept, we obtained hERG1b plasmid as well as a stable hERG1a/1b cell line from Dr. Gail Robertson at University of Wisconsin - Madison. We also transfected hERG1b plasmid to hERG1a-HEK cells. Our results showed that heterotetrameric hERG1a/1b channels were blocked to a similar degree as hERG1a/Δ2–354 hERG channels with a 2.6-fold greater potency than homotetrameric hERG1a channels (Fig. 6). Since hERG1a and hERG1b share an amino acid sequence for the S1 segment to the C terminus, including the pore region, the altered block potency may be due to changes in channel gating. One feature of hERG1a/1b channels is accelerated deactivation gating compared with hERG1a channels (Sale et al., 2008). We previously demonstrated that fentanyl is an open-channel blocker and is trapped by channel closure (Tschirhart et al., 2019). Furthermore, fentanyl block of hERG is strongly voltage-dependent, being greater at positive voltages and weaker at negative voltages. Thus, following block during a depolarizing step, a certain degree of unblock occurs upon repolarizing steps (e.g., −50 mV) before channels deactivate, which then traps fentanyl molecules that occupy the pore within the closed channels (Tschirhart et al., 2019). Thus, accelerated deactivation would result in increased trapping of drugs, leading to enhanced block. This notion is supported by our results obtained in several hERG N-terminal deletion mutants, all of which demonstrated accelerated deactivation and enhanced sensitivity to fentanyl (Figs. 5⇑–7). Furthermore, plotting the IC50 for fentanyl block versus τf-deact and τs-deact resulted in a statistically significant correlation between deactivation time constants and block potency (Fig. 7, C and D). In addition to accelerated deactivation, due to faster activation and recovery from inactivation (Sale et al., 2008), hERG1a/1b channels occupy the open state for longer than hERG1a channels. Thus, the possibility that a greater affinity for the open state of channels contributes to the greater sensitivity of hERG1a/1b channels to fentanyl cannot be excluded (Sale et al., 2008; Abi-Gerges et al., 2011).
Finally, our results show that the responses of hERG1a/1b channels to hypoxia, hypokalemia, and pH along with fentanyl are similar to hERG (hERG1a) channels (Fig. 8).
In summary, we have uncovered several circumstances in which the block of hERG by fentanyl may become more significant. First, hERG1a/1b channels are more susceptible to block, raising the possibility that fentanyl block may be more clinically significant than previously thought since both hERG1a and hERG1b are present in the human heart. In addition, conditions such as hypoxia, hypokalemia, and alkalosis can exacerbate fentanyl-mediated impairment of hERG current, creating situations in which arrhythmias due to hERG blockade may underlie fentanyl-related sudden death.
Acknowledgments
We acknowledge Jun Guo and Wentao Li for assistance with cell culture and some patch-clamp experiments. We thank Dr. Gail Robertson (University of Wisconsin – Madison) for providing the hERG1a/1b stable cell line as well as hERG1a and hERG1b plasmids.
Authorship Contributions
Participated in research design: Tschirhart, Zhang.
Conducted experiments: Tschirhart.
Performed data analysis: Tschirhart.
Wrote or contributed to the writing of the manuscript: Tschirhart, Zhang.
Footnotes
- Received December 23, 2019.
- Accepted April 8, 2020.
This work was supported by the Canadian Institutes of Health Research [MOP 72911] and the Heart and Stroke Foundation of Canada [G-17-0018754] to S.Z.
The authors declare that they have no conflicts of interest with the contents of this article.
Abbreviations
- APD
- action potential duration
- CTL
- control
- τf-deact
- fast time constant of hERG deactivation
- HEK
- human embryonic kidney
- hERG
- human ether a-go-go–related gene
- IhERG
- hERG current
- IKr
- rapidly activating delayed rectifier potassium current
- LQTS
- long QT syndrome
- MEM
- minimum essential medium
- τs-deact
- slow time constant of hERG deactivation
- WT
- wild type
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}