
Abstract
Camera-trap studies in the wild record true-positive data, but data loss from false-negatives (i.e. an animal is present but not recorded) is likely to vary and widely impact data quality. Detection probability is defined as the probability of recording an animal if present in the study area. We propose a framework of sequential processes within detection – a pass, trigger, image registration, and images being of sufficient quality. Using closed-circuit television (CCTV) combined with camera-trap arrays we quantified variation in, and drivers of, these processes for three medium-sized mammal species. We also compared trigger success of wet and dry otter Lutra lutra, as an example of a semiaquatic species. Data loss from failed trigger, failed registration and poor capture quality varied between species, camera-trap model and settings, and were affected by different environmental and animal variables. Distance had a negative effect on trigger probability and a positive effect on registration probability. Faster animals had both reduced trigger and registration probabilities. Close passes (1 m) frequently did not generate triggers, resulting in over 20% data loss for all species. Our results, linked to the framework describing processes, can inform study design to minimize or account for data loss during analysis and interpretation.
Similar content being viewed by others

Introduction
Camera-traps (CTs) are used for a range of ecological studies from determining presence or occupancy (Mugerwa et al. 2013; Tobler et al. 2015) to activity (Lim and Ng 2008). Studies using CTs have proliferated; however, it is not considered ‘fully mature as a methodological discipline’ (Rowcliffe 2017). The technical aspects of how CTs using passive infrared (PIR) motion detectors function and clarification of associated terminology have been described (Welbourne et al. 2016). In short, a specialized ‘Fresnel’ lens focuses background infrared radiation (IR), filtered to 8–14 μm onto a pyroelectric sensor. This sensor detects rapid changes in background IR which triggers the camera to record. As with more traditional census techniques, it is recognized that PIR CTs are prone to false-negatives, i.e. fail to detect a species which is present (Gužvica et al. 2014). Detection probability is a fundamental issue in CT studies of occupation and population density, particularly in studies using random encounter modelling (REM) of animals that lack easily distinguishable individual markings (Rowcliffe et al. 2008).
Field data from CTs can only include true-positives: when an animal pass elicits a trigger which results in registration of the animal as recorded footage. In order to achieve a true-positive, a number of sequential processes have to occur, all of which must have a successful outcome (Fig. 1), and these sequential processes underlie a series of measurable conditional probabilities. False positives, such as misidentification of species, sex or individual, are errors by the observer of the footage and not the CT itself. Some species may be more prone to being incorrectly identified, such as Scottish wildcat Felis silvestris silvestris, where the phenotype of the ‘pure’ species and the hybrid are very similar. True negatives are the result of an absence of footage in an area where a species is absent. False-negatives can arise from failure of any processes in Fig. 1. True and false-negatives cannot be distinguished from each other which is why it is important to try to understand and account for the latter.

The sequential processes required to detect an animal on a camera-trap given that it is present. Failure of any of these processes leads to a false-negative; therefore, detection success requires a positive outcome from all the component processes. Specific terminology we use in this study to quantify these processes is also shown. ‘Detection probability’ can thus be considered the product of a series of conditional probabilities representing each of these processes
Process 1: Encounter probability P(pass|presence). This is the probability an animal will pass through the putative ‘detection zone’ of a CT given that it is present in the study area. This has been demonstrated to be affected by aspects of survey design such as the density and placement of CTs in relation to the species rarity and home-range size (O’Connor et al. 2017), sampling effort, specifically number of CT days and number of CTs deployed (Tobler et al. 2008), use of attractants such as bait (Hamel et al. 2013) and animal reaction to CT presence (Larrucea et al. 2007). Inappropriate sampling design could affect the probability of a pass, for instance, setting the CT at ground level for arboreal species.
Process 2: Trigger probability P(trigger|pass). This is the probability that the CT’s PIR sensor senses a change in infrared from the pass of an animal which causes the CT to trigger. It has been suggested that mammals with aquatic lifestyles result in low trigger probability as their thermal footprint can be compromised by wet fur after exiting water (Lerone et al. 2015).
Process 3: Registration probability P(registration|trigger). A CT trigger is not sufficient alone to record an animal – the animal must also be visible on the CT image or video. Trigger latency or trigger speed is the interval of time between PIR trigger and initiation of the camera (Rovero et al. 2013) which can vary widely between CT models (Randler and Kalb 2018). A slow trigger speed coupled with fast moving animals means that not all triggers lead to registration as the animal has passed through the field-of-view before the camera has been activated (Rovero et al. 2013). The field-of-view of the camera is not necessarily the same width as the detection zone monitored by the PIR motion detector (Rovero et al. 2013; Trolliet et al. 2014; Rovero and Zimmermann 2016), thus affecting registration probability. Previous studies, without use of a control (to identify scenarios where an animal triggers the camera but is not recorded) have only been able to measure the combined detection of processes 2 and 3 (Rowcliffe et al. 2011; Hofmeester et al. 2017). So whilst body mass, season and relative position of an animal with respect to the camera are likely to influence across processes 2 and 3 (Rowcliffe et al. 2011), these may operate on trigger probability, registration probability or both.
Process 4: Capture quality probability P(capture quality|registration). Not all footage/images of a study species are of equal value, as images of a given quality may be required depending on a study’s objectives. ‘Quality’ here refers to the contents of the footage/images rather than image resolution per se. For example, if aiming to identify individuals, reliable unique markers need to be visible, so a given angle of view or fully body image may be required (Foster and Harmsen 2012). Similarly, in species where it is possible to determine sex, and the study aims require this, footage containing sufficient views of an animal in terms of primary and/or secondary sexual characteristics may be required (Findlay et al. 2017), and whilst video may be better than stills, sexing animals may not be possible for every registration.
Hofmeester et al. (2019) developed a conceptual framework for detectability in CT studies which considers animal characteristics, CT specifications, CT set-up protocols and environmental variables in context with a hierarchy of different spatial scales and six orders of habitat selection. Our framework broadly converges with this. In practice, most CT studies cannot quantify trigger probability in isolation from registration probability, and often trigger probability is misrepresented as a combination of trigger and registration together. Using closed-circuit television (CCTV), we look specifically at Processes 2–3 (Fig. 1), which equate to the 5th and 6th scale described by Hofmeester et al. (2019), i.e. what happens when an animal passes in front of a CT, and we also present capture quality probability as a separate process.
We hypothesize that different environmental and animal-based factors will bias/influence each process as they result from different functional components of the CT (the PIR sensor and the camera). For example, trigger probability will relate to changes in IR received by the PIR sensor, and the PIR sensitivity setting. This received IR will in turn will be governed by the spatial relationship between the animal and the PIR sensor as the animal enters the putative zone of detection, as well as the thermal properties of the animal’s surface in relation to the background, CT height and vegetation density (see Hofmeester et al. 2019). Registration probability only applies when the PIR sensor has triggered and will be governed by the spatio-temporal relationship between the animal and the camera’s field-of-view in the time between the trigger and camera initiation (i.e. the trigger speed), and may also be affected by variables such as the speed of the passing animal, and variables with potential to completely obscure the image such as dense vegetation and fog. Capture quality probability may be affected by the proportion, and which portion, of the animal that is within the image, in addition to factors that may affect the quality of the image, e.g. the speed of the passing animal (blurring), vegetation density (obscuring view), weather (mist and rain) and time of day (glare from sun).
We used CCTV as a control to record all passes of each of our target species through the putative detection zones of arrays of CTs in order to observe at which process CTs produced false-negatives. CCTV explicitly allowed us to observe all passes, even when these did not elicit a trigger or did elicit a trigger but not a registration. Using CCTV enables distinction between the latter and genuine ‘false triggers’ (i.e. triggers caused by extraneous stimuli which also result in footage not containing the target species). Such a distinction cannot be made without a control (e.g. CCTV or direct observation). Two CT models were chosen to contrast field-of-view and detection zone differences, one with a more standard detection zone and field-of-view (Bushnell) and one with wide detection and field-of-view (Acorn). We were able to separately investigate variation in trigger probability, registration probability and elements of capture quality probability for one semiaquatic (Eurasian otter Lutra lutra) and two terrestrial (red fox Vulpes vulpes and Eurasian badger Meles meles) mammal species of a similar size (hereafter ‘otter’, ‘fox’ and ‘badger’). We hypothesised that the variables driving success in processes 2, 3 and 4 would be different, for example, we would expect trigger probability to be influenced primarily by distance, whilst registration probability would be most influenced by movement patterns, such as speed. Furthermore, we hypothesized that trigger probability of wet otters would be lower than that of dry otters (Lerone et al. 2015). We use our findings to suggest key considerations of study design and potential sources of bias in CT studies.
Materials and methods
Data collection
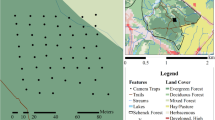
We used two study sites. The first was a wild area in SE Scotland (55.9 °N, 3.2 °W). We targeted a mammal run in woodland known to be used by both badger and fox. The second was a captive otter enclosure (50.6 °N, 4.2 °W) in SW England. The enclosure was approximately 700 m2, with a pond accounting for approximately a third of the area. The enclosure included two wooden hutches for denning, termed ‘holts’. A male and a female otter lived in the enclosure; they were not intended for release and were habituated to humans. In both study areas, we set up two CCTV cameras (Swann SRPRO-842) at approximately 2 m above ground to continuously record to a CCTV recorder (M2/UTC-FDVR-4). The CCTV used IR illumination at night and was able to observe 24 h per day. Both sites had flat topography, and work was undertaken in winter when vegetation would be at minimum density and height (otter: 14 Nov–5 Dec 2017, fox & badger: 21 Feb–14 April 2017). At both sites, we set up four CT stations, subsequently referred to as CT ‘positions’, within the CCTV field-of-view with the PIR at 27 cm above the ground approximating average shoulder height of the three species studied. CTs were aimed parallel to the ground and placed in security boxes so that they could be replaced at the same height and angle.
For both trials, we used Bushnell Aggressor (model 119,776) CTs programmed to record 5 s video with an interval of 5 s between recordings. Video potentially captures more data than still images, and use of video is likely to increase due to technological advances (Swinnen et al. 2014). In the otter enclosure, at each recording station, we also set a Bushnell CT to record a burst of 3 still images with a 5 s interval between bursts and a Little Acorn (model 5310 WA) CT to record 5 s video with a 5 s interval, see Fig. 2. We set Bushnell CTs to ‘auto’ sensitivity as recommended by the manufacturer. The Acorn was set to medium sensitivity. The Acorn was used as a contrast to the Bushnell as its PIR sensor has an advertised 100° detection angle and 100 ° camera field-of-view, compared to an advertised 55° detection angle and 40 ° field-of-view for the Bushnell. At both sites, we fixed a data logger (Onset Hobo) 1.5 m above the ground to record hourly air temperature, and in the otter enclosure pond, we secured a data logger at 30-cm depth to record hourly water temperature.

Schematic maps showing the positions of the camera-trap (CT) arrays and closed-circuit television (CCTV) at the study sites for a badger and fox, and b otter. Scales and relative positions are approximate, and CTs and CCTVs are oversized. Arrows indicate direction CT stations faced
At both sites, we determined distances between each CT and features visible on the CCTV such as habitually used trails and trees in each CTs’ field-of-view. CCTV footage was reviewed to identify passes of a single animal, and we created a chronological list of passes. We defined a ‘pass’ as a single animal moving across the central line of the CT’s field-of-view (see Hofmeester et al. 2017). As CTs targeted mammal runs, virtually all animals passed the central line. We included passes where the target species was considered the only potential stimulus for the CT PIR sensor, so we excluded passes where extraneous stimuli were present, such as birds and rodents. Waving vegetation and direct sunlight would also have been seen as an extraneous stimuli, but these were not an issue during our study period because vegetation was sparse at the time of year of the study, and it was overcast and not windy. We also excluded passes where the animal was less than 1 m from the CT, as the animals could potentially pass beneath the PIR sensor and/or field-of-view (Rowcliffe et al. 2011).
We cross-referenced passes on the CCTV footage against the CT footage using their respective timestamps. This enabled us to separately quantify Processes 2 and 3 (Fig. 1), i.e. distinguishing an animal passing but not triggering the CT from an animal triggering the CT but not registering in its footage. This process eliminated any false triggers (i.e. where a CT triggered but no otter, fox or badger had passed).
Variables recorded
We quantified trigger probability P(trigger|pass) with a binary variable of passes which either triggered the camera (1) or did not (0), regardless of whether its footage registered the animal. We also quantified registration probability P(registration|trigger) with a binary variable of passes which either triggered the camera and registered the animal (1) or triggered the camera but failed to register the animal (0).
As discussed, capture quality probability P(capture quality|registration) depends on a study’s objectives. In many studies of mammals, identifying presence of the species is not necessarily sufficient, but rather a good view of the head and body is needed to identify the age category/sex/breeding status of the individual (for instance, lactating females) (Sollmann and Kelly 2013; Findlay et al. 2017) or to observe individual natural markings (Karanth 1995; Silver et al. 2004). We used capture of the head of the animal in the first video frame or image as an indication of minimum capture quality as more of the animals would normally be captured in the following video footage or images. We quantified capture quality probability with a binary variable categorizing good capture quality as capture of head only, head and body, or head, body and tail (1) or poor capture quality when the head had already passed through the field-of-view (0).
From the CCTV footage and data loggers, a suite of animal and environmental variables were recorded for each pass (Table 1). The orientation of the animal pass to the CT was recorded, using three categories. A lateral pass was when the animal passed exposing a complete side view and an anterior pass was when the animal approached the camera-trap presenting the head, shoulders and front legs and a posterior pass when the animal approached the CT from behind and walked away exposing its hindquarters. We chose to record an animal’s gait (i.e. walk, trot, run) to represent speed as gait was quickly identifiable whilst estimating ms-1 over such short distances would be prone to inaccuracies from perspective using CCTV footage and inconsistencies due to instances of the animal pausing. Running animals were subsequently combined with trotting animals as running animals were too infrequent to analyse separately; our variable GAIT therefore had two categories (walk/trot or run). We recorded whether there was any delay in the animal passing through the field-of-view as a result of the animal pausing to sniff or scent mark (i.e. loitering). This was recorded as a binary variable LOIT. For otter, we also recorded whether the animal was dry after being in the holt and prior to immersion in water (from holt) or whether the animal had been immersed in water since leaving the holt (not from holt). This enabled us to subset the data to include passes where the otter was fully dry or not fully dry. For fox and badger, we only used Bushnell CTs on video setting. For otter, we had stations of three CTs (Bushnell video, Bushnell still images, Acorn video) together to maximize data acquisition from each pass. We analysed data for each of the three CT models/settings separately so we could compare Bushnell video between fox/badger and otter, and because aspects of the three CT models/setting differ substantially in key elements such as detection zone, field-of-view, etc..
To understand how the otters’ IR footprint develops after exiting from water, we used a thermal imager (FLIR PAL65) to take thermal images of otter on dry ground from the point of exiting water to 300 s post-immersion. Seventeen images were taken, the land temperature ranged between 6 and 10 °C and the water was 9.5 °C. Mean temperature of the otter trunk and an equivalent area of ground adjacent to the otter were measured using FLIR Tools software (v5.13.17214.2001). The absolute difference in temperature was plotted against time from water (Fig. 3), and an exponential model was fitted to the data. Approximately a 2.7 °C difference between animals emitted with IR and the background IR is needed for a PIR sensor to initiate a trigger (Meek et al. 2012), although this will depend on the CT model and PIR sensitivity setting. Under these conditions, the fitted model predicts 32 s to have elapsed before the temperature difference reaches a conservative 3 °C.

Absolute difference (ΔABS) in temperature (°C) between an otter’s trunk and surrounding land against time after being immersed in water illustrating how long since immersion it takes for the otter to emit enough heat (c. 3 °C) for a passive infrared sensor to theoretically detect the otter. To describe the asymptotic relationship, we fitted an exponential model in the form y = a(1-e-bx) + c where y is the temperature difference, x is the time since exiting water, and a, b and c are parameters estimated by the model. The absolute difference between air and water temperatures is also plotted, using temperature from data loggers
Modelling trigger and registration probabilities
We carried out modelling in R version 3.2.2 (RCore Team 2015) within R Studio (RStudioTeam 2015), fitting generalized linear mixed models (GLMMs) using lme4 (Bates et al. 2015) and generating model comparison tables using MuMIn (Barton 2016). We used the package manipulate (Allaire 2014) to fit the exponential model in Fig. 3.
We used GLMMs with a binomial distribution to investigate variation in the response variables P (trigger|pass) and P(registration|trigger) for each species and CT model. The CTs positions potentially had different local conditions. Therefore, we set CT position as a categorical random effect and built a list of candidate models (online resource 1) containing combinations of appropriate variables in Table 1, including a null model in each.
Distance to CT and orientation of animal could not be investigated in the same model sets, as the trigger distance could not be measured for anterior passes, i.e. when the animal approaches the CT at 180°, whilst for most posterior passes when the animal walks away at 180° the animal would have to enter the detection zone close to the CT. Distance was prioritized as a variable, and lateral passes approximating 90 ° were selected for analysis unless otherwise stated.
We investigated whether immersion in water negatively affected trigger probability for otter, as suggested by Lerone et al. (2015). First we modelled trigger probability for dry otters after they had emerged from their holts and prior to entering water. This allowed us to compare dry otter to fox and badger. Then, we repeated the model comparison including a generated binary variable WET.DRY to distinguish passes where the otter was fully ‘wet’ (≤ 10 s since exiting water) and passes where the otter was fully ‘dry’ (passes where FROM.HOLT = 1). Finally, using all passes where FROM.HOLT = 0, we repeated the model comparison including TFW to test whether it was a significant variable, but it was not well supported. We tested all GLMMs for overdispersion and used a threshold of ΔAIC ≤ 2 to indicate models with ‘substantial support’ (Burnham and Anderson 2004). For brevity, we only include plots for the best supported model (ΔAIC = 0) in the main text, but other plots of all models with ΔAIC ≤ 2 and parameter estimates for all models are provided in the online supplement.
Quantifying detection in a ‘worst-case scenario’
Poor triggering of CTs by otters after emergence from water (Lerone et al. 2015) implies that studies on semiaquatic mammals could carry large bias, particularly if some CTs are closer to water than others. We hypothesized that a ‘worst-case scenario’ would be an otter emerging directly from water into the detection zone, with an anterior or posterior orientation, i.e. travelling towards or away from the CT. An otter after immersion may emit less IR radiation relative to the background (Kuhn and Meyer 2009). Anterior and posterior passes presents a smaller surface area to the PIR sensor and are less likely to create enough movement across the PIR which is required for a trigger (see Rovero and Zimmermann 2016 for further details). One of our CT stations in the otter enclosure faced the pond at a distance of 2.5 m. Thus, we quantified trigger and registration percentages for any anterior passes of otter following immersion, although the sample size (n = 28) was too small for further analyses.
Latency between trigger and registration
Trigger speeds of the CT models were tested by placing a digital clock within the field-of-view of a CT and simultaneously triggering the CT with a moving hand whilst starting the clock; thus, the trigger speed was displayed on the clock in the first frame of the video or still. Across 40 repeats per camera, trigger speeds were: Bushnell video 2.4 s (± 0.1 SD), Bushnell still 0.5 s (± 0.1 SD); Acorn video 2.3 s (± 0.1 SD); and Acorn still 0.7 s (± 0.1 SD).
Results
False-negatives were recorded at each stage of detection we studied (triggering, registering, capture quality), but the extent of false-negatives from each process varied between species, within species (e.g. wet vs dry otters), with CT mode (still vs video) and CT model (Acorn vs Bushnell) (Fig. 4). For all scenarios, at least 20% of passes did not elicit a trigger despite the animal entering the putative detection area (Fig. 4, white bars). For otters, badgers and foxes on videos, a substantial component of false-negatives occurred when the CT triggered but did not register the animal, whilst for stills (otters only) this occurred very infrequently (stippled bars). Based on our specific criteria of recording the animal’s head, substantial data loss occurred due to poor capture quality regardless of whether stills or videos were used, although this varied widely between scenarios (light grey bars). There was substantial variation in the proportion of passes that registered images (combined dark and light grey bars) or images of sufficient quality (dark grey bars).

Success rate of trigger, trigger and registration, and trigger and registration of head as a proportion of the number of passes for a terrestrial mammals on video and dry otter on video and still images b otter passes not from holt c all otter passes (passes from holt and not from holt)
Trigger probability P(trigger|pass)
For the terrestrial mammals and fully dry otters, model comparison results and plots of lowest AIC models are in Fig. 5. DIST and GAIT influenced trigger probability for all species using the Bushnell CTs. DIST has a negative effect in each scenario, with a slower GAIT having greater trigger probability except for the interaction seen in badger where this was only true close to the CT. Trigger rate by the Acorn CT was influenced by AIR and DIST with trigger probability being better at the higher air temperature, but again decreasing with increased DIST.

Model selection tables and plots of the best supported model for Trigger Probability, P(trigger|pass), for a badger with Bushnell camera-trap (CT) on video setting b fox with Bushnell CT on video and c dry otter with Bushnell CT on video and d dry otter with Acorn CT on video. Model variables are defined in Table 1. For brevity, only models with ΔAIC ≤ 2 and the null model are shown in the ranking tables. Full model results are included in online resource 1
Figure 6 shows model comparisons for trigger probability of the best supported models in which fully wet and fully dry otter were considered. With both CT models, DIST had a negative effect, but the negative effect was reduced for dry otter compared to wet.

Model selection tables and plots of the best supported model for Trigger Probability for otter, P(trigger|pass), including the variable WET.DRY, using a Bushnell video and b Acorn video. Model variables are defined in Table 1. For brevity, only models with ΔAIC ≤ 2 and the null model are shown in the ranking tables. Full model results are included in online resource 1
Registration probability P(registration|trigger)
Registration probabilities for the Bushnell still images of otter were almost perfect (i.e. only 2–4% data was lost from cameras triggering but not registering), see Fig. 4, so we did not model these. For videos, registration probability model comparisons are in Fig. 7. Because registration probability is conditional on the camera having triggered, we did not expect the thermal properties of the animal relative to the background to influence it, so we combined wet and dry otter passes for the analysis.

Model selection tables and plots of best models for registration probability P(registration|trigger) for a badger, Bushnell video b fox, Bushnell video c otter (all passes), Bushnell video and d otter (all passes), Acorn video. Only lateral passes were included (see text). Model variables are defined in Table 1. For brevity, only models with ΔAIC ≤ 2 and the null model are shown in the ranking tables. Full model results are included in online resource 1
For video, in each species the model of LOIT+GAIT+DIST had strong support. Notably for registration, the probability increased with distance in most cases, except for Acorn CTs where there was no relationship. In all cases, the registration probability was substantially better when animals were walking and loitering than when they were moving more rapidly.
Capture quality probability
GLMMs were not possible for capture quality probability as loss of data from the trigger and registration stages reduced the number of captured images; furthermore the associated variables (GAIT, LOIT, DIST) were too unevenly distributed. A summary table is provided, see Table 2.
Detection in a ‘worst-case scenario’
For 28 anterior passes of otters emerging from water at the CT station 2.5 m from the pond, the percentage of triggers, registrations and overall capture probabilities are in Table 3.
Discussion
Consideration of the separate component processes of detectability, aligned with their measurable probabilities (Fig. 1) facilitated a clearer understanding of false-negatives when camera-trapping our study species. We demonstrated that substantial data loss through false-negatives can occur at Processes 2–4 (Fig. 4) but that this varies with context (species, camera model, footage type). These false-negatives are driven by different variables as demonstrated by differences between drivers of trigger and registration probabilities. There are some clear methodological considerations that can be drawn from our findings.
PIR sensitivity caused loss of data at close distances
Decreased capture with increased distance is well documented (Rowcliffe 2017; Randler and Kalb 2018), but our data demonstrate this occurs primarily because of reduction in triggering, not a reduction in registering of animals on footage. The PIR sensor receives long-wave infrared (IR) through an 8–14 μm filter. Atmospheric transmission of long-wave IR through air is good (Usamentiaga et al. 2014); therefore, absorption (by atmospheric gases such as CO2 and water vapour) of IR energy between the animal and PIR sensor is not thought to be of consequence (Welbourne et al. 2016). Other mechanisms are therefore needed to explain decreasing trigger probability with increased distance. We suggest that there are two ways that distance can affect the presentation of the animals IR footprint to the PIR sensor. The first relates to the loss of intensity of the animals emitted IR with increasing distance, as the energy per unit area from a point source decreases according to the inverse-square law (Papacosta and Linscheid 2014). The second is that the further away the animal is from the PIR, the more likely there are to be objects or vegetation between the animal and PIR sensor which could block the passage of IR and reduce capture rates (Hofmeester et al. 2017). Whilst distance will always have a predictable negative effect on trigger probability due to the loss of intensity of IR, this will be compounded by objects within the detection zone and lead to variation in the relationship between trigger probability and distance, depending on context, such as local vegetation density.
The negative effect of distance is critical in CT studies that adopt the random encounter model (REM) to estimate population densities when individuals cannot be identified (Rowcliffe and Carbone 2008). This has been an important development in density estimation using camera-traps because capture-recapture methods cannot be applied to species that are not individually identifiable. The REM or similar could be used for all species, therefore removing any potential error from misidentification of individuals. REMs require knowledge of the size of the detection zone of CTs (Rowcliffe et al. 2008). However, because detection probability is variable within the detection zone, distance sampling has been integrated into REMs to estimate effective detection distances for species (i.e. the distance within which the number of animals not captured equals the number captured beyond) (Hofmeester et al. 2017). This relies upon ‘a shoulder of certain detectability up to a certain distance’ from the camera-trap (Rowcliffe et al. 2011), i.e. there is an assumed zone close to the camera with a 100% capture probability for a passing animal. However, we found that at 1 m there was a substantial predicted rate of false-negatives due to trigger failure. At 1 m, trigger probability was already compromised, notably at faster gaits: fox 69%; badger run/trot 58% (walk 88%); and dry otter from holt with Bushnell CTs run/trot 74% (walk 93%). The REM approach is caveated with the assumption that PIR response must be reliable (Rowcliffe et al. 2011). Our trials with two frequently used models of camera-trap demonstrate important limitations in PIR sensitivity. Similar poor capture at close distance (1 m) has also been found in a study of birds (mean of 60% across six size classes of bird and six CT models), where CTs were programmed to capture still images and high sensitivity (Randler and Kalb 2018). We suggest that imperfect triggering at close distances for small to medium homoiotherms may be ubiquitous in CT technology and thus needs to be evaluated prior to distance sampling and other quantitative studies, with a CCTV control being a useful method.
Speed is important in registration probability
Gait was an important variable affecting trigger probability for badger and dry otter, but less so for fox with a slower gait increasing trigger probability. We used gait to represent the relative speed of passes within each species, but in some species, there is also a difference in the vertical movement (i.e. bounce) as well as horizontal movement with different gaits. The bouncing gait of a trotting badger will interact with a larger proportion of its background, possibly creating a better signal to the PIR. This may lessen the effect of distance on trigger probability, as seen in the interaction of GAIT and DIST in Fig. 5. There was a more consistent effect of gait on registration probability, in all cases slower passes are more likely to register in an image/video; see Fig. 7. Observations of running animals were rare in our study, and this has been noted in other mammal groups such as the Felidae (Anile and Devillard 2016), so speed may cause greater bias in multispecies surveys where species move at different speeds affecting both trigger and registration probability (Hofmeester et al. 2019).
Distance drives trigger and registration probability in opposite directions
In contrast to the strong negative effect of distance on trigger probability, there was a positive, though less marked, relationship between distance and registration probabilities when using Bushnell CTs on video setting. This is likely a function of the time interval between the PIR detecting the animal and the camera switching on, i.e. the trigger speed. Registration probability for CTs recording video was consistently affected by gait, loitering and distance across species and CT models, contrasting with the minimal data loss due to high registration probability on ‘still’ image setting. The longer trigger speed of videos (just over 2 s) required slower passes and/or loitering (e.g. to scent mark or sniff) to achieve better registration probability. Also, the further the subject is from the CT, the greater the width of field-of-view of the camera, and therefore it takes longer to pass through the field-of-view and is more likely to be within it when the camera starts recording.
A hypothetical scenario illustrating a mechanism by which registration probability for a lateral pass is likely to increase with distance, and how this is likely to interact with animal speed, is shown in Fig. 8. This interpretation presents a hypothesis that could be tested in future experiments.

Hypothesized mechanism showing how distance to camera-trap (CT) can interact with animal speed to influence registration probability. Registration probability is positively affected by distance due to the larger area within the field-of-view at greater distances. Conversely, faster moving animals can completely pass through the small width of the field-of-view close to the CT before the camera takes an image
Given this reasoning, a stronger positive effect of distance on registration probability would have been expected with the Acorn CTs due to their wider field-of-view, but this was not observed. The Acorn’s wide field-of-view led to difficulties identifying otter at greater distances as the otter had a smaller apparent size, thus reducing registration probability.
The choice between still image and video capture
The fast trigger speed for Bushnell still images resulted in high registration probability: 96–98% of passes that triggered resulted in the otter being registered. This contrasts with the registration probability for Bushnell videos, where a lower 65–79% of passes that triggered resulted in registered otter. Survey design therefore needs to consider potential false-negatives due to longer trigger speeds of the video setting, which should influence the choice of CT make/model. Video capture, however, can facilitate behavioural observations which may be essential but are not possible with still capture. For example, animal vocalizations can be recorded on video mode with CT models that have microphones.
Still capture is indicated for capture-recapture density studies where a key consideration is high-quality images to distinguish pelage details (Trolliet et al. 2014); still capture also enables the use of xenon white flash. It is also more efficient for faunal inventories and occupancy studies where data generated by videos is not usually required. Density studies using REM can use video or a burst of still images to the estimate average speed of an animal (Rowcliffe et al. 2016). Whilst there will be lost data from both settings due to imperfect trigger probability, the video setting is also likely to have reduced registration probability, unless the trigger speeds are comparable. Where data from video is required, for instance, in behavioural studies, CTs should be aimed at areas with field signs indicating activity that delays the passage of a passing animal, such as at dens, bait stations or scent marking sites.
Although trigger speeds for video recording are generally slower than for still images, models are now available with an advertised trigger speed of less than 1 s (e.g. Bushnell Core DS), and these could be chosen if video is the preferred mode of study to increase registration probability. An additional constraint for video recording is that video data requires more storage capacity, and viewing video footage takes longer than still images. Whilst software to enable automated species identification is being developed and may be used in the future, this is directed at still images (Yu et al. 2013; Tabak et al. 2019).
Effects of immersion of otter on detection are short-lived
The trigger probability of dry otter passes on Bushnell videos broadly reflected those of the two terrestrial species, with distance and/or gait being important in all the best fitting models although the best supported model for the Acorn video CT included air temperature and distance. Our results corroborate observations that wet otters are poor in eliciting a PIR trigger (Lerone et al. 2015). However, time from exiting water was not an important variable in trigger success, indicating that other variables may impact on the rate of change in IR emitted after an otter has left water. Otter thermoregulation in cold water can result in reduced emission of IR from an otter’s body and tail; however, the intensity and duration of swimming prior to exiting water can affect thermoregulation and hence the amount of IR emitted (Kuhn and Meyer 2009). These variables, and others, may confound any effect of time from exiting water on trigger success. When we set a CT facing water at 2.5 m to record otter emerging from water, the trigger probabilities for Bushnell (video and still) and Acorn CTs were very poor (36–43%). The slower trigger speed for video led to poor registration probability of 40–63% (Table 2); the resulting capture of all passes on video setting (e.g. 14% for Acorn) is unlikely to be fit for any purpose. Within the limits of our study conditions and limited sample size, thermal imaging readings indicated that when an otter emerges from water, its surface temperature nearly matches water temperature (see Fig. 3). It only takes a short period of time from immersion (≤ 1 min) for an otter to develop a thermal footprint with a 3 °C difference from the background, 3 °C being an approximate difference that would trigger a camera-trap PIR (Meek et al. 2012). Although this is likely to be affected by background temperatures, and the otter’s prior activity, it indicates such effects are potentially short-lived.
Understanding the stages of detectability will improve study design
CTs can be used for a range of study types; hence, study design needs to consider CT model specifications, placement and settings (Rovero and Zimmermann 2016). Recognition of detection as a sequence of processes (Fig. 1) enables each process to be considered independently when planning CT studies, as the mechanisms for success in each process are different. Understanding how the animal, environment and equipment interact is important for all CT studies and can help in considering potential bias, for example, from detection heterogeneity between sites or species in a study. We demonstrate the high level of data loss (on both video and still setting) on medium-sized animals due to poor triggering, even at close distances. This would need to be accounted for within population density analyses such as the REM when distance sampling is used to estimate effective detection distances. Using CCTV as a control, the influences of different seasons, temperatures, humidity and vegetation structure could also be quantified.
We found that trigger probability for otter was compromised after recent emergence from water, and it is anticipated that this would apply for other semiaquatic species. In a pilot study, we also found very low trigger probabilities for European beaver Castor fiber in an enclosure where they spent a significant time in water (unpubl. data). Careful CT placement is therefore critical when studying semiaquatic mammals and CTs set on in-stream features such as stones or on entry/exit points from water are likely to have poor trigger probability, as previously demonstrated (Lerone et al. 2015). Trigger probability would improve if CTs were set to anticipate semiaquatic mammal passes where the animal has been out of water long enough to develop a better thermal footprint.
We would recommend that the trigger speed of the chosen CT model and mode of recording is established, either from the manufacturer’s specification or via testing. Video trigger speeds are rarely specified by manufacturers, perhaps because they are usually significantly slower than those for still images.
Conclusions
Our approach has demonstrated where false-negatives potentially occur during the process of detection using camera-traps and what factors drive variation in trigger and registration probabilities, and this can help optimize camera-trap deployments to try to reduce false-negatives given the study species, environmental context and study aims. Our findings could generalize to other species of medium-sized terrestrial and semi-aquatic mammals. Similarly, this approach, using CCTV as a control to separate component processes of detection (trigger, registration and capture quality), could be carried out as a precursor to CT studies in different contexts, such as with small or large mammals, or in different seasons and environmental conditions. Results could be used to inform modelling of detection functions for REM with distance sampling and would help to improve study design more widely.
References
Allaire JJ (2014) Manipulate: interactive plots for RStudio. R package version 1.0.1. https://CRAN.R-project.org/package=manipulate
Anile S, Devillard S (2016) Study design and body mass influence RAIs from camera trap studies: evidence from the Felidae. Anim Conserv 19:35–45. https://doi.org/10.1111/acv.12214
Barton K (2016) MuMIn: Multi-Modal Inference. R Package Version 1.42.1. https://CRAN.R-project.org/package=MuMIn
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Burnham KP, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res 33:261–304. https://doi.org/10.1177/0049124104268644
Findlay MA, Briers RA, Diamond N, White PJC (2017) Developing an empirical approach to optimal camera-trap deployment at mammal resting sites: evidence from a longitudinal study of an otter Lutra lutra holt. Eur J Wildl Res 63:96–13. https://doi.org/10.1007/s10344-017-1143-0
Foster RJ, Harmsen BJ (2012) A critique of density estimation from camera-trap data. J Wildl Manag 76:224–236. https://doi.org/10.1002/jwmg.275
Gužvica G, Bošnjafile I, Bielen A et al (2014) Comparative analysis of three different methods for monitoring the use of green bridges by wildlife. PLoS One 9:1–12. https://doi.org/10.1371/journal.pone.0106194
Hamel S, Killengreen ST, Henden JA et al (2013) Towards good practice guidance in using camera-traps in ecology: influence of sampling design on validity of ecological inferences. Methods Ecol Evol 4:105–113. https://doi.org/10.1111/j.2041-210x.2012.00262.x
Hofmeester TR, Rowcliffe JM, Jansen PA (2017) A simple method for estimating the effective detection distance of camera traps. Remote Sens Ecol Conserv 3:81–89. https://doi.org/10.1002/rse2.25
Hofmeester TR, Cromsigt JPGM, Odden J, Andrén H, Kindberg J, Linnell JDC (2019) Framing pictures: a conceptual framework to identify and correct for biases in detection probability of camera traps enabling multi-species comparison. Ecol Evol 9:2320–2336. https://doi.org/10.1002/ece3.4878
Karanth KU (1995) Estimating tiger Panthera tigris populations from camera-trap data using capture-recapture models. Biol Conserv 71:333–338. https://doi.org/10.1016/0006-3207(94)00057-W
Kuhn R, Meyer W (2009) Infrared thermography of the body surface in the Eurasian otter Lutra lutra and the giant otter Pteronura brasiliensis. Aquat Biol 6:143–152. https://doi.org/10.3354/ab00176
Larrucea ES, Brussard PF, Jaegar MM, Barrett RH (2007) Cameras, coyotes, and the assumption of equal detectability. J Wildl Manag 71:1682–1689. https://doi.org/10.2193/2006-407
Lerone L, Carpaneto GM, Loy A (2015) Why camera traps fail to detect a semi-aquatic mammal: activation devices as possible cause. Wildl Soc Bull 39:193–196. https://doi.org/10.1002/wsb.508
Lim NTL, Ng PKL (2008) Home range, activity cycle and natal den usage of a female Sunda pangolin Manis javanica (Mammalia: Pholidota) in Singapore. Endanger Species Res 4:233–240. https://doi.org/10.3354/esr00032
Meek P, Ballard G, Fleming PJS (2012) An introduction to camera trapping for wildlife surveys in Australia. Animals Cooperative Research Centre, Canberra
Mugerwa B, Sheil D, Ssekiranda P et al (2013) A camera trap assessment of terrestrial vertebrates in Bwindi impenetrable National Park, Uganda. Afr J Ecol 51:21–31. https://doi.org/10.1111/aje.12004
O’Connor KM, Nathan LR, Liberati MR et al (2017) Camera trap arrays improve detection probability of wildlife: investigating study design considerations using an empirical dataset. PLoS One 12:1–12. https://doi.org/10.1371/journal.pone.0175684
Papacosta P, Linscheid N (2014) The confirmation of the inverse square law using diffraction gratings. Phys Teach 52:243–245. https://doi.org/10.1119/1.4868944
Randler C, Kalb N (2018) Distance and size matters: a comparison of six wildlife camera traps and their usefulness for wild birds. Ecol Evol 8:7151–7163. https://doi.org/10.1002/ece3.4240
RCore Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rovero F, Zimmermann F (eds) (2016) Camera trapping for wildlife research, 1st edn. Pelagic Publishing,UK, Exeter
Rovero F, Zimmermann F, Berzi D, Meek P (2013) “Which camera trap type and how many do I need?” a review of camera features and study designs for a range of wildlife research applications. Hystrix 24:148–156. https://doi.org/10.4404/hystrix-24.2-6316
Rowcliffe M (2017) Key frontiers in camera trapping research. Remote Sens Ecol Conserv. https://doi.org/10.1002/rse2.65
Rowcliffe JM, Carbone C (2008) Surveys using camera traps: are we looking to a brighter future? Anim Conserv 11:185–186. https://doi.org/10.1111/j.1469-1795.2008.00180.x
Rowcliffe JM, Field J, Turvey ST, Carbone C (2008) Estimating animal density using camera traps without the need for individual recognition. J Appl Ecol 45:1228–1236. https://doi.org/10.1111/j.1365-2664.2008.01473.x
Rowcliffe MJ, Carbone C, Jansen PA et al (2011) Quantifying the sensitivity of camera traps: an adapted distance sampling approach. Methods Ecol Evol 2:464–476. https://doi.org/10.1111/j.2041-210X.2011.00094.x
Rowcliffe JM, Jansen PA, Kays R et al (2016) Wildlife speed cameras: measuring animal travel speed and day range using camera traps. Remote Sens Ecol Conserv 2:84–94. https://doi.org/10.1002/rse2.17
RStudioTeam (2015) RStudio: Integrated Development for R
Silver SC, Ostro LET, Marsh LK et al (2004) The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx 38:148–154. https://doi.org/10.1017/S0030605304000286
Sollmann R, Kelly MJ (2013) Camera trapping for the study and conservation of tropical carnivores. Raffles Bull Zool 28:21–42
Swinnen KRR, Reijniers J, Breno M, Leirs H (2014) A novel method to reduce time investment when processing videos from camera trap studies. PLoS One 9:e98881. https://doi.org/10.1371/journal.pone.0098881
Tabak MA, Norouzzadeh MS, Wolfson DW et al (2019) Machine learning to classify animal species in camera trap images: applications in ecology. Methods Ecol Evol 10:585–590. https://doi.org/10.1111/2041-210X.13120
Tobler MW, Carrillo-Percastegui SE, Leite Pitman R et al (2008) An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Anim Conserv 11:169–178. https://doi.org/10.1111/j.1469-1795.2008.00169.x
Tobler MW, Zúñiga Hartley A, Carrillo-Percastegui SE, Powell GVN (2015) Spatiotemporal hierarchical modelling of species richness and occupancy using camera trap data. J Appl Ecol 52:413–421. https://doi.org/10.1111/1365-2664.12399
Trolliet F, Huynen M-C, Vermeulen C, Hambuckers A (2014) Use of camera traps for wildlife studies. A review. Biotechnol Agron Soc Environ 18:466–454
Usamentiaga R, Venegas P, Guerediaga J, Vega L, Molleda J, Bulnes FG (2014) Infrared thermography for temperature measurement and non-destructive testing. Sensors 14:12305–12348. https://doi.org/10.3390/s140712305
Welbourne DJ, Claridge AW, Paull DJ, Lambert A (2016) How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions. Remote Sens Ecol Conserv 2:77–83. https://doi.org/10.1002/rse2.20
Yu X, Wang J, Kays R et al (2013) Automated identification of animal species in camera trap images. Eurasip J Image Video Process:2013. https://doi.org/10.1186/1687-5281-2013-52
Acknowledgments
Joanna Coxon, Prof. John Currie, Laura Duggan, Marianne Freeman, Royal Zoological Society of Scotland staff, Ben Harrower, Simon Girling, Derek Gow and staff, Roger Ingledew, Douglas Richardson, Susan Maclean, Alice Martin-Walker, Estelle Roche, Alarms for Farms (for CCTV equipment), Pakatak (for donating Acorn camera traps). Two anonymous reviewers commented on earlier drafts.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Dries Kuijper
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Online resources: The R file and datasets are available at: https://github.com/melaniefindlay/CT-Detection
ESM 1
(DOCX 546 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Findlay, M.A., Briers, R.A. & White, P.J.C. Component processes of detection probability in camera-trap studies: understanding the occurrence of false-negatives. Mamm Res 65, 167–180 (2020). https://doi.org/10.1007/s13364-020-00478-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-020-00478-y