
Abstract
Particularly after disturbance events, the early successional colonist Betula pendula Roth is experiencing renewed silvicultural interest with respect to the natural regeneration of large disturbed forest areas. In a case study, we therefore studied the seed dispersal of B. pendula from two adjacent spruce stands to large storm-felled sites at high altitudes in the Thuringian Forest (Germany) over a 2-year period. We applied inverse modelling to describe the distance-dependent seed distribution using a negative exponential kernel and seed production function of birch seed trees. Maximum seed numbers of 2015 n m−2 (non-mast year) and 9557 n m−2 (medium year) occurred within 40‒50 m distance to a seed tree. The predicted seed production rate of a birch seed tree with a reference dbh of 20 cm was approximately 350,000 n tree−1 (non-mast year) and 1,500,000 n tree−1 (medium year). Regardless of the seed crop, the dispersal distances were similar in both years. The isotropic model showed mean dispersal distances of 86 and 97 m (uphill) and 367 and 380 m (downhill) for the 2 years of seed sampling. No directionality in seed dispersal was found. The findings showed birch seed dispersal to be strongly influenced by site inclination, seed tree position (valley, slope or plateau) and distance to the storm-felled site. Furthermore, the seed shadow is influenced by the number of seed sources. Therefore, risk-adapted forest management should include the ‘spatial optimization’ of birch seed trees, ideally creating a network of small seed tree groups scattered more or less regularly within pure conifer forests.
Similar content being viewed by others


Introduction
As an anemochorously dispersed pioneer tree species with a wide natural range throughout Eurasia, silver birch (Betula pendula Roth) has a high ecological value within temperate and boreal forest ecosystems (Atkinson 1992; Hynynen et al. 2010). Silver birches enhance soil nutrition and soil stability, provide watershed protection, act as structural elements with a long-term stabilizing effect, and provide habitats and food for many organisms (see Patterson 1993; Humphrey et al. 1998; Ferris and Humphrey 1999; Priha 1999; Beck et al. 2016). In some European countries, like England, Sweden, Finland and Latvia, silver birch is the most important broadleaved tree species for timber production, plywood or veneer production (Cameron 1996; Luostarinen and Verkasalo 2000; Hynynen et al. 2010). Within their natural geographical range in Europe, birch species were often considered as a forest weed and, therefore, rigorously thinned out of forest stands during the last century (Röhrig and Gussone 1990; Koski and Rousi 2005). However, silver birch recently received renewed interest with respect to forest management at higher altitudes. The importance of silver birch in forest management has been increasing because of (a) the species’ ability to promptly and extensively recolonize disturbed sites due to its high annual seed production and its fast juvenile growth, even in open areas with extreme climatic conditions (Perala and Alm 1990; Atkinson 1992; Zerbe 2001; Hynynen et al. 2010), and (b) the heightened risk of catastrophic events in central European spruce forests. The small winged nuts (1.5‒2.0 mm) of birch are mainly dispersed by wind between June and November (Brouwer and Stählin 1975; Huth 2009). Pioneer forests composed of birches are able to quickly close water and nutrient cycles, thus soon creating a forest climate appropriate for the establishment of climax tree species (Zerbe 2009).
Therefore, empirical information about the seed production, seed dispersal distances, and deposited seed numbers of silver birch is required to establish ‘precautionary’ forest management systems that anticipate the high risk of catastrophic events, particularly in mountain spruce forests, and to ensure successful birch regeneration on disturbed sites. An approximate knowledge about the number of seeds per seed tree and the quality of the seed crop in a particular year is important to forecast the probability of natural regeneration in disturbed areas. The amount of seeds produced by an individual tree is influenced by, e.g. tree stem and crown dimension, vitality and age (Sarvas 1948, 1952; Moles et al. 2004) as well as by weather-dependent pollination and flowering success (Sarvas 1952). The allometric relationships between growth parameters and seed production per tree have been proven (Grisez 1975; Greene et al. 2004; Huth 2009; Da Ponte Canova 2018). Seed dispersal is an important driver for species movement, site colonization and the restoration of treeless or disturbed areas (Skarpaas et al. 2006), but can also produce limitations in spatial distribution of seeds (Daniels 2001; Karlsson 2001; Huth 2009).
It is generally difficult to determine the amount of produced seeds per tree and to study the process of seed dispersal (van Putten et al. 2012). For an exact determination of seeds per individual seed tree, cuttings would be necessary to count the seed numbers. For non-destructive methods, seeds must be marked or genetically analysed to link dispersed seeds to a specific seed source (van Putten et al. 2012; Nathan and Muller-Landau 2000; Tiebel et al. 2019). Therefore, phenomenological models are an appropriate statistical tool to predict seed dispersal, directionality and seed production on the basis of trapped seeds (Clark et al. 1998; van Putten et al. 2012). Huth (2009), who also used inverse modelling, reported mean seed dispersal distances (MDD) of 37 to 90 m for admixed silver birch trees within closed Norway spruce forests and seed production numbers of 0.18–7.3 million seeds per tree and year. Other studies analysing the seed dispersal distances of B. alleghaniensis reported mean dispersal distances between 26.9 and 37.1 m (Clark et al. 1998; Greene et al. 2004). However, most studies on aspects of birch seed distribution have been conducted within closed forest stands (e.g. Skoglund and Verwijst 1989; Houle and Payette 1990; Graber and Leak 1992; Leder 1992; Houle 1998; Wagner et al. 2004; Huth 2009). Only few studies focused on the seed dispersal of birch in open areas or large gaps (Bjorkbom 1971; Hughes and Fahey 1988; Greene and Johnson 1996; Karlsson 2001).
As shown in different wind tunnel experiments, experimental results under controlled conditions cannot easily be transferred to real field conditions (Augspurger and Franson 1987; Johnson and West 1988, cited in Bakker et al. 1996; Kadereit and Leins 1988; van Dorp et al. 1996; Greene and Johnson 1997). This lack of transferability is caused by highly complex and variable environmental factors related to field conditions (e.g. wind conditions, site surface relief and ground vegetation cover or seed characteristics) (Fenner 1985; Okubo and Levin 1989; Skarpaas et al. 2006).Therefore, reliable information about the temporal and spatial patterns of seed rain in open areas is needed to assess the natural regeneration potential of B. pendula seedlings and to develop recolonization and restoration management strategies (see Zhao et al. 2016; Holmström et al. 2017). This applies particularly to the large wind thrown forest areas in central Europe, which were created by the storm events of the last decades (Gregow et al. 2017) and will probably become more and more frequent with progressing climate change (Mölter et al. 2016).
The aim of this case study was to generate new knowledge about the dependence of seed distribution patterns on wind thrown sites on seed tree position in surrounding forest stands, seed tree numbers, seed crops, wind direction and topography. We observed the seed dispersal of B. pendula in 2015 and 2016 at two wind thrown forest sites at high altitudes (715‒775 m a.s.l.) in Thuringia, Germany. (1) We used inverse modelling to predict mean dispersal distance, directionality and seed crop per seed tree of B. pendula, based on seed trapping data. (2) The dependence of predicted seed distribution patterns on relief inclination, wind direction, seed production per tree, seed tree number around the studied storm-felled sites as well as the position of the seed tree (valley, slope or plateau) could not be explained by inverse models. Therefore, we discussed these factors and their influence on the observed and predicted seed densities in the discussion section. Finally, (3) we used simulations to spatially optimize the positioning of the seed trees in relation to the studied sites with regard to optimal seed distribution in the open areas.
Materials and methods
Study area
The study area is located at high elevations and along the ridges of the Thuringian Forest, a mountain range in the federal state of Thuringia, Germany (50° 40′ N and 10° 45′ E). The area is situated between 400 and 982 m above sea level (a.s.l.), with a prevailing south-westerly exposition. The area is characterized by many slopes and an almost total absence of plateaus (Burse et al. 1997; Waesch 2003; Gauer and Aldinger 2005). The mean annual precipitation ranges from 800 mm in the south-west to 1200 mm along the ridges and falls to a level of 700 mm in the north-east (Burse et al. 1997; Gauer and Aldinger 2005; Bushart and Suck 2008). The annual average temperature in the region varies between 4 and 6 °C (Burse et al. 1997; Bushart and Suck 2008). The area is influenced by an Atlantic, moderately cool and moist central mountain climate (Burse et al. 1997; Gauer and Aldinger 2005). The prevailing winds are from the south-west, with a secondary wind maximum originating from the north-east. The average annual wind speed in the study area is 3.5‒4.5 m s−1 (Bürger 2003). The averaged meteorological data (based on half-hourly values) for the seed trapping periods of the presented study (2015 and 2016) are listed in Table 1. While no extreme events in wind speed were observed, the wind direction showed a high variability during the study periods.
The dominant soil types of the forest sites are low-base cambisols with low to medium nutrient contents (Gauer and Aldinger 2005). The regional landscape features a largely contiguous forest system with ~ 90% forest cover, some small upland meadows in stream valleys and occasional small raised bogs. The study area is dominated by single-layered, even-aged Norway spruce forests (Picea abies (L.) Karst.). Without anthropogenic influence, the potential natural vegetation would be dominated by Luzulo-Fagetum and Asperulo-Fagetum beech forests (Frischbier et al. 2014).
We selected two study sites (B and E) 6 km apart from each other, located on slopes at higher elevations of the Thuringian Forest (715‒775 m a.s.l.). Each site consisted of an open area surrounded by a forested search zone of 200 m (see 2.2). The open areas were wind thrown by the storm ‘Kyrill’ in January 2007 (Fink et al. 2009). Representative for the region, the stand conditions before the storm were dominated by 68‒100 year-old Norway spruce. After the storm, the damaged areas were completely cleared, and no birch seed trees were present in the open areas. The size of the open areas was 4.0 ha and 12.7 ha, respectively (Table 2), and no closed regeneration layer had yet established at any of the study sites. The open areas were surrounded by 59‒105 year-old Norway spruce forests admixed with a small number of adult isolated Betula pendula Roth, Salix caprea L. and Sorbus aucuparia L. trees.
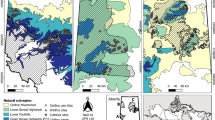
Located along slopes, the choice of study sites allowed us to separately investigate uphill (site B) and downhill (site E) seed dispersal. The seed trees at site B were located in the valley at approximately 710‒730 m a.s.l., and they were equipped with seed traps from the seed sources all the way to the uphill plateau at 760 m a.s.l. (Figure 1). At site E, seed trees were mainly found on a plateau (785‒805 m a.s.l.) within a stand consisting of Salix caprea, Sorbus aucuparia and B. pendula, and seed traps were placed close to the seed sources on the upper slope (775 m a.s.l.) and downhill along the slope to the valley (675 m a.s.l.).

Maps of the experimental study design at sites B (top) and E (bottom)
Sampling design
Within the 200 m forested search zone at each study site, we mapped all B. pendula trees that were expected to potentially produce seeds (≥ 12 cm diameter at breast height [dbh]; see Popadyuk et al. 1995; Roloff and Pietzarka 2010) using a blumax Bluetooth GPS-4013 Receiver. For each B. pendula tree, we recorded the dbh and observed flowering in both years. The search zone distance of 200 m was chosen as a compromise between feasibility and prior knowledge of suggested effective birch seed dispersal distance in open and forested sites (Sarvas 1948; Karlsson 2001; Huth 2009). Sixteen and 83 seed trees with 24‒42 cm and 13‒37 cm in dbh were identified at sites B and E, respectively (Table 2).
At the study sites, we placed 54 (site B) and 41 seed traps (site E). Due to the vast areal extent of the open areas, seed traps were placed along two crossing line transects (site E) and four crossing line transects (site B), with intervals of 20 m between the traps, rather than along a regular grid (Fig. 1; see also Bjorkbom 1971; Greene and Johnson 1996). The orientation and length of the line transects were not uniform, due to the differences in the size and shape of the two open areas. The line transects extended over the entire open area of each study site and into the surrounding Norway spruce forests. The minimum distances between the seed trees and the nearest seed trap were 12 and 74 m (Table 2).
The funnel-shaped seed traps had a diameter of 0.5 m and surface area of 0.196 m2. To ensure the functioning of the seed traps despite strong winds, a perforated plastic cup weighted with a stone was placed into each funnel-shaped net. The percolated plastic cups allowed rain water to runoff. The net funnels were fixed onto a bar 1 m above the ground. The traps were emptied periodically every 3 to 4 weeks and the number of seeds per trap was counted. The seed dispersal sampling periods each lasted 4 months from mid-July to early November in 2015 and 2016.
Data analysis
Mean seed densities per m2 were calculated for each seed trap across both study sites and years. Differences between seed densities at the two sites and between the two sampling years were analysed using the Mann–Whitney U-test, because the data were not normally distributed (Zar 2010). Significant differences were accepted at a p value of < 0.05. Furthermore, Quantum GIS (QGIS 2.4.0 Chugiak) was used to create maps of both study sites based on original forest maps and aerial orthophotographs. These maps were used to outline the boundaries of the open areas at each study site, to determine the surrounding 200 m forested search zone, and to mark the positions of seed trees and seed traps (Fig. 1).
Seed dispersal model
A phenomenological model (provided as R-script) developed by van Putten et al. (2012) was used to investigate birch seed dispersal, including directionality (e.g. the effect of wind), the probability of seed deposition at certain distances from the seed source and a dbh-related prediction of seeds per tree and per year. The applied model is capable of accounting for the direction of seed dispersal by differentiating between isotropic and anisotropic dispersal kernels (see Eqs. 1 and 2). ‘Isotropic’ means that seed densities are equally dispersed in all azimuth directions, whereas ‘anisotropic’ dispersal accounts for a directional effect (e.g. due to wind) on seed density distributions (Wagner et al. 2004; Wälder et al. 2009). Because of the above-mentioned model category, which is not mechanistic, the available meteorological data (Table 1) were only used to critically discuss the model results, for example the congruence of wind and seed dispersal direction as part of the anisotropic model.
We fitted seed shadows using the isotropic model and the anisotropic no-shift elliptic distorted-distance model with the free parameters β, ψ and γ. The most important algorithms were described by van Putten et al. (2012). To model the distance-dependent seed distribution (isotropic model), i.e. the ‘kernel’ of the model in the Cartesian coordinate system (x, y), we used the negative exponential distribution as a density function (d(r(x,y)), Eq. 1):
The dispersal distance within the negative exponential function is described by the parameter λ. The value r(x,y) describes the distance between the position of the seed trees and seed traps using the Cartesian coordinates x and y, where seed density is known. To model the anisotropic dispersal, we combined the used ‘kernel’ function of isotropic model (Eq. 1) with the ‘distorted-distance function’ (Eq. 2). The ‘function’ transforms the isotropic (circular) contour lines into anisotropic (non-circular) contour lines (van Putten et al. 2012).
The parameter β (coherency) determines the flattening of the elliptic contour lines, ψ (rotation) describes the rotation of the elliptic contour lines around a seed tree along the common axis, and γ (drift) moves the centre of elliptic contour lines into a positive direction along the common axis (van Putten et al. 2012). Other models, e.g. lognormal, have been tested without improving the results. Additional information such as tree height, crown radius or shelter by neighbouring trees is not considered in this phenomenological model.
Isotropic and anisotropic seed dispersal was modelled separately for each study site (B and E) and year (2015 and 2016). The two study sites were split into the categories ‘uphill’ (B) and ‘downhill’ (E) on the basis of the relief-related seed dispersal. Inverse modelling was applied to fit the observed seed densities. The seed number modelled for each seed trap was calculated by summing the seed rain at a specific location relative to all seed trees. The mean dispersal distance (MDD) in the negative exponential kernel equals λ. In the case of isotropic modelling, the parameter equals MDDiso. The fecundity of a seed tree φ was calculated using the following equation (Eq. 3):
with α as a fecundity parameter defining the allometric relationship between the dbh (mm) and the seed production of a tree. All parameters in the equations were simultaneously estimated by the provided R-script of the inverse models. General references to isotropic and anisotropic inverse modelling can be found in Okubo and Levin (1989), Ribbens et al. (1994), Clark et al. (1999), Skarpaas et al. (2004), Wagner et al. (2004), Soubeyrand et al. (2007), Wälder et al. (2009) and van Putten et al. (2012).
The parametric bootstrap approach, described by Faraway (2006) and Tekle et al. (2016), was used to compare the isotropic and anisotropic inverse model fits and to make a decision on the respective significance level. The likelihood ratio test (LRT) allows comparing models with different numbers of parameters by means of differences in log-likelihood between them. Bootstrap samples (data sets) were generated under a ‘null model’ (isotropic model) using the estimated parameters. The ‘null’ and ‘alternative models’ (anisotropic model) were then fitted based on these data sets and the LRT statistic was computed. This procedure was repeated 99 times for each study site and year. The differences in log likelihood between the isotropic and anisotropic models were used to derive anempirical distribution of LRT, where the null-hypothesis was true. The p value was estimated by comparing the empirical distribution of LRT to the observed values of LRT output (Faraway 2006; Tekle et al. 2016; Frischbier et al. 2019). Additionally, Akaike’s information criterion (AIC) provides information about differences between the statistical models. Spearman´s rank correlation coefficient including p value was used to test the goodness-of-fit (i.e. relation between the observed and predicted seed densities). All computations were performed using the R software version 3.3.2 (package: boot; R Core Team 2014).
Simulations of seed dispersal scenarios
To apply our findings of seed dispersal in a practical context and to support silvicultural management decisions in the context of reforesting disturbed sites, two alternative scenarios of seed tree distribution were designed for study sites B and E, based on the area-specific seed dispersal model results with MDDs of 100 and 350 m, respectively. A regular distribution of seed trees on a 100 m grid surrounding the open areas (i.e. 30 trees) was compared with an aggregated seed tree distribution of the same tree numbers. The simulation was done for two conceptual forested sites with a size of 42 ha (700 m × 600 m) in which the two differently sized and shaped open areas from study sites B and E were integrated. All birch seed trees were assumed to have a reference dbh of 20 cm and to produce 1.5 million seeds as fitted by inverse modelling in 2016.
For analysing the scenarios, we could only consider the open areas, because the open area at site B represents a hilltop with only uphill seed dispersal (not the forested areas), and the sloped open area at site E forms a relief funnel which only represents downhill seed dispersal.
Results
Seed production
The densities of the deposited seeds in both open areas and surrounding forests were significantly higher in 2016 than in 2015 (Mann–Whitney U-test: p < 0.001). Average seed densities ranged between 93 and 23 n m−2 in 2015 and 445 and 86 n m−2 in 2016 at sites B and E, respectively (Fig. 2). Overall, the recorded birch seed densities in traps were at least four times higher in 2016 than in 2015, with a maximum of 9557 (site B) and 311 n m−2 (site E). In both years, birch seed numbers tended to be higher at site B, but this difference was not significant (Mann–Whitney U-test: p > 0.05).

Betula pendula seed densities [n m−2] collected in 2015 and 2016 at study sites B and E. White circles show outliers and black circles inside the boxes are mean values. Differences between years at each study site are significant (Mann–Whitney U-test: p < 0.05)
The allometric relationships between tree dbh and seed production for isotropic models were very tight. The fecundity levels (i.e. ‘α’ in Eq. 3) for the years 2015 (2.0‒2.2) or 2016 (3.6) were relatively similar, indicating a slightly lower fecundity of seed trees in 2015. The expected seed production rate of the isotropic inverse model for a birch seed tree with a mean dbh of 20 cm was approximately 300,000‒366,000 (2015) and 1,430,000‒1,530,000 seeds per tree (2016) (Table 3). Birch seed trees with a dbh of 13‒42 cm produced 0.14‒1.5 million and 0.62‒6.5 million seeds per tree in 2015 and 2016, respectively (Fig. 3).

Isotropic inverse model predictions of seed production per seed tree depending on dbh (diameter at breast height) in 2015 and 2016
Spatial patterns and seed dispersal distances
The source tree-related pattern of cumulated trapped seed numbers followed a negative exponential function (Fig. 4).The highest seed densities at the study sites were found close to the seed sources, e.g. at the northern edge of the open area at site B or close to a stand of Salix caprea, Sorbus aucuparia and B. pendula within a spruce forest adjacent to the open area at site E (see Fig. 1). Seed densities decreased rapidly with increasing distance from the seed source. At a distance of 100 m from the seed sources, mean seed densities of only 24 and 41 n m−2 were observed at site B (uphill dispersal) and E (downhill dispersal) in 2015, respectively, compared to 114 and 181 n m−2 in 2016. During the same period, seed densities of only 15‒25 n m−2 were recorded downhill at a distance of 300 m from the seed sources at site E. In both years, the number of seeds trapped at the same distance from the seed source was slightly higher at site E compared to site B, which is, however, not visible through the overlapping lines in Fig. 4.

Complete set of measured Betula pendula seed densities [n m−2] at study sites B and E in 2015 and 2016 related to the distance from the seed source [m]. Note the different scales of the y-axes
According to the isotropic model, the estimated uphill and downhill mean dispersal distances at site B and E were 97 (2015) and 86 m (2016) and 367 (2015) and 380 m (2016), respectively (Table 3; see left of Fig. 5).

Inverse model predictions of seed shadows [n m−2] simulated for a theoretical single Betula pendula seed tree with a dbh of 20 cm, given isotropic (left) and anisotropic (right) seed dispersal for study sites B and E in 2015 and 2016
Directionality
The probability distribution of the seed dispersal distance of a single tree in the anisotropic models was quite similar to that of the isotropic models (Table 3), although the predicted seed shadows showed directional variation (Fig. 5). Values of β, γ, ψ, which characterize the spatial distribution of the anisotropic seed shadows, are listed in Table 3. However, the anisotropic models mostly featured higher AIC values than the isotropic models (with the exception of study site B in 2016), which indicates the anisotropic model as over-parameterized. The bootstrap statistic tests the hypothesis that the isotropic inverse model does not differ from the anisotropic model. The results of the bootstrap analysis for each year and study site also indicated that the anisotropic model was over-parameterized (bootstrap: p > 0.22) and that the isotropic model was an appropriate approach for all sites and years (Table 3; Fig. 6).

Correlation plots of observed and predicted birch seed densities [n m−2] for study sites B and E in 2015 and 2016
Simulations of seed dispersal scenarios
Based on the study results, scenarios of regular and aggregated seed tree distributions around the studied open areas were created to show that the distribution, the distance between seed trees and the disturbed sites, and the inclination have varying effects on the deposited seed densities.
For an even, systematic seed tree distribution, the comparison of seed shadows between both sites showed higher deposited seed densities in the vicinity of seed trees at site B (Fig. 7a, c), because the same number of produced seeds are distributed uphill over smaller distances than downhill (see Fig. 5). However, in an undisturbed forest with a systematic seed tree distribution, there would be no areas without seed rain as the seed shadows of single seed trees would overlap at both sites. If disturbances interrupted the systematic seed tree grids—like observed at the studied storm-felled areas—and no seed trees were left in the disturbed area, approximately 20‒25 n m−2 birch seeds would still reach the hilltop (site B) and the valley (site E), independent of the inclination. It seems like there are no differences between sites, but the different sizes of the open areas must be considered. If the open area at site B were to exceed the size of the open area at site E, the seed densities deposited in the open area would be considerably lower, which illustrates the aggregated distribution of the same numbers of seed trees (Fig. 7b, d). At distances of 5‒250 m between the seed trees and the forest edges, the seeds dispersed uphill with the shorter MDD are not able to reach the entire open area at site B, but only the southern part. In contrast, the seed shadow of the seeds dispersed downhill from the aggregated seed trees at site E does not really differ from the spatial pattern of systematically distributed seed trees. At least 20 n m−2 seeds are reaching almost all parts of the storm-felled area at site E with the exception of one small spot in the north.

Isotropic seed shadow scenarios [n m−2] of simulated systematic (left) and aggregated (right) seed tree distributions of Betula pendula around the studied open areas at sites B (top) and E (bottom), illustrated as bold lines. Note the different seed density scales for colour illustration in the figure panels
Discussion
Seed production
There is solid evidence for strict allometric relationships between growth parameters, such as dbh, crown radius or basal area of a tree, and the individual seed crop of a tree (Grisez 1975; Greene et al. 2004; Huth 2009; Da Ponte Canova 2018). The fitted fecundity parameter α of the isotropic models for study site B and E was almost identical within each study year, which indicates a strong correlation between tree dbh and seed production, as previously assumed and confirmed for birch by Sato and Hiura (1998), Wagner et al. (2004) and Huth (2009). Therefore, the seed crop of birch trees of variable size can be easily estimated based on their dbh, if the seed production is not hampered by low viability or unfavourable weather conditions (see Grisez 1975). Birches with limited growing space, small crown projected areas and crown volumes have lower seed crops than large solitary individuals (Sarvas 1948). Nevertheless, Huth (2009) noted a restriction of these relationships. With the onsetting senescence of a tree, its seed crop progressively decreases while its diameter and crown dimension generally continue to increase. The strength of the relationship therefore decreases with progressing tree age.
The predicted seed production of a single birch seed tree in 2016 was four times higher than in 2015. Birch is known for a large interannual variability in seed production (Sarvas 1948; Houle and Payette 1990; Kullman 1993; Huth 2009), which is mainly a response to the climatic conditions of the previous year (Kullman 1993; Holmström et al. 2017). On average, good seed years (i.e. so-called mast years) occur every 3 years (Sarvas 1948), during which the percentage of seed germination is significantly higher than in intervening (non-mast) years (Sarvas 1952; Bjorkbom 1971; Houle and Payette 1990). For a single silver birch, Denisow (2007, cited in Huth 2009) reported a seed production of 40,000‒50,000 seeds in intervening years and 3.7‒4.9 million seeds in a mast year. Compared to our study, significantly higher seed crops of individual trees with up to 7.3‒10.0 million seeds (dbh of 24‒80 cm) were reported by e.g. Arnborg (1948, cited in Perala and Alm 1990), Popadyuk et al. (1995), Wagner et al. (2004) and Huth (2009). In the present study, the slightly lower seed production of 0.6‒6.5 million seeds per single seed tree (dbh of13‒42 cm) in 2016 was either a medium seed production year or a mast year. If it was a mast year, the lower seed production numbers might have resulted from smaller birch tree crowns due to strong spruce competition and a lack of release thinning in the past.
Spatial patterns and seed dispersal distances
As expected, the highest seed densities where observed close to the seed trees. At study site B, where birch seed trees were positioned at the edge of the open area, higher densities were observed up to distances of 40‒50 m. Similar results were reported by Sarvas (1948), Fries (1984), Skoglund and Verwijst (1989) and Cameron (1996). In our study, the trapped seed numbers at site B decreased rapidly at distances exceeding 50 m (see Fig. 4). The seed distribution thus showed a negative exponential seed dispersal kernel, as Bjorkbom (1971), Hughes and Fahey (1988), Greene and Johnson (1996) and Karlsson (2001) previously reported for B. alleghaniensis, B. pendula, B. pubescence and B. papyrifera. In theory, one may expect a log-normal distribution of the distances the tiny and lightweight wind-dispersed seeds can travel (Stoyan and Wagner 2001; Huth 2009), but the majority of records in this study showed no peaks at specific distances from the seed trees (as in Greene and Johnson 1996; Stoyan and Wagner 2001; Huth 2009). A feasible explanation for this observation was given by Marquise (1969), who reported that primarily the heaviest and viable birch seeds were deposited close to the seed trees.
In the present study, seed dispersal distances within the sites were similar in both years regardless of the extent of seed production. Large differences of dispersal distances only occurred between the study sites. The modelled mean isotropic dispersal distances (MDDiso) of birch seeds distributed from the forest edge into the open area at site B were 86 and 97 m, as also detected by Wagner et al. (2004) and Huth (2009) for birch in level closed forest stands. In contrast, the MDDiso of 367 and 380 m modelled for seed dispersal from within the adjacent spruce forest stands into the larger open area E indicated dispersal distances that were four times larger. Compared to the present study, Hughes and Fahey (1988), Daniels (2001) and Karlsson (2001) recorded lower dispersal distances of 30‒125 m for B. alleghaniensis, B. pendula and B. pubescens in open areas.
Our very contradictory results of MDD are only comparable with studies from McEuen and Curran (2004), who found seed dispersal distances of B. papyrifera of 700 m between landscape fragments. The long distances were explained by an enormous seed tree presence (see also Zhao et al. 2016) and their extremely high seed production numbers. Similarly, we also had a huge seed source presence in the form of a mixed willow–rowan–birch stand at a distance of 74 m from the open area at site E (see Table 2; Fig. 1). This potential explanation is further be confirmed by Greene and Johnson (1996), Zhao et al. (2016) and Holmström et al. (2017) who mentioned that seeds are dispersed over larger distances if the size of the canopy opening increases, if seed source densities are located close to forest edges or if seed tree density is high. Therefore, to increase the probability of getting birch seeds onto storm-felled sites, an increasing number of seed sources would be needed at longer distances to the respective sites. Nevertheless, the few birch seed trees around the open area at site B were standing at the forest edge (see Fig. 1), making it actually more likely that seeds would disperse over larger distances than in closed forests (Holmström et al. 2017).
Thus, another effect should be considered: the relief, although this effect has not often been mentioned in previous studies. In this study, it seems that the dispersal distances of birch seeds decreased uphill (site B) or increased downhill (site E) depending on the inclination and seed tree position (valley, slope or plateau) relative to the storm-felled site. Hill and Stevens (1981), who studied soil seed banks, found a 30 m shorter distance for uphill deposition of birch seeds than for downhill deposition. Based on the estimated MDDiso, we may therefore assume a strong effect of inclination on the dispersal distance of birch seeds. However, the two study sites were not selected rigorously enough to test for inclination effects. Therefore, the results should be conservatively interpreted. Some of the other aforementioned factors, e.g. differently sized open areas, seed tree densities and distances between seed sources and forest edges, may also have influenced the dispersal distances. Consequently, the documented results of this case study are not as such directly transferable to all forest areas.
The distance of long dispersal is particularly determined by secondary dispersal (Matlack 1989; Greene and Johnsons 1997), but this factor can be neglected in the present study due to similar ground vegetation cover and thus likely similar secondary dispersal in both open areas. Based on our data, we have no chance to check for secondary seed dispersal in any way.
Directionality
Due to non-random anemochorous seed dispersal, previous studies often showed directionality for birch seed dispersal (Wagner et al. 2004; Wright et al. 2008; Huth 2009). However, this was not clearly confirmed by results of the present study. An explanation for the surprising isotropy of seed dispersal in this study might be the relatively long seed collection periods (4 month). The variability of wind directions and wind speeds occurring during a period of 4 months—boosted and modified by turbulence and varying wind speeds in the open areas due to vegetation cover and structure (see Moon et al. 2013)—may explain the lack of observed anisotropy. While the half-hourly meteorological data (Table 1) showed no extreme wind events during the two study periods, with a maximum wind speed of 9.9 m s−1, the monthly mean wind direction featured high standard deviations of ± 25 to ± 122°. On some days during the study periods, the variability of the wind direction was as high as ± 206°. The observed isotropic distributions are thus plausible, because no prevailing wind direction was identified for either study period and birch seeds were dispersed in a variety of directions. The assumption that the anisotropic models reveal a better fit to the empirical 4-month data was therefore rejected. Houle and Payette (1990) found anisotropic spatial patterns of the seed shadows of B. alleghaniensis after subdividing the seed rain period into shorter study periods. If we had chosen shorter periods for emptying the seed traps in this study (e.g. 14 day periods), we might have also been able to detect anisotropy by inverse modelling (Wagner et al. 2004).
However, short-term analyses of 14 day periods would not be useful for deriving silvicultural recommendations, because in this context the entire period of seed rain, i.e. 3‒4 months in summer and autumn, has to be considered. For silvicultural practice, it is important to know that equally distributed seed rain can be expected around seed trees if strong wind regimes with variable wind directions prevail at a specific site.
Simulations of seed dispersal scenarios
The abundance and the spatial pattern of potential seed trees near a particular disturbed site are important determinants upon which forest managers could base silvicultural decisions about risk-adapted reforestation concepts and preventative risk-adapted seed tree management.
The seed dispersal scenarios for silvicultural management decisions showed that in case of a systematic, even seed tree distribution, inclination differences are not clearly affecting the seed shadow, and known limits of birch dispersal distances (MDD) can generally be compensated, especially for uphill seed dispersal. However, if seed trees are aggregated and unevenly distributed, the effect of the inclination is clearly visible in the resulting seed shadow. In the latter case, the inclination has an important influence on the regeneration success of storm-felled sites in addition to the distance between the seed sources and the open area as well as the number of seed trees or tree groups.
Recommendations of silvicultural management decisions
For the practical management of regenerating disturbed sites, it can be concluded that seed dispersal is very sensitive to the distribution, the number and loss of seed trees in nearby forests as well as the size and inclination of disturbed areas. Hence, Willis et al. (2016) highlighted the importance of local seed source availability for successful birch regeneration. Sarvas (1948) mentioned that at least 100 to 200 n m−2 of viable seeds are necessary for successful regeneration and recommends a density of 4 to 8 n ha−1 of B. pendula seed trees, whereas Safford and Jacobs (1983) advocate a seed tree number of 7 to 12 n ha−1 for B. papyrifera. Based on Sarva’s (1948) minimum recommendation of 100 seeds m−2, we can also give recommendations on the required seed tree numbers in forests. Derived from the uphill and downhill mean dispersal distances and the deposited seed numbers at study sites B and E (cf. Figure 4 and 7), 4 and 16 n ha−1 seed trees are needed under unfavourable conditions, e.g. in non-mast years. This corresponds to seed tree grid intervals of 60 and 30 m, respectively. For small groups of trees, the distance can be slightly larger, as illustrated in Fig. 7. Higher source tree numbers are needed for ensuring the natural regeneration of disturbed sites if the present seed trees produce insufficient seed crops as a result of lacking tending measures. Thus, the aforementioned seed tree numbers should be considered as minimum numbers.
However, with respect to uphill dispersal, we assume that areas with insufficient numbers of deposited seeds will probably always occur in disturbed sites, unless the area is completely surrounded by seed trees. In case of disturbed sites of more than 4 ha, it is thus impossible to ensure a full cover of natural birch regeneration due to the limited seed dispersal distances. The natural regeneration of such large areas should therefore be supported by additional reforestation measures. In addition, seed trees surviving after a disturbance event must not be removed from the disturbed sites and seed trees in the vicinity of the sites should be promoted and vitalized wherever possible. Advance regeneration, which established underneath the canopy prior to the disturbance event, can also provide valuable benefits for regenerating disturbed sites. For the regeneration of small disturbed areas, which usually occur more frequently (Brang et al. 2015), risk-adapted forest management should include the ‘spatial optimization’ of birch seed trees within conifer forests, due to the limited dispersal distance of birch seeds in general. A few groups of aggregated seed trees within a forest stand or some seed trees along the forest edge and forest roads in otherwise pure conifer forests are not sufficient for regenerating disturbed areas, but can be a good initial for the integration of birch trees in conifer forests. Along forest roads, paths or trails, birch trees have the possibility of unrestricted crown growth on one side and thus feature a more proliferous seed production. However, a network of more or less regularly distributed individual birches is needed within conifer forests, preferably even of small groups of seed trees, because silvicultural measures are easier to implement (see Cameron 1996; Hynynen et al. 2010) and the chances of successfully regenerating more distant sites is higher. For birches, Cameron (1996) mentioned required thinning intervals of 5‒7 years for ensuring a good crown growth. Conservation, vitalization and propagation are important factors for annual birch seed crop quality and quantity, which makes more sense for tree groups than for single trees due to costs and the regulation of interspecific competition.
Conclusions
To ensure successful regeneration, reliable empirical information concerning seed production and seed dispersal distances are necessary. Inverse modelling was applied to fit seed distribution and seed production on the basis of observed seed densities.
The findings of our case study indicated a strong influence of (1) site inclination, (2) seed tree position (valley, slope or plateau) and distance to a storm-felled site on the seed dispersal of B. pendula. The seed shadow of birch trees is also influenced by (3) the number of seed sources. In the context of natural B. pendula regeneration on disturbed sites, it must be noted that seed dispersal is spatially limited compared to other anemochorous tree species such as Salix ssp. (Gage and Cooper 2005; Tiebel et al. 2019). However, in light of the specific characteristics of the selected study areas, the transfer of the presented results to other sites and regions should be cautious.
For the practical management of regenerating disturbed sites, it can be concluded that seed dispersal is very sensitive to the distribution, the number and loss of seed trees in nearby forests as well as the size and inclination of disturbed areas. Therefore, risk-adapted forest management should include the ‘spatial optimization’ of birch seed trees, ideally creating a network of small seed tree groups scattered more or less regularly within pure conifer forests.
Change history
13 August 2021
A Correction to this paper has been published: https://doi.org/10.1007/s10342-021-01404-w
References
Atkinson MD (1992) Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. J Ecol 80:837–870. https://doi.org/10.2307/2260870
Augspurger CK, Franson SE (1987) Wind dispersal of artificial fruits varying in mass, area, and morphology. Ecology 68:27–42. https://doi.org/10.2307/1938802
Bakker JP, Poschlod P, Strykstra RJ, Bekker RM, Thompson K (1996) Seed banks and seed dispersal: important topics in restoration ecology. Acta Bot Neerl 45:461–490. https://doi.org/10.1111/j.1438-8677.1996.tb00806.x
Beck P, Caudullo G, de Rigo D, Tinner W (2016) Betula pendula, Betula pubescens and other birches in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (eds) European atlas of forest tree species. Publications Office of the European Union, Luxembourg, pp 70–73. https://doi.org/10.2788/038466
Bjorkbom JC (1971) Production and germination of paper birch seed and its dispersal into a forest opening. Northeastern Forest Experiment Station, Forest Service Research Paper NE-209, U.S. Department of Agriculture, Upper Darby, USA
Brang P, Hilfiker S, Wasem U, Schwyzer A, Wohlgemuth T (2015) Langzeitforschung auf Sturmflächen zeigt Potenzial und Grenzen der Naturverjüngung. Schweiz Z Forstwes 166:147–158
Brouwer W, Stählin A (1975) Handbuch der Samenkunde für Landwirtschaft, Gartenbau und Forstwirtschaft. DLG-Verlags-GmbH, Frankfurt (Main)
Bürger M (2003) Bodennahe Windverhältnisse und windrelevante Reliefstrukturen. In: Leibnitz-Institut für Länderkunde (ed) Nationalatlas Bundesrepublik Deutschland. Spektrum Akademischer Verlag, pp 52–55
Burse K-D, Schramm H-J, Geiling S, Meinhardt H, Schölch M (1997) Die forstlichen Wuchsbezirke Thüringens-Kurzbeschreibung. Mitteilungen der Landesanstalt für Wald und Forstwirtschaft, Gotha
Bushart M, Suck R (2008) Potenzielle natürliche Vegetation Thüringens, Schriftenr. der Thüringer Landesanstalt für Umwelt und Geologie, Jena
Cameron AD (1996) Managing birch woodlands for the production of quality timber. Forestry 69:357–371. https://doi.org/10.1093/forestry/69.4.357
Clark JS, Macklin E, Wood L (1998) Stages and spatial scales of recruitment limitation in southern Appalachian forests. Ecol Monogr 68:213–235. https://doi.org/10.1890/0012-9615(1998)068%5b0213:SASSOR%5d2.0.CO;2
Clark JS, Silman M, Kern R, Macklin E, HilleRisLambers J (1999) Seed dispersal near and far: patterns across temperate and tropical forests. Ecol Monogr 80:1475–1494. https://doi.org/10.1890/0012-9658(1999)080%5b1475:SDNAFP%5d2.0.CO;2
Da Ponte Canova G (2018) Regeneration ecology of anemochorous tree species Qualea grandiflora Mart. and Aspidosperma tomentosum Mart. of the cerrado Aguara Ñu located in the Mbaracayú Nature Forest Reserve (MNFR), Paraguay. Dissertation, Technical University of Dresden
Daniels J (2001) Ausbreitung der Moorbirke (Betula pubescens Ehrh. agg.) in gestörten Hochmooren der Diepholzer Moorniederung. Osnabrücker Naturwissenschaftliche Mitteilungen 27:39–49
Faraway JJ (2006) Extending the linear model with R—generalized linear mixed effects and nonparametric regression models. CRC, Boca Raton
Fenner M (1985) Seed ecology. Chapman and Hall Ltd, London
Ferris R, Humphrey JW (1999) A review of potential biodiversity indicators for application in British forests. Forestry 72:313–328. https://doi.org/10.1093/forestry/72.4.313
Fink AH, Brücher T, Ermert V, Krüger A, Pinto JG (2009) The European storm Kyrill in January 2007: synoptic evolution, meteorological impacts and some considerations with respect to climate change. Nat Hazards Earth Syst Sci 9:405–423. https://doi.org/10.5194/nhess-9-405-2009
Fries G (1984) Den frösådda björkens invandring på hygget (Immigration of birch into clearfelled areas). Sver. Skogsvardsfdrb. Tidskr. 82:35–39
Frischbier N, Profft I, Hagemann U (2014) Potential impacts of climate change on forest habitats in the Biosphere Reserve Vessertal-Thuringian Forest in Germany. In: Rannow S, Neubert M (eds) Managing protected areas in Central and Eastern Europe under climate change, advances in global change research. Springer, Dordrecht, pp 243–257. https://doi.org/10.1007/978-94-007-7960-0_16
Frischbier N, Tiebel K, Tischer A, Wagner S (2019) Small scale rainfall partitioning in a European beech forest ecosystem reveals heterogeneity of leaf area index and its connectivity to hydro- and atmosphere. Geosciences 9:1–23. https://doi.org/10.3390/geosciences9090393
Gage EA, Cooper DJ (2005) Patterns of willow seed dispersal, seed entrapment, and seedling establishment in a heavily browsed montane riparian ecosystem. Can J Bot 83:678–687. https://doi.org/10.1139/B05-042
Gauer J, Aldinger E (2005) Waldökologische Naturräume Deutschlands: Forstliche Wuchsgebiete und Wuchsbezirke-mit Karte 1:1.000.000, Mitteilungen des Vereins für Forstliche Standortskunde und Forstpflanzenzüchtung, Stuttgart
Graber RE, Leak WB (1992) Seed fall in an old-growth northern hardwood forest. Northeastern Forest Experiment Station, Forest Service Research Paper NE-663, Pennsylvania, USA
Greene DF, Johnson EA (1996) Wind dispersal of seeds from a forest into a clearing. Ecology 77:595–609. https://doi.org/10.2307/2265633
Greene DF, Johnson EA (1997) Secondary dispersal of tree seeds on snow. J Ecol 85(3):329–340. https://doi.org/10.2307/2960505
Greene DF, Canham CD, Coates KD, Lepage PT (2004) An evaluation of alternative dispersal functions for trees. J Ecol 92:758–766. https://doi.org/10.1111/j.0022-0477.2004.00921.x
Gregow H, Laaksonen A, Alper ME (2017) Increasing large scale windstorm damage in Western, Central and Northern European forests, 1951–2010. Sci Rep 7:1–7. https://doi.org/10.1038/srep46397
Grisez T (1975) Flowering and seed production in seven hardwood species. Northeastern Forest Experiment Station, Forest Service Research Paper NE-315, Upper Darby, USA
Hill MO, Stevens PA (1981) The density of viable seed in soils of forest plantations in upland Britain. J Ecol 69:693–709. https://doi.org/10.2307/2259692
Holmström E, Karlsson M, Nilsson U (2017) Modelling birch seed supply and seedling establishment during forest regeneration. Ecol Model 352:31–39. https://doi.org/10.1016/j.ecolmodel.2017.02.027
Houle G (1998) Seed dispersal and seedling recruitment of Betula alleghaniensis: spatial inconsistency in time. Ecology 79:807–818
Houle G, Payette S (1990) Seed dynamics of Betula alleghaniensis in a deciduous forest of north- eastern North America. J Ecol 78:677–690. https://doi.org/10.2307/2260892
Hughes JW, Fahey TJ (1988) Seed dispersal and colonization in a disturbed northern hardwood forest. Bull Torrey Bot Club 115:89–99
Humphrey JW, Holl K, Broome AC (1998) Birch in spruce plantations—management for biodiversity. Forestry Commission Technical Paper 26
Huth F (2009) Untersuchungen zur Verjüngungsökologie der Sand-Birke (Betula pendula Roth). Dissertation, Technical University of Dresden
Hynynen J, Niemistö P, Viherä-Aarnio A, Brunner A, Hein S, Velling P (2010) Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83:103–119. https://doi.org/10.1093/forestry/cpp035
Kadereit JW, Leins P (1988) A wind tunnel experiment on seed dispersal in Papaver L. sects. Argemonidium SPACH and Rhoeadium SPACH (Papaveraceae). Flora 181:189–203. https://doi.org/10.1016/S0367-2530(17)30365-1
Karlsson M (2001) Natural regeneration of broadleaved tree species in southern Sweden: effects of silvicultural treatments and seed dispersal from surrounding stands. Dissertation, Swedish University of Agricultural science
Koski V, Rousi M (2005) A review of the promises and constraints of breeding silver birch (Betula pendula Roth) in Finland. Forestry 78:187–198. https://doi.org/10.1093/forestry/cpi017
Kullman L (1993) Tree limit dynamics of Betula pubescens ssp. tortuosa in relation to climate variability: evidence from central Sweden. J Veg Sci 4:765–772. https://doi.org/10.2307/3235613
Leder B (1992) Weichlaubhölzer: Verjüngungsökologie, Jugendwachstum und Bedeutung in Jungbeständen der Hauptbaumarten Buchen und Eiche, Schriftenreihe der Landesanstalt für Forstwissenschaft Nordrhein-Westfalen
Luostarinen K, Verkasalo E (2000) Birch as sawn timber and in mechanical further processing in Finland. A literature study. Finnish Society of Forest Science–Finnish Forest Rearch Institut, Helsinki
Marquise DA (1969) Silvical requirements for natural birch regeneration. Northeastern Forest Experiment Station, Forest Service, Upper Darby, USA
Matlack GR (1989) Dispersal of seed across snow in Betula lenta, a gap-colonizing tree species. J Ecol 77:853–869. https://doi.org/10.2307/2260990
McEuen AB, Curran LM (2004) Seed dispersal and recruitment limitation across spatial scales in temperate forest fragments. Ecology 85:507–518. https://doi.org/10.1890/03-4006
Moles AL, Falster DS, Leishman MR, Westoby M (2004) Small-seeded species produce more seeds per square metre of canopy per year, but not per individual per lifetime. J Ecol 92:384–396. https://doi.org/10.1111/j.0022-0477.2004.00880.x
Mölter T, Schindler D, Albrecht AT, Kohnle U (2016) Review on the projections of future storminess over the North Atlantic European region. Atmosphere 7:1–40. https://doi.org/10.3390/atmos7040060
Moon K, Duff TJ, Tolhurst KG (2013) Characterising forest wind profiles for utilisation in fire spread models. In: Piantadosi J, Anderssen RS, Boland J (eds) MODSIM2013, Modelling and simulation society of Australia and New Zealand. Presented at the 20th International Congress on Modelling and Simulation, Adelaide, Australia, pp 214–220
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Tree 15:278–285. https://doi.org/10.1016/S0169-5347(00)01874-7
Okubo A, Levin SA (1989) A theoretical framework for data analysis of wind dispersal of seeds and pollen. Ecology 70:329–338. https://doi.org/10.2307/1937537
Patterson GS (1993) The value of birch in upland forests for wildlife conservation. HMSO Publications Centre, London
Perala DA, Alm AA (1990) Reproductive ecology of birch: a review. For Ecol Manag 32:1–38. https://doi.org/10.1016/0378-1127(90)90104-J
Popadyuk RV, Smirnova OV, Evstigneev OI, Yanitskaya TO, Chumatchenko SI, Zaugolnova LB, Korotkov VN, Chistyakova AA, Khanina LG, Komarov AS (1995) Current state of broad-leaved forests in Russia, Belorussia, Ukraine: historical development, biodiversity, structure and dynamic. PRC RAS, Pushchino
Priha O (1999) Microbial activities in soils under Scots pine, Norway spruce and silver birch. Dissertation, University of Helsinki
Ribbens E, Silander JA Jr, Pacala SW (1994) Seedling recruitment in forest: calibrating models to predict patterns of tree seedling dispersion. Ecology 75:1794–1806. https://doi.org/10.2307/1939638
Röhrig E, Gussone HA (1990) Waldbau auf ökologischer Grundlage. Band 2: Baumartenwahl, Bestandesbegründung und Bestandespflege. Verlag Paul Parey, Hamburg
Roloff A, Pietzarka U (2010) Betula pendula ROTH. In: Roloff A, Weisberger H, Lang U, Stimm B (eds) Bäume Mitteleuropas. Wiley, New York, pp 61–75
R Core Team (2014) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org
Safford LO, Jacobs RD (1983) Paper birch. In: Burns RM (ed) Silvicultural systems for the major forest types of the United States, U.S. Department of Agriculture Washington, D. C., Agriculture Handbook No. 445, pp 145–147
Sarvas R (1948) A research on the regeneration of birch in South Finland. Commun. Inst. For. Fenn. 35:82–91
Sarvas R (1952) On the flowering of birch and the quality of seed crop. Commun Inst For Fenn 40:1–35
Sato H, Hiura T (1998) Estimation of overlapping seed shadows in a northern mixed forest. For Ecol Manag 104:69–76. https://doi.org/10.1016/S0378-1127(97)00247-8
Skarpaas O, Stabbetorp OE, Rønning I, Svennungsen TO (2004) How far can a hawk’s beard fly? Measuring and modelling the dispersal of Crepis praemorsa. J Ecol 92:747–757. https://doi.org/10.1111/j.0022-0477.2004.00915.x
Skarpaas O, Auhl R, Shea K (2006) Environmental variability and the initiation of dispersal: turbulence strongly increases seed release. Proc. Biol. Sci. 273:751-756. https://doi.org/10.1098/rspb.2005.3366
Skoglund J, Verwijst T (1989) Age structure of woody species populations in relation to seed rain, germination and establishment along the river Dalälven, Swedenn. Vegetation 82(25):34. https://doi.org/10.1007/BF00217979
Soubeyrand S, Enjalbert J, Sanchez A, Sache I (2007) Anisotropy, in density and in distance, of the dispersal of yellow rust of wheat: experiments in large field plots and estimation. Phytopathology 97:1315–1324. https://doi.org/10.1094/PHYTO-97-10-1315
Stoyan D, Wagner S (2001) Estimating the fruit dispersion of anemochorous forest trees. Ecol Model 145:35–47. https://doi.org/10.1016/S0304-3800(01)00385-4
Tekle FB, Gudicha DW, Vermunt JK (2016) Power analysis for the bootstrap likelihood ratio test for the number of classes in latent class models. Adv Data Anal Classif 10:209–224. https://doi.org/10.1007/s11634-016-0251-0
Tiebel K, Leinemann L, Hosius B, Schlicht R, Frischbier N, Wagner S (2019) Seed dispersal capacity of Salix caprea L. assessed by seed trapping and parentage analysis. Eur J For Res 138:495–511. https://doi.org/10.1007/s10342-019-01186-2
van Dorp D, van den Hoek WPM, Daleboudt C (1996) Seed dispersal capacity of six perennial grassland species measured in a wind tunnel at varying wind speed and height. Can J Bot 74:1956–1963. https://doi.org/10.1139/b96-234
van Putten B, Visser MD, Muller-Landau HC, Jansen PA (2012) Distorted-distance models for directional dispersal: a general framework with application to a wind-dispersed tree. Methods Ecol Evol 3:642–652. https://doi.org/10.1111/j.2041-210X.2012.00208.x
Waesch G (2003) Montane Graslandvegetationen des Thüringer Waldes: Aktueller Zustand, historische Analyse und Entwicklungsmöglichkeiten. Cuvillier Verlag, Göttingen
Wagner S, Wälder K, Ribbens E, Zeibig A (2004) Directionality in fruit dispersal models for anemochorous forest trees. Ecol Model 179:487–498. https://doi.org/10.1016/j.ecolmodel.2004.02.020
Wälder K, Näther W, Wagner S (2009) Improving inverse model fitting in trees-Anisotropy, multiplicative effects, and Bayes estimation. Ecol Model 220:1044–1053. https://doi.org/10.1016/j.ecolmodel.2009.01.034
Willis JL, Walters MB, Farinosi E (2016) Local seed source availability limits young seedling populations for some species more than other factors in northern hardwood forests. For Sci 62:440–448. https://doi.org/10.5849/forsci.15-143
Wright SJ, Trakhtenbrot A, Bohrer G, Detto M, Katul GG, Horvitz N, Muller-Landau HC, Jones FA, Nathan R (2008) Understanding strategies for seed dispersal by wind under contrasting atmospheric conditions. PNAS 105:19084–19089.
Zar JH (2010) Biostatistical analysis. Prentice Hall, Upper Saddle River
Zerbe S (2001) On the ecology of Sorbus aucuparia (Rosaceae) with special regard to germination, establishment and growth. Pol Bot J 46:229–239. https://doi.org/10.1234/12345678
Zerbe S (2009) Renaturierung von Waldökosystemen. In: Zerbe S, Wiegleb G (eds) Renaturierung von Ökosystemen in Mitteleuropa. Spektrum Akademischer Verlag, Heidelberg, pp 153–182
Zhao F, Qi L, Fang L, Yang J (2016) Influencing factors of seed long-distance dispersal on a fragmented forest landscape on Changbai Mountains, China. Chin Geogr Sci 26:68–77. https://doi.org/10.1007/s11769-015-0747-0
Acknowledgements
This work was financially supported by a scholarship granted to K. Tiebel by the foundation ‘Deutsche Bundesstiftung Umwelt’ (DBU), the ‘Graduiertenakademie’ (GA) of TU Dresden and ThüringenForst, Forestry Research and Competence Center, Gotha, Germany. We would like to thank Sonja Gockel (Thuringian forest conversion project) and colleagues from ThüringenForst for providing the study sites; and Antje Karge, Angelika Otto, Alexandra Wehnert and Jörg Wollmerstädt from TU Dresden for field assistance. We thank Ulrike Hagemann for proofreading the text.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Christian Ammer.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised due to a retrospective Open Access order.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tiebel, K., Huth, F., Frischbier, N. et al. Restrictions on natural regeneration of storm-felled spruce sites by silver birch (Betula pendula Roth) through limitations in fructification and seed dispersal. Eur J Forest Res 139, 731–745 (2020). https://doi.org/10.1007/s10342-020-01281-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-020-01281-9