Fungal Diversity in the Phyllosphere of Pinus heldreichii H. Christ—An Endemic and High-Altitude Pine of the Mediterranean Region


1
Biotechnical faculty, University of Montenegro, Mihaila Lalića 15, 81 000 Podgorica, Montenegro
2
Department of Forest Mycology and Plant Pathology, Uppsala BioCenter, Swedish University of Agricultural Sciences, P.O. Box 7026, SE-75007 Uppsala, Sweden
*
Author to whom correspondence should be addressed.
Diversity 2020, 12(5), 172; https://doi.org/10.3390/d12050172
Submission received: 30 March 2020
/
Revised: 16 April 2020
/
Accepted: 21 April 2020
/
Published: 28 April 2020
(This article belongs to the Special Issue Fungal Diversity in the Mediterranean Area)
Abstract
:Pinus heldreichii is a high-altitude coniferous tree species naturaly occurring in small and disjuncted populations in the Balkans and southern Italy. The aim of this study was to assess diversity and composition of fungal communities in living needles of P. heldreichii specifically focusing on fungal pathogens. Sampling was carried out at six different sites in Montenegro, where 2-4 year-old living needles of P. heldreichii were collected. Following DNA isolation, it was amplified using ITS2 rDNA as a marker and subjected to high-throughput sequencing. Sequencing resulted in 31,831 high quality reads, which after assembly were found to represent 375 fungal taxa. The detected fungi were 295 (78.7%) Ascomycota, 79 (21.0%) Basidiomycota and 1 (0.2%) Mortierellomycotina. The most common fungi were Lophodermium pinastri (12.5% of all high-quality sequences), L. conigenum (10.9%), Sydowia polyspora (8.8%), Cyclaneusma niveum (5.5%), Unidentified sp. 2814_1 (5.4%) and Phaeosphaeria punctiformis (4.4%). The community composition varied among different sites, but in this respect two sites at higher altitudes (harsh growing conditions) were separated from three sites at lower altitudes (milder growing conditions), suggesting that environmental conditions were among major determinants of fungal communities associated with needles of P. heldreichii. Trees on one study site were attacked by bark beetles, leading to discolouration and frequent dieback of needles, thereby strongly affecting the fungal community structure. Among all functional groups of fungi, pathogens appeared to be an important component of fungal communities in the phyllosphere of P. heldreichii, especially in those trees under strong abiotic and biotic stress.
1. Introduction
Pinus heldreichii is a high-altitude (grows at ca. 1200–2000 m) conifer tree species with a discontinuous and restricted distribution in the Mediterranean region. Its forests are naturaly regenerated and consists of small and disjuncted populations located in high mountain areas influenced by the Mediterranean climate in the Balkans and southern Italy. Although in the past P. heldreichii formed a continuous forest belt in the Balkans, currently its forests are scattered and largely isolated. Growing primary on shallow calcareous soils, it inhabits typical tree line habitats, often on steep ridges, mountain sides and screens. Such habitats are nutrient poor, exposed, dry and cold during the winter [1,2]. Larger P. heldreichii forests can still be found on mountain plateaus or in valleys situated at altitudes of ca. 1200–1300 m that are characterised by more developed soils (cambisols). Those forests are considered to be climazonal i.e., characterised by permanent and stable vegetation forests of P. heldreichii, that are growing at ecological optimum for the species [3]. Being a tertiary relic, P. heldreichii evolved to survive severe winter frosts, and short, dry and warm summers with intensive sun radiation. Due to the short growing season, P. heldreichii grows very slowly, reaching ca. 15 m height after about 150 years [1]. It develops thick and branched roots, which penetrate deep in cracks of calcareous stones [2]. In the last few centuries, P. heldreichii forests were affected by intensive livestock grazing, exploitation and forest fires [4]. Intensive grazing, as well as the short and dry growing season have also resulted in very limited natural regeneration [2,3,4], though recent observations show some regeneration in the abandoned mountain areas [5].
Pinus heldreichii is a protected species both in Balkan countries and in Italy due to the key importance of nature conservation, protection against gravitational natural hazards, landscape conservation and recreation. Hence, P. heldreichii requires special attention, i.e., the development and application of conservation measures [1,2,3,4,6].
Fungi represent the largest microbial component associated with forest trees. They play key roles in forest ecosystems, especially in pines forest that are obligatory mutualistic, and are important contributors to the primary production and carbon, nutrient and water cycling [7,8,9]. Pathogenic fungi may negatively affect health and growth of forest trees [10,11], while fungal endophytes and epiphytes support ecological adaptations of host plants and constitute an important component of microbial biodiversity [12,13]. However, information about fungal communities associated with P. heldreichii is limited. Previous studies on needle pathogens of P. heldreichii have focused on either those that were affecting natural regeneration [14,15,16], or potentially invasive ones such as Dothistroma septosporum [17]. Information about the occurrence of D. septosporum in stands of P. heldreichii was not known until recently [17], what indicated the need for a wider assessment of fungal diversity, including fungal pathogens associated with the phyllosphere of this tree species.
The aim of this study was to assess the diversity and composition of fungal communities in living needles of P. heldreichii specifically focusing on fungal pathogens. Needles were sampled across the natural distribution range of P. heldreichii in Montenegro, including sites under different environmental conditions. This was expected to demonstrate potential site-specific effects of environmental conditions on fungal diversity and community composition. By using high-throughput sequencing of fungal ITS2 rDNA, we examined fungal communities in 2-4 year-old living needles of P. heldreichii from six sites situated in four mountain areas in Montenegro.
2. Materials and Methods
2.1. Study Sites
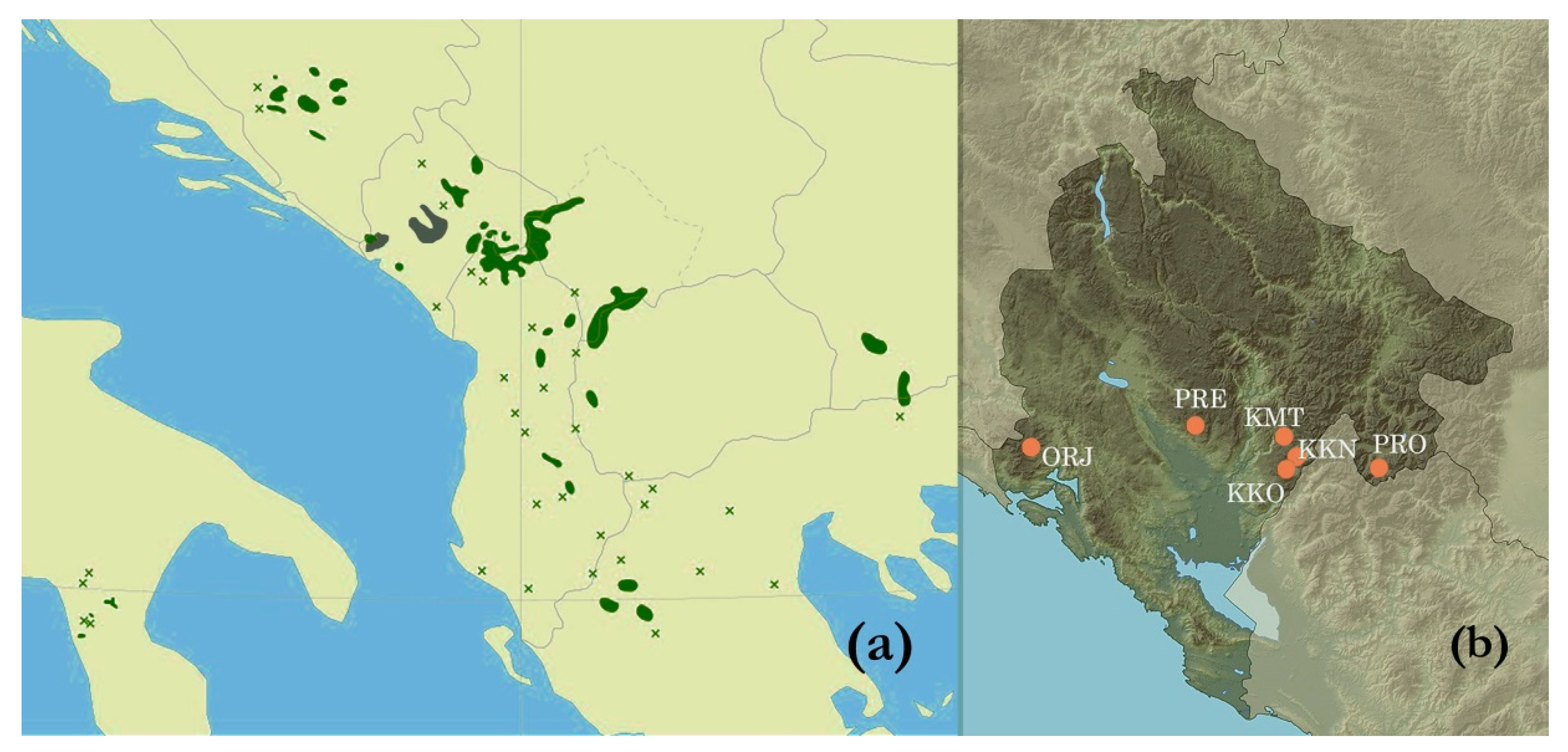
Samples were taken at six sites that were situated in four mountain areas, representing the P. heldreichii distribution range in Montenegro (Figure 1). The sampling sites were Orjen (ORJ), Prekornica (PRE), Kučka Korita (KKO), Kučka korita North (KKN), Kuči MT (KMT) and Prokletije (PRO) (Figure 1, Table 1).
The sampling sites differed in altitude, morphology of terrain and soil properties (Table 1, Figure 2). Sites at ORJ and KMT were on altitudes of ca. 1800 m, with trees growing on stone ridges, high inclinations, and lithosols. Sites at PRE, PRO and KKO were on an altitude of ca. 1250 m, on flat terrain and more developed soil that was leptic cambisol. A site at KKN was in close proximity to the KKO, but it was on a slope with a thinner soil layer (molic leptosol). Trees at the KKN were in groups heavily infested by Tomicus sp. bark beetles (Scolitinae). Trees at ORJ and KMT were growing at the upper tree line, and thus, these sites were considered as sites with harsh growing conditions, while other sites had moderate growing conditions.
Climate at the study sites is characterised as a humid warm temperate climate type (Cf), which is represented by subtypes Cfsb, Cfs’’b and Cfb [22,23]. Summers are short, dry and chilly, and winters are cold and windy. At different study sites, the mean annual air temperature is between 1 °C (at PRO) and 6 °C (at ORJ). The mean daily summer maximum is between 7 and 11 °C, and the mean daily winter minimum is between −5 and 0 °C. The absolute minimum is in winter at ca. −30 °C and the maximum is in summer, at ca. 30 °C. The mean annual precipitation is highest at ORJ with ca. 3800 mm, and lowest at PRO with ca. 2000 mm. The rainfall reaches maximum in late autumn and early winter, while the minimum is during the summer months that is often followed by 40–70 day-long periods of droughts. The mean summer precipitation is 300 mm at ORJ and ca. 220 mm on other sites, making ca. 10% of the total annual precipitation [22].
The bedrock at the study sites is solid chalk limestone, which contains a small proportion of insoluble residues. In general, soils are poorly developed, very water porous, skeletal leptosols (rendzina), having only A horizon, characterized by accumulation of humus (A-R profile). According to the classification of texture, the soil is a sandy loam and the structure is powdery. Soil is faintly differentiated down in the soil profile. Soil is rich in humus content (10–25%), poor in calcium-carbonate and has a weak acid reaction (pH ca. 6). On more flat terrains, leptosol develops into the leptic cambisol, which contains initial B horizon. Brown B horizon improves the water retention capacity of soil [20,24].
2.2. Experimental Design and Sampling
At the sampling sites, stands of P. heldreichii were healthy-looking (Figure 2a). An exception was the KKN site, where trees were damaged by Tomicus sp. bark beetles, resulting in discolouration and fungal infection of needles. At all sites, the shoot dieback was occasional, but with higher rates at the KKN site.
Sampling was carried out in May 2015, i.e., before the beginning of the growing season. At each site, experimental design included sampling of five twigs with needles from five different trees. Hence, at each site, five mature P. heldreichii trees situated at a distance of ca. 50 m from each other were selected and five twigs of up to 15 cm long and up to 2 m from the ground were sampled from different parts of the crown using secateurs. This sampling approach was used in order (i) to get a joint representative sample per each site (all samples per site were amplified with the same barcode, see below); (ii) to compare fungal communities among different sites, but not within the same site. Collected twigs were placed in plastic bags and transported to the laboratory.
In the laboratory, twigs and needles were assessed for the presence of disease symptoms. The current-year needles were typically green and without disease symptoms. Symptomatic needles were generally two years old or older and symptoms included changed colour that was yellow, orange, pale green or yellowish with red bands. Other symptoms were dying needle tips and a necrotic base of needles. In the case of insect attack (KKN site), defoliation and necrotic needles were common. In order to sample the potentially entire fungal community associated with needles of P. heldreichii, both healthy looking needles and needles with disease symptoms were selected. No surface sterilization was carried out. Following morphological examination, up to 5 representative needles that were 2–4 years old, were selected per twig. Selected needles were cut into 0.5–1 cm long segments, containing a random mixture of both asymptomatic and symptomatic parts of needles, placed in 2 mL screw cap tubes, and stored frozen at −20 °C before DNA extraction.
2.3. DNA Isolation, Amplificationand Sequencing
DNA extractions were done from 150 samples (6 sites × 5 trees × 5 needle samples), which previously were freeze-dried for 48 h. For isolation of total DNA, needles were homogenised in a Fastprep machine (Precellys, Montigny-le-Bretonneux, France). The extraction was completed using CTAB protocol [25]. After extraction, DNA samples were purified using a JetQuick DNA purification kit (Genomed GmbH, Leinfelden, Germany). The DNA concentration of each sample was determined using a NanoDrop™ One spectrophotometer (Thermo Scientific, Rodchester, NY, USA) and adjusted to 1–10 ng/µL. Amplification by PCR of the ITS2 rDNA region was done using barcoded fungal-specific primer gITS7 [26] and barcoded universal primer ITS4 [27]. All 25 samples from the same site were amplified using primers with the same barcode, resulting in six different barcodes representing each site. Amplification of several samples with the same barcode was done to get a broader representativeness of fungal communities per site. Amplifications were performed using the Applied Biosystems 2720 thermal cycler (Foster City, CA, USA). An initial denaturation step started at 95 °C for 2 min, followed by 27 amplification cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 60 s. The thermal cycling was ended by a final extension step at 72 °C for 7 min. The PCR products were analyzed using gel electrophoresis on 1% agarose gels stained with Nancy-520 (Sigma-Aldrich, Sweden). PCR products were purified using a sodium acetate protocol [28]. Purified PCR products were quantified using a Qubit fluorometer 4.0 (Thermo Fisher Scientific, Waltham, MA, USA), and an equimolar mix of all PCR products with six barcodes (one per each site) was used for high-throughput sequencing using a Pacific Biosciences platform (Menlo Park, CA, USA) at the SciLifeLab (Uppsala, Sweden).
2.4. Bioinformatics
Principles of bioinformatics followed that specified in [29]. The sequences obtained from the six samples that represented six sampling sites (Table 2), were subjected to quality control and clustering in the SCATA NGS sequencing pipeline [30]. The initial procedure started with quality filtering of the sequences that included the removal of sequences shorter than 200 bps, sequences with low read quality, primer dimers and homopolymers, which were collapsed to 3 bps before clustering. Only sequences containing a barcode and primer were retained. Then, the primer and sample barcodes were removed from the sequence, but information on the sample and sequence association was stored as meta-data. A single-linkage clustering based on 98% similarity was used to cluster sequences into different taxa. For each cluster, the sequence of the most common genotype was used for taxonomic identification. For clusters containing only two sequences, a consensus sequence was produced. The taxa were taxonomically identified using the GenBank database and the Blastn algorithm [31]. The following criteria were used for identification: sequence coverage >80%; 94–97% similarity to genus level and >98% similarity to species level. Sequences deviating from these criteria were identified only to a high taxonomic rank and were given unique names as shown in Table 3 and Table S1. Representative sequences of fungal nonsingletons are available from GenBank under accession numbers MT241905–MT242268.
2.5. Statistical Analyses
Rarefaction analysis was carried out using Analytical Rarefaction v.1.3 [32]. Differences in richness of fungal taxa in different study sites of P. heldreichii were compared by nonparametric chi-square testing [33]. As each of the datasets was subjected to multiple comparisons, confidence limits for p-values of chi-square tests were reduced the corresponding number of times as required by the Bonferroni correction [34]. The Shannon diversity index, qualitative Sorensen similarity index and nonmetric multidimensional scaling (NMDS) in Canoco 5 [33,35,36] were used to characterize the diversity and composition of fungal communities.
3. Results
The phyllosphere fungi of P. heldreichii were examined from four different mountain regions, represented by six sampling sites across the distribution range of P. heldreichii in Montenegro (Figure 1). Amplification of fungal ITS2 rDNA from 150 needle samples, PacBio sequencing and quality filtering resulted in 31,829 high quality reads. Sequence assembly and BLASTn analyses showed that the fungal community in the phyllosphere of P. heldreichii was composed of 375 fungal taxa (Table 2, Table S1). Nonfungal taxa and singletons were excluded. The detected fungi were 295 (78.7%) Ascomycota, 79 (21.0%) Basidiomycota and 1 (0.2%) Mortierellomycotina. Identification at least to genus level was successful for 254 (67.7%) out of 375 fungal taxa (Table S1), and those represented 78.5% of all fungal sequences.
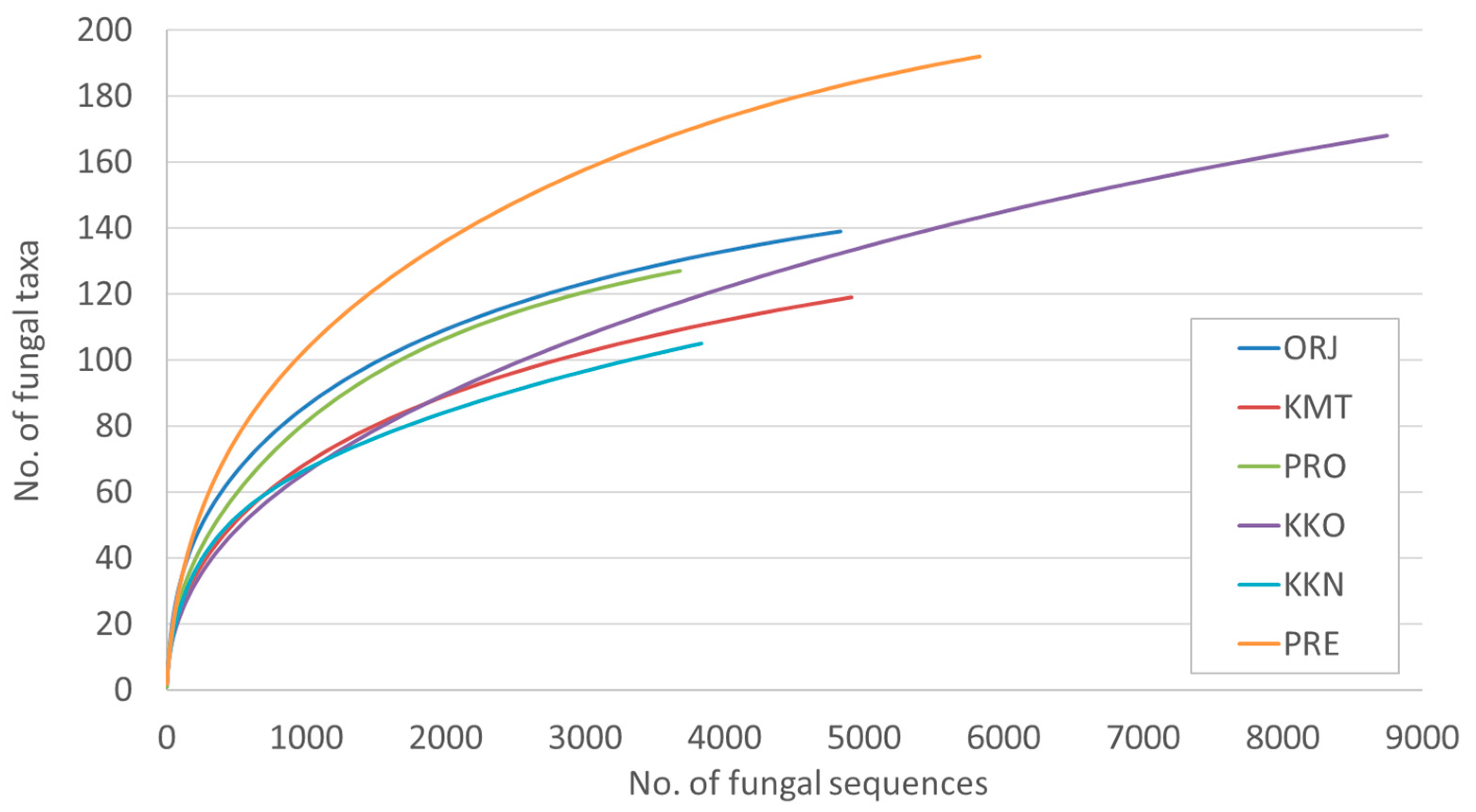
In different study sites, the number of fungal taxa varied between 104 and 192 (Table 2). The chi-square test showed that the largest difference in richness of fungal taxa was between KKO and the remaining sites (Table 2). Rarefaction showed that fungal taxa detected in all sites did not reach the species saturation (Figure 3).
Information on the 25 most common fungal taxa representing 79.0% of all fungal sequences is in Table 3. The most common fungi in the phyllosphere of P. heldreichii were Lophodermium pinastri (12.5% of all fungal sequences), Lophodermium conigenum (10.9%), Sydowia polyspora (8.8%), Cyclaneusma niveum (5.5%) and Unidentified sp. 2814_1 (5.4%) (Table 3). The most common fungal pathogens of pine needles were S. polyspora (8.9%), C. niveum (5.5%), Neocatenulostroma germanicum (2.3%), Allantophomopsiella pseudotsugae (2.1%) and Cenangium acuum (0.7%) (Table 3). A pine needle pathogen, Dothistroma septosporum (0.25%) was also detected, but at lower relative abundance (Table S1). The other detected pathogenic species that are mainly known as wound pathogens of deciduous trees and/or agricultural crops, were Phaeomoniella 2814_15 (3.0%), Ramoconidiophora euphorbiae (1.5%), Collophorina sp. (0.9%), Geastrumia sp. (1.4%) and Athelia acrospora (0.7%). The detected fungal endophytes that are known to produce antimicrobial metabolites were Phaeosphaeria pontiformis (4.4%), Microsphaeropsis olivacea (2.2%), Lachnellula calyciformis (2.0%) and Mollisia ligni (0.7%). Ubiquitous saprotrophs were rare (Table S1).
The community composition of the phyllosphere fungi varied among different sites (Figure 4). Dothideomycetes dominated fungal communities at PRO and KKN sites, while Leotiomycetes dominated at KMT and PRE sites (Figure 4). At ORJ and KKO sites, the relative abundance of these two classes was similar (Figure 4). Sordariomycetes showed higher relative abundance at ORJ (22.6%), while all other fungal classes in different sampling sites were less abundant (Figure 4). Agaricomycetes (Basidiomycotina) were rare and their relative abundance at different sites varied between 0.03% and 6.3% (Figure 4).
The NMDS of fungal communities associated with needles of P. heldreichii (Figure 5) showed that KKO, PRO and PRE sites clustered together and were separated from the remaining sites along axis 1. Fungal communities at the ORJ and KMT showed a closer proximity (Figure 5), while fungal communities at the KKN site differed from all remaining sites and were separated along both axis 1 and 2.
At ORJ and KMT sites that are characterized by harsh growing conditions, the fungal community was dominated by Lophodermium pinastri, Sydowia polyspora, Phaeomoniella sp., Allanthopomopsiella pseudotsugae, Lachnellula calyciformis, Ramoconidiophora euphorbiae, Collophorina sp., Geastrumia sp. and Mollisia ligni (Table 3). At PRE, PRO and KKO sites that are characterized by moderate growing conditions, fungal community was dominated by Lophodermium conigenum, L. pinastri, Cyclaneusma niveum, Phaeosphaeria punctiformis, Unidentified sp. 2814_11, Cenangium acuum and Dothistroma septosporum. At the KKN, with insect attacks and partly necrotic needles, fungal community was dominated by S. polyspora, Microsphaeropsis olivacea, Neocatelunostroma germanicum, Uncultured sp. 2814_10, Chaetothyriales 2814_18 and Geastrumia sp. (Table S1).
4. Discussion
A recent development of high-throughput sequencing methods provides powerful tools to explore fungal diversity. Such tools enable identification of complex fungal communities and individual community components directly from environmental samples. Besides, while providing detailed and semi-quantitative information, these tools enable studying the effects of different factors on fungal diversity and community composition [29,37,38]. By using PacBio sequencing, we detected nearly 400 fungal taxa, which were associated with needles of P. heldreichii including fungi present in very small abundances (Table S1). Many fungal taxa remained unidentified, which remains a major challenge in fungal taxonomy [39]. However, while using high-throughput sequencing, we need be aware of potential limitations that can include methodological biases, limitations of markers and bioinformatics challenges [40,41].
The observed diversity of the phyllosphere fungi can be considered being high and comparable with similar studies on different Pinus species. For example, there were 446 and 260 fungal taxa recorded during studies of needle-associated fungi of Pinus sylvestris in Sweden and Poland, respectively [39,42]. Furthermore, by using fungal culturing, there were 118 fungal taxa associated with needles of Pinus taeda in USA [43], 35 taxa associated with needles of Pinus halepensis [44] and P. sylvestris in Spain [45]. According to [45], the species diversity of fungal endophytes varied from 21 in Pinus monticola to 49 in Pinus nigra. At different study sites, the growing conditions of P. heldreichii had in general a minor effect of the absolute richness of fungal taxa, which was similar among different sites (Table 2). However, with respect to the number of sequences obtained from samples of each site, lower richness of fungal taxa was at the KKO (Table 2), what was likely due to the dominance of several fungal taxa such as, e.g., Lophodermium species, Cylaneusma niveum, Unidentified sp. 2814_11, Phaeosphaeria pontiformis and Unidentified sp. 2814_1. Several fungal taxa of the present study were recorded for the first time in the Balkan region such as, e.g., pathogens Neocatelunostroma germanicum and Allantophomopsiella pseudotsugae. The study also revealed a number of fungal pathogens that were previously known from agricultural crops as, e.g., Phaeomoniella, Ramoconidiophora, Geastrumia and Athelia. These could have been overlooked in other studies due to latent occurrence [46]. As P. heldreichii belongs to a group of forest trees that grows under harsh enviromental conditions, the possibility should not be excluded that both the host and the specific environment influence the composition of associated fungal communities. Studies on plant–endophyte associations in high stress habitats have revealed that at least some fungal endophytes can contribute to stress tolerance of host plants [12]. Indeed, the results of this study revealed site-specific differences in fungal communities associated with needles of P. heldreichii. For example, fungal communities were more similar among sites situated at lower altitudes, i.e., with moderate growing conditions (PRE, PRO, KKO) as compared to those sites at higher altitudes, i.e., with harsh growing conditions (ORJ, KMT) (Table 1, Figure 5). Further, the fungal community at the KKN site was largely different from the remaining sites, what was likely due to insect damage to trees, resulting in mainly symptomatic and partly necrotic needles. These observations demonstrate that the composition of fungal communities and their succession in needles of P. heldreichii can be determined by different abiotic and biotic factors. Different environmental factors have earlier been shown to play an important role in shaping fungal communities both in the phyllosphere and in the rhizosphere of forest trees [5,42,47,48].
Fungal pathogens represent an important biotic factor that may negatively affect health and growth of forest trees [11]. In the present study, a fungal pathogen Sydowia polyspora was among the most dominant fungi (Table 3). Sydowia polyspora has a wide geographical range [48] and is common in Europe. The pathogenicity of S. polyspora to young conifers (genera Thuja, Abies, Tsuga, Larix, Picea and Pinus) was previously reported [49]. In the Balkan region, it has been reported in forest plantations of P. nigra and P. sylvestris in Serbia, occurring on needles damaged by drought or frost [50]. It was also detected on needles of P. halepensis in Italy and Spain [44,51], P. sylvestris in Poland and Lithuania [29,39] and on P. ponderosa in North America [52]. Sydowia polyspora has recently been reported as a pathogen that dominates fungal communities vectored by bark beetles associated with P. radiata, P. nigra subsp. salzmannii and P. sylvestris in Spain [53] and Pinus yunnanensis in China [54]. Sydowia symptoms include needle necrosis and shoot dieback. The fungus is favoured by a warm climate, especially if the host is stressed by summer drought or insect or mite attack [53]. Indeed, in the present study the highest relative abundance of S. polyspora was at the KKN (Table 2), the site that was subjected to insect attack. Besides, it appears that S. polyspora had a negative effect on the relative abundance of L. pinastri (Table 2), which is in agreement with Behnke-Borowczyk et al. [39], who have observed the similar pattern on P. sylvestris. Cyclaneusma niveum was also among the dominant fungi (Table 3). It is one of the two fungal pathogens causing Cyclaneusma needle cast, which is an important needle disease reported from many pine species including P. nigra and P. sylvestris in the Balkan region and in Crimea [50,55,56], and reported from P. halepensis in Spain [44]. It was suggested that C. niveum is more frequently associated with P. nigra under warmer climate conitions, while Cyclaneusma minus is favoured by wet, humid, above-freezing conditions, and thus, more commonly infects P. sylvestris grown in central and northern Europe [39,50,57]. Our study is in agreement, as on P. heldreichii, C. niveum dominated the fungal community, while C. minus was recorded at a low frequency (Table S1). Interestingly, C. niveum was more common on sites with prevailing moderate than harsh enviromental conditions. Neocatenulostroma germanicum is a recently identified fungal pathogen causing needle blight on P. mugo, P. sylvestris and P. nigra subsp. pallsiana in Lithuania, Ukraine and Poland, where it was found to be commonly associated with Dothistroma, Lecanosticta acicola and Cyclaneusma needle cast infections [39,58]. In this study, it was present in the needles of P. heldreichii from five sites, with considerably higher relative abundance on symptomatic needles at the KKN site and the neighbouring KKO site as compared to the remaining sites. Allantophomopsiella pseudotsugae (syn. Phomopsis pseudotsugae) was a pathogen commonly detected on needles of P. heldreichii grown on sites under harsh growth conditions (ORJ and KMT) (Table 3). While being a pathogen of conifers, that is, mainly infecting pines, it develops on young shoots [59] and has been reported in several European countries [60]. The present finding of the fungus represents a new record for this part of Europe, indicating that A. pseudotsugae could be among important pathogens of P. heldreichii growing under harsh conditions. Cenangium accum was recorded on P. heldreichii needles at the PRE site (Table 3). Previously it was demonstrated as a pathogen of weakened P. nigra and P. sylvestris in the Balkan region [50,55]. It has also been shown to be associated with Cyclaneusma needle cast in P. sylvestris in western Poland [39]. Cenangium accum develops predominantly on needles damaged by frosts or drought and its development is favoured by a high humidity. Similarly, Dothistroma septosporum, was recorded at a low relative abundance and only on the PRE (Table S1). Investigation of Dothistroma septosporum accomplished using PCR and species-specific primers, has demonstrated the presence of this potentially invasive pathogen across the P. heldreichii distribution range in Montenegro [17]. It was suggested that enviromental conditions present at P. heldreichii sites suppress the development of Dothistroma needle blight and that P. heldreichii is only slightly susceptible to D. septosporum. However, the infection level by D. septosporum may vary in different years, especially after rainy periods [10,17].
Among the other fungi detected, there were fungal endophytes that are known to be commonly associated with different tissues of forest trees [61,62]. The relationship between endophytic fungi and plants is not clearly understood and may change depending on the health status of the plant. However, for at least part of their life, they colonise plant tissues asymptomatically [12,63,64]. In the present study, L. pinastri and L. conigenum were the most commonly detected fungi on P. heldreichii needles. They are globaly distributed and commonly associated with pines [42,65]. It was shown recently that L. pinastri colonises healthy needles latently as an endophyte, initiates active growth at the begining of needle senescence and sporulates after needle fall. It is a dominant coloniser of dying needles and a saprotroph contributing to a needle decomposition [39]. In the Balkan region, L. pinastri was frequently reported from P. nigra and P. sylvestris grown in forests, forest nurseries and plantations [50,55]. Lophodermium conigenum is known as a coloniser of damaged needles and can also form fruitbodies on broken branches [65]. In comparison to L. pinastri, it is less frequently reported on both P. nigra and P. sylvestris in Serbia [50] and on P. sylvestris in Spain [45] and the UK [66]. In the present study, L. pinastri was present on all investigated sites, while L. conigenum showed a higher relative abundance on sites characterised by moderate growth conditions (PRO, KKO and PRE). The relative abundance of L. pinastri was at least 10 times lower on the KKN site than on any other site, what was likely due to its exclusion by pathogenic fungi which dominated on this site.
Some fungal endophytes of conifers can produce bioactive secondary metabolites, supporting ecological adaptations of host plants owing strong antimicrobial activities [63,67]. Microsphaeropsis olivacea (syn. Coniothyrium olivaceum) is a fungal endophyte, which was shown to produce bioactive compounds that are considered as promising antibacterial and antifungal agents [68]. Microsphaeropsis olivacea was among the most abundant species on the KKN, where Sydowia polyspora and Neocatenulostroma germanicum were also abundant. Mollisia ligni has been shown to produce mollisin, a compound that is known for a strong fungicidal activity against Sydowia polyspora [69]. A high frequency of Mollisia ligni may result in decreased frequency of S. polyspora on P. sylvestris [39]. On P. heldreichii, N. germanicum was absent in the cases where M. ligni was present (KMT).
The study has also detected a number of fungi previously known as pathogens of deciduous trees and/or of agricultural crops including Phaeomoniella 2814_15, Ramoconidiophora euphorbiae and Geastrumia sp., which were for the first time recorded on conifers, possibly as latent endophytes. Fungi from genus Phaeomoniella are important pathogens of grapevines, causing grapevine trunk disease [70], Ramoconidiophora and Collophorina species have been reported from necrotic and symptomless wood and leaves of Prunus, Castanea, Vitis, and from roots of Calluna [71], while Geastrumia sp. is known as one of the species involved in disease complex of sooty blotch and flyspeck of apple (Malus domestica) [72]. Phaeomoniella sp., Ramoconidiophora, Collophorina and Geastrumia were more abundant on sites with harsh growth conditions (ORJ and KMT). Among other pathogens, Lachnellula calyciformis was abundant on sites under harsh growing conditions, while Phaeosphaeria pontiformis was on sites characterised by moderate growing conditions. Corticoid fungus from genus Athelia, which typically occurs as saprotroph [73,74] was present on sites with moderate growth conditions.
Fungi may modulate stress tolerance, enhance growth and increase reproduction [7,47,75]. Habitats subjected to a high abiotic stress can be inhabited by a species-specific fungal community [76,77]. Our results demonstrated that needles of P. heldreichii constitute a habitat for a species-rich community of fungi, the composition of which was found to be largely driven by environmental conditions and/or health status of host trees. Indeed, similar patterns were also observed for fungal communities in roots of P. heldreichii [5], indicating that without the habitat-adapted fungal symbionts, plants are hardly capable of surviving in high stress habitats.
Fungal communities evolve together with host plants [12], suggesting that relic and endemic pine species such as P. heldreichii [5,54], can be associated with a specific fungal community. For example among the 50 mushroom species, which were recorded in P. heldreichii forests to date [78,79,80,81], many were rarely observed and exclusively recorded in P. heldreichii forests. Hence, Chalciporus ammarelus, Geastrum minimum, Hygroporus gliocyclus, Hygroporus hypothejus, Rhizopogon roseolus and Morchella esculenta s.l. are listed on the preliminary red list [79], and thus, are protected [82,83]. Furthermore, recently described species Erioscyphella curvispora [84] was discovered in P. heldreichii needle litter and a newly described genus Perzia, accommodated by a type species Perzia triseptata, gen. nov. was discovered on the xeric bark of P. heldreichii [85]. Other findings include newly described species Velutarina bertinscensis [86], Peziza montrivicola [87], Cenangiopsis ragvanii, C. junipericola [88] and rare species Trichophaea flavobrunea [89] that were determined on other tree species growing on P. heldreichii sites. Those findings represents a special value to science, and together with overall recorded fungal diversity demonstrate a high value of P. heldreichii habitats as being unique and biodiversity hotspots in high-altitude mountain areas.
In summary, the results demonstrated that needles of P. heldreichii were associated with a species-rich community of fungi, the composition of which was found to be largely dependent on environmental conditions and/or health status of host trees. Needle pathogens appeared to be an important component of fungal communities associated with needles of P. heldreichii, but caused limited damage likely due to interaction with other fungal species and/or due to plant defence mechanisms. In order not to become a threat to the health and growth of forest trees, the present and potentially new invasive pathogens should be monitored regularly. Further research is also needed to understand patterns of different fungal species coexistence.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/12/5/172/s1, Table S1: Relative abundance of fungal taxa detected in needles of Pinus heldreichii at six different sites in Montenegro, Table S2: Sørensen similarity index of the phyllosphere fungal communities among the six Pinus heldreichii sampling sites.
Author Contributions
Conceptualisation, J.L. and A.M.; methodology, J.L. and A.M.; formal analysis, J.L. and A.M.; resources, J.L.; data curation, J.L. and A.M.; writing—original draft preparation, J.L.; writing—review and editing, J.L. and A.M.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science of Montenegro through the grant INVO HERIC No: 01-646. A.M. was supported by the Swedish Research Council Formas (grant no. 2019-00597).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Janković, M.M. Betrachtungen uber gegenseitigen Bezichungen der Molika (Pinus peuce) und Panzerkiefer (Pinus heldreichii) sowie auch Uber ihre okologishen Eigenschaften, besonders in Bezug auf ihre geologishe Grundlage. Bull. L’Inst. Jard. Bot. L’univ. Beogr. 1960, 1, 141–181. [Google Scholar]
- Jovanović, B. Dendrology, 6th ed.; Faculty of Forestry, University of Belgrade: Belgrade, Serbia, 2007; p. 536. [Google Scholar]
- Blečić, V.; Lakušić, R. Forests of Pinius heldreichii Christ at Štitovo and Bjelasica in Montenegro. Bull. Repub. Inst. Prot. Nat. Mus. Nat. Hist. Titogr. 1969, 4, 5–10. [Google Scholar]
- Stevanović, V.; Jovanović, S.; Lakušić, D. Diversity of vegetation of Yugoslavia. In Biodiversity of Yugoslavia with List of Species of Special Importance; Radović, I., Angelus, J., Eds.; Yugoslavia: Ecolibri, Beograd, 1995; pp. 219–241. [Google Scholar]
- Lazarević, J.; Menkis, A. Fungi inhabiting fine roots of Pinus heldreichii in the Montenegrin montane forests. Symbiosis 2018, 74, 189–197. [Google Scholar] [CrossRef]
- Vendramin, G.G.; Fineschi, S.; Fady, B. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Bosnian Pine (Pinus heldreichii); Bioversity International: Rome, Italy, 2008; ISBN 978-92-9043-789-5. [Google Scholar]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. Fems Mycrobiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.E.; Kim, M.S.; Klopfenstein, N.B. Molecular genetic approaches toward understanding forest-associated fungi and their interactive roles within forest ecosystems. Curr. For. Rep. 2018, 4, 72–84. [Google Scholar] [CrossRef]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.K.; Crowther, T.W.; et al. Fungal functional ecology: Bringing a trait-based approach to plant-associated fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef]
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.; Vahalik, P.; Mullett, M.; Martín-García, J.; Bulman, L.; Wingfield, M.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Cacciola, S.O.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Mullett, M.; Drenkhan, T.; Oskay, F.; Aday Kaya, A.G.; et al. Potential Interactions between Invasive Fusarium circinatum and Other Pine Pathogens in Europe. Forests 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.J.; Redman, R.S. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest tree microbiomes and associated fungal endophytes: Functional roles and impact on forest health. Forests 2019, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Lazarević, J. Contribution to the study of physiological characteristics of the pathogenic fungus Herpotrichia juniperi. Agric. For. 2003, 49, 95–109. [Google Scholar]
- Lazarević, J. Pinus heldreichii as the host of brown felt blight (Herpotrichia juniperi) in Montenegro. Mycol. Monten. 2004, 7, 77–89. [Google Scholar]
- Lazarević, J. Contribution to the study of te morphological characteristics of the pathogenic fungus Herpotrichia juniperi. Mycol. Monten. 2005, 8, 137–147. [Google Scholar]
- Lazarević, J.; Davydenko, K.; Millberg, H. Dothistroma Needle Blight on High Altitude Pine Forests in Montenegro. Balt. For. 2017, 23, 294–302. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef]
- Dinaric_Alps_map-fr.svg: Sémhur, Relief Map of Montenegro svg. Available online: https://commons.wikimedia.org/wiki/File:Relief_Map_of_Montenegro.svg (accessed on 10 January 2020).
- Fuštić, B.; Đuretić, G. Soils of Montenegro; Biotechnical Institute, University of Montenegro: Podgorica, Montenegro, 2000; pp. 156–172. [Google Scholar]
- IUSS Working Group WRB. Descriptions, distribution, use and management of Reference Soil Groups. In World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, Udate 2015; World Soil Resource Reports No. 106; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; pp. 144–180. [Google Scholar]
- Burić, M.; Micev, B.; Mitrović, L. Atlas of Climate of Montenegro; Montenegrin Academy of Science and Arts: Podgorica, Montenegro, 2012; pp. 18–62, 86–102, 126–129. [Google Scholar]
- Burić, D.; Ducić, V.; Mihajlović, J. The climate of Montenegro: Modificators and types-part two. Bull. Serb. Geogr. Soc. 2014, 94, 73–90. [Google Scholar] [CrossRef] [Green Version]
- Janković, M.M.; Stefanović, K. Ecological relations between relict (sub)endemic pine species Pinus heldreichii and the character of geological substrate and soil in Jugoslavia. Ekologija 1971, 6, 49–61. [Google Scholar]
- Menkis, A.; Ihrmark, K.; Stenlid, J.; Vasaitis, R. Root-associated fungi of Rosa rugosa grown on the frontal dunes of the Baltic Sea coast in Lithuania. Microb. Ecol. 2014, 67, 769–774. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Menkis, A.; Marčiulynas, A.; Gedminas, A.; Lynikienė, J.; Povilaitienė, A. High-throughput sequencing reveals drastic changes in fungal communities in the phyllosphere of Norway spruce (Picea abies) following invasion of the spruce bud scale (Physokermes piceae). Microb. Ecol. 2015, 70, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Lynikienė, J.; Marčiulynienė, D.; Marčiulynas, A.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Managed and unmanaged Pinus sylvestris forest stands harbour similar diversity and composition of the phyllosphere and soil fungi. Microorganisms 2020, 8, 259. [Google Scholar] [CrossRef] [Green Version]
- Brandström Durling, M.; Clemmensen, K.; Stenlid, J.; Lindahl, B. SCATA—An Efficient Bioinformatic Pipeline for Species Identification and Quantification after High-Throughput Sequencing of Tagged Amplicons (Submitted). 2011. Available online: https://www.scata.mykopat.slu.se (accessed on 12 November 2019).
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [PubMed] [Green Version]
- Analytic Rarefaction, UGA Stratigraphy Lab. Available online: http://www.uga.edu/strata/software/index.html (accessed on 10 January 2020).
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; p. 192. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; H. Freeman & Company: New York, NY, USA, 2005; p. 896. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination, Version 4; Microcomputer Power: Ithaca, NY, USA, 1998; p. 351. [Google Scholar]
- Bringel, F.; Couee, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porazinska, D.L.; Sung, W.; Giblin-Davis, R.M.; Thomas, W.K. Reproducibility of read numbers in high-throughput sequencing analysis of nematode community composition and structure. Mol. Ecol. Resour. 2010, 10, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Behnke-Borowczyk, J.; Kwaśna, H.; Kulawinek, B. Fungi associated with Cyclaneusma needle cast in Scots pine in the west of Poland. For. Pathol. 2019, 49, e12487. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers—A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar]
- Tedersoo, L.; Tooming-Klunderud, A.; Anslan, S. PacBio metabarcoding of Fungi and other eukaryotes: Errors, biases and perspectives. New Phytol. 2018, 217, 1370–1385. [Google Scholar]
- Millberg, H.; Boberg, J.; Stenid, J. Changes in fungal community of Scots pine (Pinus sylvestris) needles along a latitudinal gradient in Sweden. Fungal Ecol. 2015, 17, 126–139. [Google Scholar] [CrossRef]
- Oono, R.; Lefèvre, E.; Simha, A.; Lutzoni, F. A comparison of the community diversity of foliar fungal endophytes between seedling and adult loblolly pines (Pinus taeda). Fungal Biol. 2015, 119, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Botella, L.; Santamaria, O.; Diez, J.J. Fungi associated with decline of Pinus halepensis in Spain. Fungal Divers. 2010, 40, 1–11. [Google Scholar] [CrossRef]
- Sanz-Ros, A.V.; Muller, M.M.; San Martin, R.; Diez, J.J. Fungal endophytic communities on twigs of fast and slow growing Scots pine (Pinus sylvestris L.) in northern Spain. Fungal Biol. 2015, 119, 870–883. [Google Scholar] [CrossRef]
- Parfitt, D.; Hunt, J.; Dockrell, D.; Rogers, H.J.; Boddy, L. Do all trees carry the seeds of their own destruction? PCR reveals numerous wood decay fungi latently present in sapwood of a wide range of angiosperm trees. Fungal Ecol. 2010, 3, 338–346. [Google Scholar] [CrossRef]
- Bowman, E.A.; Arnold, A.E. Distribution of ectomicorrhizal and foliar endophytic fungal communities associated with Pinus ponderosa along a spatially constrained elevation gradient. Am. J. Bot. 2018, 105, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Botella, L.; Diez, J.J. Phylogenetic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar]
- Talgo, V.; Chastanger, G.; Thomsen, I.M.; Cech, T.; Riley, K.; Lang, K.; Klemsdal, S.S. Sydowia polyspora associated with current season needle necrosis (CSNN) on true fir (Abies spp.). Fungal Biol. 2010, 114, 545. [Google Scholar] [CrossRef] [PubMed]
- Karadžić, D.; Milijašević, T. The most important parasitic and saprophytic fungi in Austrian pine and Scots pine plantations in Serbia. Bull. Fac. For. 2008, 97, 147–170. [Google Scholar]
- Tinivella, F.; Dani, E.; Minuto, G.; Minuto, A. First Report of Sydowia polyspora on Aleppo Pine (Pinus halepensis) in Italy. Plant Dis. 2014, 98, 2. [Google Scholar] [CrossRef]
- Ridout, M.; Newcombe, G. Sydowia polyspora is both a Foliar Endophyte and a Preemergent Seed Pathogen in Pinus Ponderosa. Plant Dis. 2018, 118, 3. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Adalia, E.J.; Sanz-Ros, A.V.; Flores-Pacheco, J.A.; Hantula, J.; Diez, J.J.; Vainio, E.J.; Fernández, M. Sydowia polyspora Dominates Fungal Communities Carried by Two Tomicus Species in Pine Plantations Threatened by Fusarium circinatum. Forests 2017, 8, 127. [Google Scholar] [CrossRef]
- Pan, Y.; Ye, H.; Lu, J.; Chen, P.; Zhou, X.D.; Qiao, M.; Yu, Z.-F. Isolation and identification of Sydowia polyspora and its pathogenicity on Pinus yunnanensis in Southwestern China. J. Phytopatol. 2018, 166, 386–395. [Google Scholar] [CrossRef]
- Dobreva, M.; Georgieva, M.; Dermedzhiev, P.; Nachev, R.; Velinov, V.; Terziev, P.; Georgiev, G. Fungal pathogens associated with Pinus species in the region of forest protection station Plovdiv in the period 2013–2016. For. Sci. 2016, 1–2. [Google Scholar]
- Minter, D.W.; Dudka, I.O. Fungi of Ukraine. A Preliminary Checklist; International Mycological Institute: Egham, Surrey, UK; M.G. Kholodny Institute of Botany: Kiev, Ukraine, 1996; pp. 1–361.
- Drenkhan, R.; Hanso, M. Recent invasion of foliage fungi of pines (Pinus spp.) to the Northern Baltics. For. Stud. 2009, 51, 49–64. [Google Scholar] [CrossRef]
- Markovskaja, S.; Kačergius, A.; Davydenko, K.; Fraser, S. First record of Neocatenulostroma germanicum on pines in Lithuania and Ukraine and its co- occurrence with Dothistroma spp. and other pathogens. Path 2016, 46, 522–533. [Google Scholar] [CrossRef]
- Wajihi, A.H.; Lee, S.; Das, K.; Eom, A.; Jung, H. First Report of Allantophomopsiella pseudotsugae Isolated from Soil in Korea. Korean J. Mycol. 2019, 47, 29–34. [Google Scholar] [CrossRef]
- Crous, P.W.; Quaedvlieg, W.; Hansen, K.; Hawksworth, D.L.; Groenewald, J.Z. Phacidium and Ceuthospora (Phacidiaceae) are congeneric: Taxonomic and nomenclatural implications. Ima Fungus 2014, 5, 173–193. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Kogel, K.-H.; Franken, P.; Huckelhoven, R. Endophyte or parasite-what decides. Curr. Opin. Plant Biol. 2006, 9, 358–369. [Google Scholar] [CrossRef]
- Soltani, J.; Hosseyni Moghaddam, M.S. Fungal Endophyte Diversity and Bioactivity in the Mediterranean Cypress Cupressus Sempervirens. Curr. Microbiol. 2015, 70, 580–586. [Google Scholar]
- Jeewon, R.; Yeung, Q.S.; Wannasinghe, D.N.; Rampadarath, S.; Puchooa, D.; Wang, H.K.; Hyde, K.D. Hidden mycota of pine needles: Molecular signatures from PCRDGGE and Ribosomal DNA phylogenetic characterization of novel phylotypes. Sci. Rep. 2018, 8, 18053. [Google Scholar] [CrossRef] [Green Version]
- Minter, D.W.; Staley, J.M.; Millar, C.S. Four species of Lophodermium on Pinus sylvestris. Trans. Br. Mycol. Soc. 1978, 71, 295–301. [Google Scholar]
- Reignoux, S.A.; Green, S.; Ennos, R.A. Molecular identification and relative abundance of cryptic Lophodermium species in natural populations of Scots pine, Pinus sylvestris L. Fungal Biol. 2014, 118, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.A.; Stierle, D.B. Bioactive secondary metabolites produced by fungal Endophytes of conifers. Nat. Prod. Commun. 2015, 10, 1671–1682. [Google Scholar] [PubMed] [Green Version]
- Wang, L.W.; Xu, B.G.; Wang, J.Y.; Su, Z.Z.; Lin, F.C.; Zhang, C.L.; Kubicek, C.P. Bioactive metabolites from Phoma species, an endophytic fungus from the Chinese medicinal plant Arisaema erubescens. Appl. Microbiol. Biotechnol. 2012, 93, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Johne, S.; Watzke, R.; Meusel, W.; Mollmann, V.; Harti, A.; Dahse, H.M.; Matthes, B.; Seifert, K. Biotechnological production and bioactivities of mollisin and two new, structuraly related fungal naphthequinone metabolites. Chem. Biodivers. 2005, 2, 1109–1115. [Google Scholar] [PubMed]
- Bertscha, C.; Ramırez-Sueroa, M.; Magnin-Robertb, M.; Larignonc, P.; Chonga, J.; Abou-Mansourd, E.; Spagnolob, A.; Clementb, C.; Fontaineb, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Bien, S.; Kraus, C.; Damm, U. Novel Collophorina-like genera and species from Prunus trees and vineyards in Germany. Persoonia 2020, 45, 46–67. [Google Scholar] [CrossRef]
- Piperkova, N.; Yonkova, I. Symptoms, Etiology and Control of Sooty Blotch and Flyspeck in Bulgaria. Turk. J. Agric. Nat. Sci. 2014, 1, 817–822. [Google Scholar]
- Epidemiology of the Fungus Athelia arachnoidea in Epiphytic Communities of Broadleaved Forests under Strong Anthropogenic Impact. Available online: http://www.elibrary.lt/resursai/LMA/Ekologija/0504_07_Eko.pdf (accessed on 5 March 2020).
- Adams, G.C.; Kropp, B.R. Athelia arachnoidea, the sexual state of Rhizoctonia carotae, a pathogen of carrot in cold storage. Mycologia 1996, 88, 459–472. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar]
- Gehring, C.A.; Theimer, T.C.; Whitham, G.; Keim, P. Ectomycorrhizal fungal community structure of pinyon pines growing in two environmental extremes. Ecology 1998, 79, 1562–1572. [Google Scholar]
- Rosa, L.H.; Vaz, A.B.M.; Caligiorne, R.B.; Campolina, S.; Rosa, C.A. Endophytic fungi associaed with Antarctic grass Deschampsia antarctica Devs (Poaceae). Polar Biol. 2009, 32, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Perić, B.; Perić, O. Diversity of macromycetes in Montenegro. MASA 1997, 11, 45–142. [Google Scholar]
- Perić, B.; Perić, O. Preliminary red list of Macromycetes of Montenegro 2. Mycol. Monten. 2004, 7, 7–33. [Google Scholar]
- Lazarević, J.; Perić, O.; Perić, B. Ectomycorrhizal fungi in Montenegro—Diversity and distribution. Mycol. Monten. 2011, 14, 85–115. [Google Scholar]
- Lazarević, J. Forests and biodiversity of Kuči mountains. In Katun of the Kuči Mountains; Laković, I., Ed.; University of Montenegro: Podgorica, Montenegro, 2017; pp. 64–78. [Google Scholar]
- Official Gazette of Montenegro, No 054/16, 15.08.2016. Law of Nature Protection of Montenegro. Available online: http://www.sluzbenilist.me/pregled-dokumenta/?id={0C659042-9DD7-43FB-8E8C-B7DF445298A1} (accessed on 23 March 2020).
- Official Gazette of Montenegro, No 76/06, 12.12.2006, Act on Protection of Certain Plant and Animal Species. 2006. Available online: http://www.sluzbenilist.me/pregled-dokumenta/?id={631C3E5D-4129-4985-B55B-CE4D3703CA2E} (accessed on 23 March 2020).
- Perić, B.; Baral, H.-O. Erioscyphella curvispora, spec. nov. from Montenegro. Mycol. Monten. 2015, 17, 89–104. [Google Scholar]
- Baral, H.-O.; Perić, B. Perzia triseptata Gen. et sp. nov. (Ascomycota, Insertae sedis) from xeric bark in France and Montenegro. Mycol. Monten. 2016, 19, 7–20. [Google Scholar]
- Baral, H.-O.; Perić, B. Velutarina bertiscensis and V. alpestris spp. nov., with a redescription of V. rufoolivacea and discussion of its synonyms. Mycol. Monten. 2015, 17, 17–52. [Google Scholar]
- Perić, B.; Grebenc, T. Une espèce nouvelle du genre Peziza (Pezizales): P. ontirivicola spec. Nov. Ascomycete. Org. 2015, 7, 347–356. [Google Scholar]
- Perić, B.; Baral, H.-O.; Partel, K. Cenangiopsis raghavanii and C. junipericola spp. nov (Cenangiaceae, Helotiales) collected in Montenegro, with redescription of a recent collection of C. quercicola. Mycol. Monten. 2015, 17, 7–40. [Google Scholar]
- Perić, B.; van Vooren, N.; Healy, R.; Lazarević, J. Une Trichophaea rare récoltée en France et au Monténégro: T. Flavobrunnea Comb. Nov. (Pezizales). Mycol. Monten. 2014, 16, 65–87. [Google Scholar]
Figure 1.
A map showing (a) distribution of Pinus heldreichii in the Mediterranean region; a grey area was added and represents an additional P. heldreichii site; the map is adapted from [18] and (b) sampling sites in Montenegro (ORJ-Orjen; PRE-Prekornica; KKO-Kuči Mt, Kučka korita; KKN-Kuči Mt., Kučka korita North, KMT- Kuči mountain, Momonjevo, PRO- Prokletije); a map was adapted from [19].
Figure 1.
A map showing (a) distribution of Pinus heldreichii in the Mediterranean region; a grey area was added and represents an additional P. heldreichii site; the map is adapted from [18] and (b) sampling sites in Montenegro (ORJ-Orjen; PRE-Prekornica; KKO-Kuči Mt, Kučka korita; KKN-Kuči Mt., Kučka korita North, KMT- Kuči mountain, Momonjevo, PRO- Prokletije); a map was adapted from [19].

Figure 2.
(a) Pinus heldreichii shoots with needles; trees growing at (b) Kuči Mountains, Momonjevo (KMT), and (c) Kuči Mountains, Kučka korita (KKO) sites.
Figure 2.
(a) Pinus heldreichii shoots with needles; trees growing at (b) Kuči Mountains, Momonjevo (KMT), and (c) Kuči Mountains, Kučka korita (KKO) sites.

Figure 3.
Rarefaction curves showing the relationship between the cumulative number of fungal taxa and the number of ITS2 rDNA sequences from needles of Pinus heldreichii from six different sites.
Figure 3.
Rarefaction curves showing the relationship between the cumulative number of fungal taxa and the number of ITS2 rDNA sequences from needles of Pinus heldreichii from six different sites.

Figure 4.
Relative abundance of different fungal classes in needles of Pinus heldreichii from six different sites in Montenegro. Classes comprising <1.0% of relative abundance for a given set of sequences were combined and shown as other. Sites are as in Table 1.
Figure 4.
Relative abundance of different fungal classes in needles of Pinus heldreichii from six different sites in Montenegro. Classes comprising <1.0% of relative abundance for a given set of sequences were combined and shown as other. Sites are as in Table 1.

Figure 5.
Ordination diagram based on nonmetric multidimensional scaling (NMDS) of fungal communities associated with needles of Pinus heldreichii from six different sites in Montenegro, with 56.5% variation on Axis 1 and 28.2% on Axis 2. Each point in the diagram represents a single site and the size of the point reflects richness of the fungal taxa as in Table 2. Sites are as in Table 1.
Figure 5.
Ordination diagram based on nonmetric multidimensional scaling (NMDS) of fungal communities associated with needles of Pinus heldreichii from six different sites in Montenegro, with 56.5% variation on Axis 1 and 28.2% on Axis 2. Each point in the diagram represents a single site and the size of the point reflects richness of the fungal taxa as in Table 2. Sites are as in Table 1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of Pinus heldreichii sampling sites.
| Site | GPS Coordinates | Altitude (m) | Soil Type a | Climate b | Health c |
|---|---|---|---|---|---|
| Orjen, Reovačka greda (ORJ) | 42°35′ N 18°35′ E | 1700 | Lithosol | Cfsb | Healthy-looking |
| Kuči Mt., Momonjevo (KMT) | 42°36′ N 19°32′ E | 1800 | Lithosol | Cfb | Healthy-looking |
| Prekornica, Studeno (PRE) | 42°38′ N 19°12′ E | 1200 | Leptic cambisol | Cfsb | Healthy-looking |
| Prokletije, Ropojana (PRO) | 42°29′ N 19°48′ E | 1250 | Leptic cambisol | Cfb | Healthy-looking |
| Kučka korita (KKO) | 42°29′ N 19°30′ E | 1300 | Leptic cambisol | Cfs’’b | Healthy-looking |
| Kučka korita North (KKN) | 42°30′ N 19°32′ E | 1450 | Molic leptosol | Cfs’’b | Attacked by insects |
Table 2.
Generated high-quality ITS2 rDNA fungal sequences and detected diversity of fungal taxa at different sampling sites of Pinus heldreichii. Within the column No. of Fungal Taxa, values followed by the same letter in chi-square test do not differ significantly at p > 0.05.
Table 2.
Generated high-quality ITS2 rDNA fungal sequences and detected diversity of fungal taxa at different sampling sites of Pinus heldreichii. Within the column No. of Fungal Taxa, values followed by the same letter in chi-square test do not differ significantly at p > 0.05.
| Site | No. of Fungal Sequences | No. of Fungal Taxa | Shannon Diversity Index |
|---|---|---|---|
| Orjen –ORJ | 4830 | 139 a | 3.4 |
| Kuči Mt.-KMT | 4910 | 119 ab | 2.9 |
| Prekornica-PRE | 5828 | 192 a | 3.1 |
| Prokletije -PRO | 3679 | 127 a | 3.2 |
| Kučka korita -KKO | 8748 | 167 b | 2.9 |
| Kučka korita North –KKN | 3834 | 104 ab | 2.7 |
| All | 31,829 | 375 | - |
Table 3.
Relative abundance of the 25 most common fungal taxa detected in needles of Pinus heldreichii at six sites in Montenegro. Sites are as in Table 1.
Table 3.
Relative abundance of the 25 most common fungal taxa detected in needles of Pinus heldreichii at six sites in Montenegro. Sites are as in Table 1.
| Fungal Taxon | Phylum * | GenBank Accession No. | Compared (bp) | Similarity (%) | Sites | All | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ORJ | KMT | PRE | PRO | KKO | KKN | ||||||
| Lophodermium pinastri | A | KC608049 | 361/361 | 100 | 4.7 | 22.1 | 14.3 | 14.2 | 14.9 | 0.4 | 12.5 |
| Lophodermium conigenum | A | HM060650 | 453/453 | 100 | - | 0.1 | 32.4 | 5.5 | 14.5 | 2.8 | 10.9 |
| Sydowia polyspora | A | KU516591 | 473/473 | 100 | 6.8 | 16.0 | 2.1 | 8.2 | 0.4 | 32.2 | 8.8 |
| Cyclaneusma niveum | A | AF013223 | 442/442 | 99 | 1.3 | 1.0 | 4.5 | 5.7 | 12.9 | 0.9 | 5.5 |
| Unidentified sp. 2814_1 | A | MF976656 | 238/245 | 97 | 20.2 | - | 0.5 | - | 8.2 | - | 5.4 |
| Phaeosphaeria pontiformis | A | KT000144 | 442/442 | 98 | - | - | 3.7 | 11.1 | 8.9 | - | 4.4 |
| Unidentified sp. 2814_11 | A | KU062806 | 203/252 | 81 | - | - | 1.3 | 9.0 | 10.3 | - | 4.1 |
| Unidentified sp. 2814_10 | A | KX220267 | 257/257 | 100 | 3.3 | - | 1.5 | 4.1 | 5.4 | 5.4 | 3.4 |
| Phaeomoniella sp. 2814_15 | A | GQ153187 | 257/259 | 99 | 7.2 | 5.9 | 0.2 | 1.2 | 2.9 | 0.5 | 3.0 |
| Neocatenulostroma germanicum | A | KR995100 | 242/242 | 100 | 1.3 | - | 0.4 | 1.0 | 3.4 | 7.9 | 2.3 |
| Microsphaeropsis olivacea | A | MH871969 | 249/249 | 100 | 0.7 | 0.1 | 0.3 | 0.1 | - | 16.8 | 2.2 |
| Allantophomopsiella pseudotsugae | A | MH857222 | 240/240 | 100 | 6.4 | 7.1 | - | - | - | - | 2.1 |
| Lachnellula calyciformis | A | MH858771 | 239/239 | 100 | 4.7 | 8.1 | - | - | - | - | 2.0 |
| Unidentified sp. 2814_23 | A | KF983527 | 209/244 | 86 | 1.4 | - | 1.5 | 3.7 | 0.3 | 4.3 | 1.5 |
| Ramoconidiophora euphorbiae | A | MG592740 | 232/239 | 97 | 3.2 | 6.0 | - | - | - | 0.6 | 1.5 |
| Geastrumia sp. 2814_26 | A | FJ438389 | 220/235 | 94 | 2.7 | 2.6 | 0.4 | 0.6 | 0.5 | 2.3 | 1.4 |
| Unidentified sp. 2814_16 | A | KP892077 | 240/242 | 99 | 4.8 | 0.1 | 0.6 | 1.9 | 0.3 | 0.5 | 1.2 |
| Unidentified sp. 2814_27 | A | KT244857 | 209/248 | 84 | - | - | 3.5 | 5.0 | - | - | 1.2 |
| Collophora sp. 2814_30 | A | NR_137726 | 229/241 | 95 | 3.2 | 2.2 | 0.4 | - | - | 0.3 | 0.9 |
| Unidentified sp. 2814_29 | A | MF976139 | 209/240 | 87 | 1.4 | - | 0.4 | 4.1 | 0.4 | - | 0.9 |
| Rhytismataceae sp. 2814_22 | A | KR266446 | 234/237 | 99 | 0.4 | - | 1.0 | 1.4 | 1.6 | - | 0.8 |
| Dothideomycetes sp. 2814_13 | A | KP991484 | 253/257 | 98 | 1.1 | 3.5 | - | 0.1 | - | 0.4 | 0.8 |
| Mollisia ligni | A | MF161301 | 237/241 | 98 | - | 4.7 | - | - | - | - | 0.7 |
| Cenangium acuum | A | MG597445 | 239/239 | 100 | - | - | 3.9 | - | - | - | 0.7 |
| Athelia acrospora | B | KP814375 | 296/299 | 99 | - | - | 1.8 | 0.1 | 1.3 | - | 0.7 |
| All of 25 taxa | 74.9 | 79.9 | 74.6 | 76.6 | 86.4 | 75.4 | 79.0 | ||||
* A—Ascomycota, B—Basidiomycota.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lazarević, J.; Menkis, A. Fungal Diversity in the Phyllosphere of Pinus heldreichii H. Christ—An Endemic and High-Altitude Pine of the Mediterranean Region. Diversity 2020, 12, 172. https://doi.org/10.3390/d12050172
AMA Style
Lazarević J, Menkis A. Fungal Diversity in the Phyllosphere of Pinus heldreichii H. Christ—An Endemic and High-Altitude Pine of the Mediterranean Region. Diversity. 2020; 12(5):172. https://doi.org/10.3390/d12050172
Chicago/Turabian StyleLazarević, Jelena, and Audrius Menkis. 2020. "Fungal Diversity in the Phyllosphere of Pinus heldreichii H. Christ—An Endemic and High-Altitude Pine of the Mediterranean Region" Diversity 12, no. 5: 172. https://doi.org/10.3390/d12050172
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.