Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario


1
Department of Biological Sciences, Brock University, 1812 Sir Isaac Brock Way, St. Catharines, ON L2S 3A1, Canada
2
Department of Biology, Wilfrid Laurier University, 75 University Avenue West, Waterloo, ON N2L 3C5, Canada
*
Author to whom correspondence should be addressed.
Agronomy 2020, 10(4), 529; https://doi.org/10.3390/agronomy10040529
Submission received: 19 March 2020
/
Revised: 1 April 2020
/
Accepted: 2 April 2020
/
Published: 8 April 2020
(This article belongs to the Section Weed Science and Weed Management)
Abstract
:Amaranthus hybridus is a noxious weed in Ontario, with demonstrated allelopathic properties that can lead to decreased agricultural production. We tested the germination and growth of five cover crop species exposed to A. hybridus extracts, and to dried or fresh materials in soil. A germination index was calculated, and the dry weight of plant organs were measured to quantify responses to treatments. All species had reduced germination (≤29%) in 100% extract. Trifolium pratense had significant root weight reductions in extract (52%) and dried (72%) treatments, whereas shoot weight only decreased (48%) in dried treatment. Medicago sativa shoot weight decreased (52%) in 20g fresh treatment, while root weight decreased (62%) in dried treatment. Shoot weight of Raphanus sativus increased (32%) at mid-extract concentrations, while root weight increased (33%) only with dried treatment; however, both its shoot and root weight decreased (>40%) in fresh treatment. Only the shoot weight of Lolium multiflorum increased (41% in 75% extract and 55% in dried treatment). Both Cichorium intybus shoot and root weights decreased (~50%) in fresh treatment. Crop responses to A. hybridus are complex, and material and species-dependant. Further testing in the field may provide a more comprehensive understanding of how to improve the management of A. hybridus.
Keywords:
allelopathy; cover crops; germination; plant growth; competition; facilitation; agriculture1. Introduction
Crops and weeds interact with one another in a variety of ways and can be classified as either competitive (e.g., for water, nutrients or light), facilitative (e.g., enhancement of soil nutrient availability resulting from nitrogen symbiotic fixation capacity of legumes) or neutral (i.e., no obvious interactions) [1,2]. Agricultural systems can include not only desired cash crops, but also species, often called cover crops, planted for other purposes than direct monetary gain, such as nutrient transfer (e.g., N and P in legume and potato intercrops; N in soybean and maize intercrops) or beneficial insect attraction (e.g., higher incidence of predacious ground beetles in buckwheat and squash intercrops) [3,4,5]. Cover crop species are often grown for their ability to inhibit weed establishment without any adverse effects on the main crop [6,7]. Farmers may choose to introduce cover crops alongside, or in rotation with, the main crop to reduce their use of herbicides [8]. However, the effectiveness of these cover crops will depend on their capacity to overcome weed competition. Cover crops are often used in perennial crop systems such as vineyards. They have been used for various reasons, such as protecting the soil against erosion, improving soil fertility and water holding capacity, and reducing weed pressure [8,9].
One phenomenon that is often associated with weed presence and is the subject of much debate in the literature is allelopathy [10]. Allelopathy can be defined as the direct or indirect effects (positive or negative) caused by the transfer of chemicals from one plant to any other plant species [11]. The magnitude of the impact of allelopathic effects depends on the type of allelochemicals and the target plant. While the aqueous leaf extract of Crotalaria juncea inhibits the germination of bell peppers and tomatoes, germination of other crops varies from 95% inhibition for onions to only 2% inhibition for cucumbers and winter wheat [12]. Johnson grass (Sorghum halepense L.) roots inhibit both the germination and growth of soybean (Glycine max L.), pea (Pisum sativum L.), and some vetch species (Securigera sp. DC.) [13]. Extracts of five different species of Amaranthus can also inhibit up to 95% of lettuce (Lactuca sativa L.) germination [14]. The potential for plant growth inhibition through allelopathy has been used to control weeds. In rice fields, sorghum can reduce weed biomass by 40% [15]. Furthermore, a combined plant extract made from sorghum, sunflower (Helianthus sp. L.), brassica (Brassica sp. L.), and mulberry (Morus alba L.) can inhibit the growth of horse purslane (Trianthema portulacastrum L.) [16]. Allelopathic interactions can in other cases be facilitative, as is the case for relay intercrops of eggplant (Solanum melongena L.) and garlic (Allium sativum L.), leading to an increase in eggplant yield [17]. Incorporating blackboard tree (Alstonia scholaris L.) residue into the soil used to grow sword lily (Gladiolus grandiflorus Andrews) reduces corm-rot disease incidence by 80%, thereby increasing final plant dry weight [18].
Despite a rich body of evidence that indicates the existence of allelopathy, it is often argued that allelopathy does not directly influence plant interactions but may be a facilitator of some other mechanism that is ultimately responsible for the plant responses to one another [19]. Yanyan et al. (2018) find that alligator weed (Alternanthera philoxeroides Mart.) root extracts affect both soil enzymes and microbial community structure, suggesting that allelopathic plants can indirectly influence other plants by changing microbial action in the soil [20]. Nonetheless, whether or not allelopathy is responsible for directly or indirectly influencing plant interactions, further research is required to understand if the potential for such interactions exists, so that current cropping management strategies, especially in organic farming systems, can be improved.
Smooth pigweed (Amaranthus hybridus L.) is recognized by the Ontario Ministry of Agriculture, Food, and Rural Affairs (OMAFRA) as being one of three pigweed species capable of having a negative impact on crops in Southern Ontario, Canada [21]. Its rapid growth during the summer and its ability to outcompete crop species for water, light, and nutrients lead to the reduction of crop yields [20]. In addition, A. hybridus has been shown to be an acceptable alternate host to many common agriculture pests, such as flea beetle (tribe Alticini Spinola), tarnished plant bug (Lygus lineolaris P. Beauv.), and green peach aphid (Myzus persicae Sulzer) [21,22]. While A. hybridus is known to be detrimental to crops due to its physical presence in a cultivated field, comparatively less is known regarding its impact on cover crops, which are used in some vineyards in Ontario, Canada. Recent studies have demonstrated that exudates of a closely-related species, red-root pigweed (Amaranthus retroflexus L.), have negative impacts through allelopathic effects on the germination and growth of several crops [23,24,25,26]. It is essential to determine what A. hybridus presence means for local cover crop management strategies, so that we can attempt to mitigate economic loss. Of additional concern, Trucco et al. (2006) report that localized populations of A. hybridus in Illinois, USA, have developed some resistance to commercially available herbicides [27].
The objective of this study was to determine if locally-sourced A. hybridus extracts had the potential to influence the germination and growth characteristics of five commonly-used cover crop species (two forbs, two legumes, and one grass species, respectively): common chicory (Cichorium intybus L.), oilseed radish (Raphanus sativus L. variety Nitro), red clover (Trifolium pratense L.), alfalfa (Medicago sativa L.), and annual ryegrass (Lolium multiflorum Lam.) These species were selected as they are commonly used in vineyards of the Niagara region, Canada. In a recent field-study, Arroyo et al. (2018) argue that lab studies using extracts tend to overestimate allelopathic effects and call for more realistic exposure conditions when conducting tests [28]. To respond to this criticism, in this study, the cover crops were also exposed to dry and fresh field plant material, to better mimic what can happen in the field with plant residues left in the soil after tillage. Since plant organ exudates from A. hybridus and other closely-related species have a demonstrated ability to inhibit the germination and growth of some cash crop species, we hypothesized that germination and growth of the cover crop species used would be adversely affected. Furthermore, since exposing plants to concentrated extracts from potentially allelopathic species is not typically how plants encounter allelopathic chemicals in the environment, we predicted that plant response to an allelopathic individual would change based on the mode of exposure (extracts versus dried ground material versus fresh field plant material).
2. Materials and Methods
A. hybridus entire specimens were collected from a vineyard located in the Niagara region, Ontario, Canada, in September 2016 and September 2017. Plants collected in September 2016 were used for the germination and two of three growth experiments, i.e., treatments consisting of tea solution or dried ground material. Plants were dried in a plant dryer (30 ˚C) for a period of 14 days, and then dissected into roots, leaves, and stems (always maintained in the dark (no light exposure) during the experiments). Once dried, each organ was individually ground using a coffee bean mill and mixed to get a 25 g mixture of dried material (5 g roots, 5 g stems, 15 g leaves). This mixture was then added to 1 L of sterilized distilled water at room temperature and allowed to steep for 24 hours (entire procedure taken from Shahrokhi et al. 2011) [25]. This solution, which we referred to as tea solution, was used immediately after the steeping period. New batches were prepared as required. Fresh A. hybridus plants were also collected in September 2017 to be used in the third growth experiment, which consisted of fresh trimmed material of entire A. hybridus plants. The processed fresh plant material used in this third experiment was kept refrigerated for four weeks until the experiment started. All fresh materials were kept in the dark as well.
The tea was used as a treatment to measure the A. hybridus allelopathic effects on both cover crop seed germination and plant growth. The incorporation of dried ground plant material to the soil was used as a complementary treatment to assess growth, as it was more reflective of how plants encounter allelopathy in nature. Testing of crop growth using fresh A. hybridus material (trimmed in pieces of 1cm2 to reflect what would be left behind after mechanical ploughing and incorporated throughout each entire pot of soil to reflect the depth at which mechanical ploughing would have distributed/integrated the material in the field) was included as a final experiment. This treatment was added as it would most closely resemble how A. hybridus would be distributed in the environment after being mechanically turned over in the soil during the plowing process. All experiments targeted the same species (common chicory, oilseed radish, red clover, alfalfa, and annual ryegrass). Annual ryegrass seeds were obtained from the vineyard (Beamsville, ON). Chicory, oilseed radish, red clover and alfalfa were obtained from Canadian Comfort Alpacas (Fenwick, ON). Four experiments in total were performed, and we referred to them below as (1) germination experiment, (2) growth experiment with tea solution, (3) growth experiment with dried ground material, and (4) growth experiment with fresh trimmed material.
2.1. Germination Experiment
To determine how A. hybridus affected seed germination, tea extracts were used; the tea was made using distilled water to obtain the following tea dilutions: 10%, 25%, 50%, and 75%. Undiluted tea (100% pure solution) and a control of pure distilled water (0% of A. hybridus solution) were used as two additional treatments. Ten seeds from each cover crop species were placed in separate Petri dishes (10 cm, plastic Fisherbrand™), lined with a single sheet of Whatman No. 4 sterile filter paper. Ten replicates were performed for each crop species-treatment category combination (10 seeds times 6 treatment dilutions times 10 replicates equal 600 seeds per species). Each Petri dish was then filled with 5 ml of its corresponding treatment solution, sealed with Saran™ wrap, and placed randomly in a growth cabinet (set for complete darkness, 24 ˚C, with a relative humidity of 65%). Seeds were checked every 24 hours over the course of seven days for signs of germination. Germination was considered successful when the radicle was equal to, or longer than, the seed at its longest point [25]. When a seed was found to be germinated, it was recorded as such, along with the day of germination, and then discarded.
A germination index (GI) was used to account for both the percentage and speed of germination [29]. Both the number of germinated seeds and the day at which germination occurred were integrated into the following equation:
where n1, n2 … n7 indicates the number of seeds germinated on each individual day up until day seven; 7, 6 …1 are weights given to seeds according to their germination date with those that germinated earlier receiving higher weights [29].
GI = (7 × n1) + (6 × n2) + … + (1 × n7)
2.2. Plant Growth Experiments
Three different growth experiments were conducted to determine the responses of cover crops to the presence of A. hybridus (10 replicates for each species and each treatment). They were tea solution, dried ground material and fresh trimmed material. For all growth experiments, the commercial potting soil used was SunGro Sunshine® Mix 1; 1 L pots were filled according to the treatments (see below), soaked with water, and allowed to sit overnight, before planting. Seeds of each of the cover crops were sown individually in a pot at a depth specific to each species (provided by supplier), and the pots were checked daily for soil dryness and watered as necessary (i.e., when the top of the soil felt dry to touch). In all cases, plants were harvested 35 days after initial sowing and freshly harvested plants were separated between shoot and root and the organs dried in a plant dryer set at 40˚C for approximately 14 days. The dry weight of each individual organ was then recorded. The mortality rate, i.e., the percentage of plants that died prior to harvest, was also considered in the fresh trimmed treatment experiment, as early plant death was observed in many of the replicates; it should be noted that plant mortality was not observed in the tea solution or dried ground treatments.
2.2.1. Tea
In the tea experiment, plants were exposed to one of five allelopathic tea concentrations: 0% (control), 25%, 50%, 75%, or 100%. Individual plants received 10 mL of the appropriate solution one week following emergence (after the first shoot was visible in 60% of replicates) and again once every ten days for a total of 30 mL solution. Solutions were applied to the soil without direct above-ground contact with the seedlings.
2.2.2. Dried ground
In the dried ground experiment, 1.5 g of the plant material (prepared as described above) was mixed into the soil prior to plant sowing. We chose this amount to reflect the amount of A. hybridus present in the field where collection occurred (~ 10% ground cover in a 1m x 1m plot, which works out to approximately 1.5g of dried material per 20 cm2). A control treatment that contained no addition of dried ground material was included for each species.
2.2.3. Fresh Trimmed
The fresh trimmed material was used in this experiment with the following amounts: 0g (control), 10g or 20g of fresh material (prepared as described above), each of which were thoroughly mixed into 800 g of soil per replicate (reflective of what would happen in the field where the 10g treatment would work out to approximately 10g of fresh material per 20 cm2 incorporated into the first ~ 3 cm of soil).
2.3. Statistical Analysis
In all experiments, seed germination indices or individual dry weights of both shoot and root of each plant were first tested to ensure normality and homogeneity of variance. The germination indices and the weight of plants exposed to tea solutions were each compared among treatments using a general linear model followed by a Tukey’s range post-hoc test. Plant weight of each cover crop grown in the soil treated with dried ground material was compared to that of control (water only), using an independent-samples t-test. Plant weights of those individuals grown in soil treated with fresh trimmed material were compared between treatments using a general linear model followed by a Scheffe post-hoc test. The Scheffe test was chosen as it is sensitive to type I error rates, particularly in unbalanced designs [30], which was the case in our trials due to the high mortality rates of some species in the 20g treatments. Mortality rates were included for the fresh trimmed experiment for interpretive purposes only, because many treatments did not yield enough viable plants for robust statistical analysis. Data analyses were performed using IBM SPSS Statistics version 22.0.
3. Results
3.1. Seed Germination Experiment
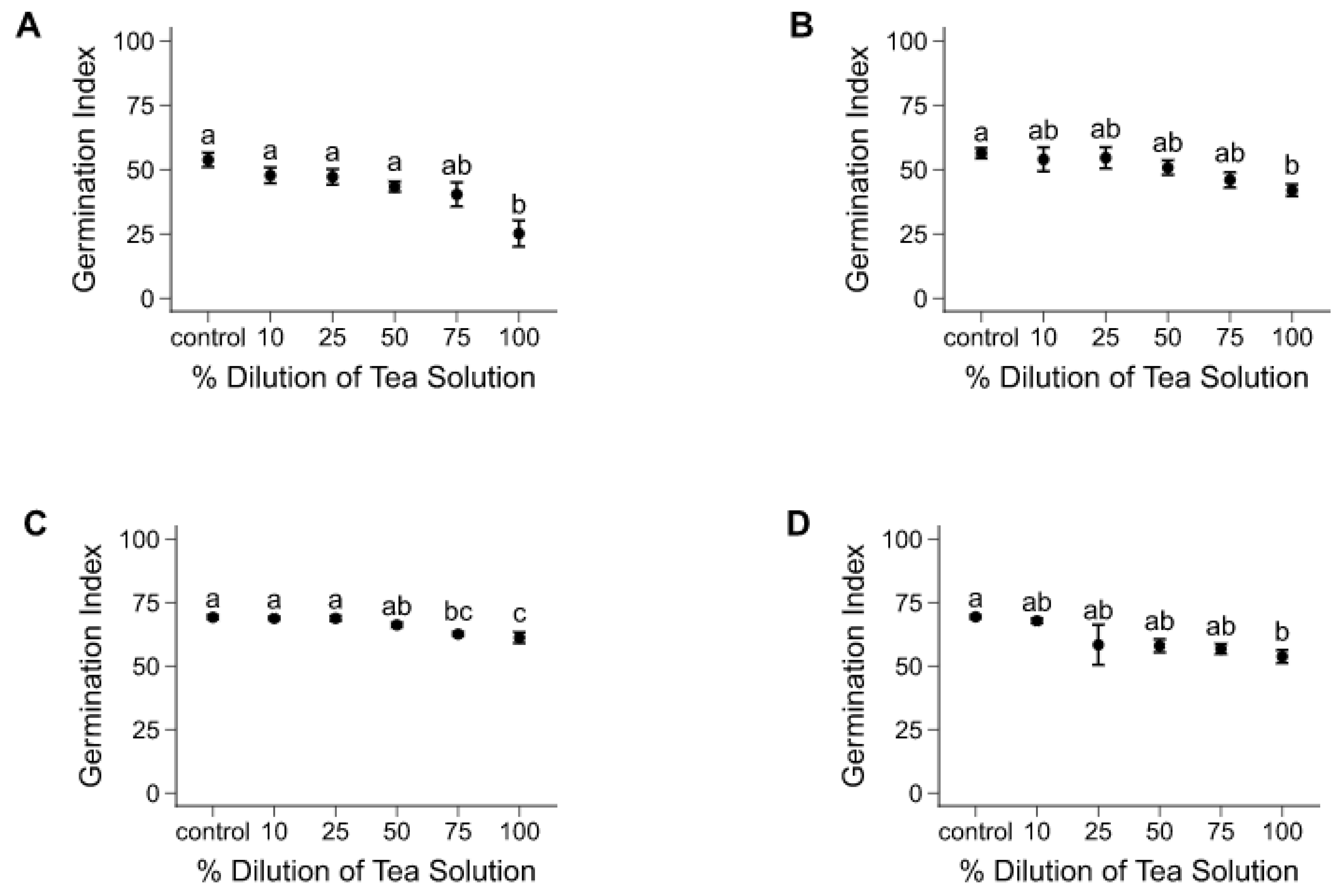
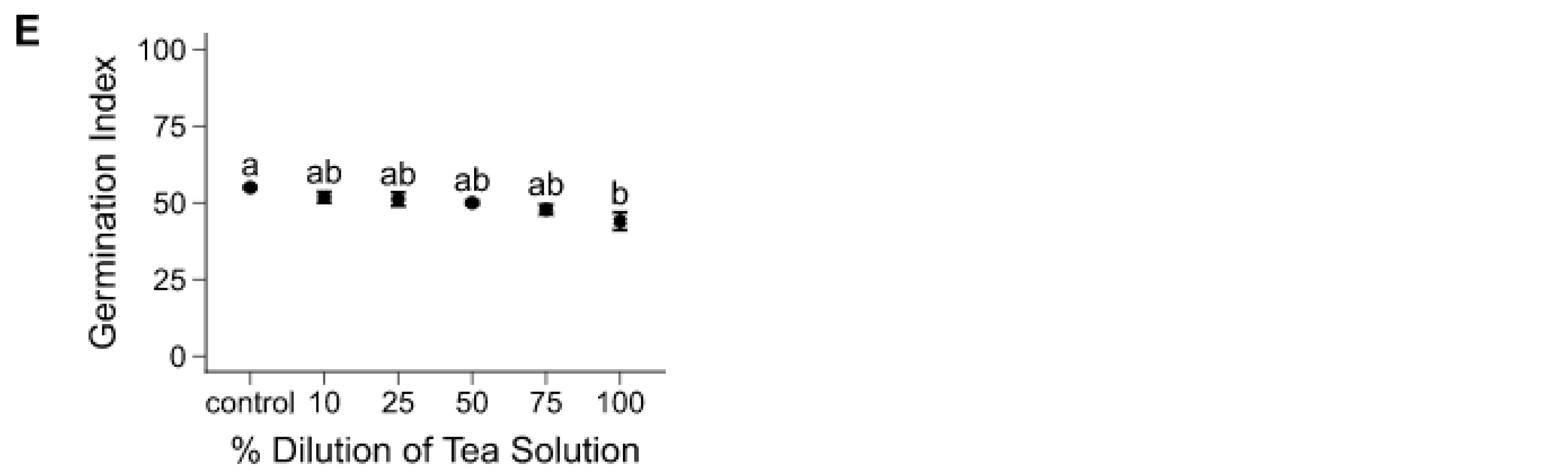
For most species, seed germination, expressed as GI, was significantly affected by the presence of A. hybridus tea solutions, especially at 100% solution (Figure 1). In all treatments, most seeds germinated by the seventh day. Mean GI trends indicated mainly a lower germination potential that varied in magnitude (from 1%–15% species and dose-dependant) with each increase in tea concentration. Chicory showed the most important decline in GIs (53% decrease between control and 100% dilution; Figure 1A). Red clover had the greatest sensitivity to the tea solution, as its seeds were affected by both 100% (12% GI decrease) and 75% (9% GI decrease) concentration relative to control (Figure 1C).
3.2. Growth Experiments
3.2.1. Tea
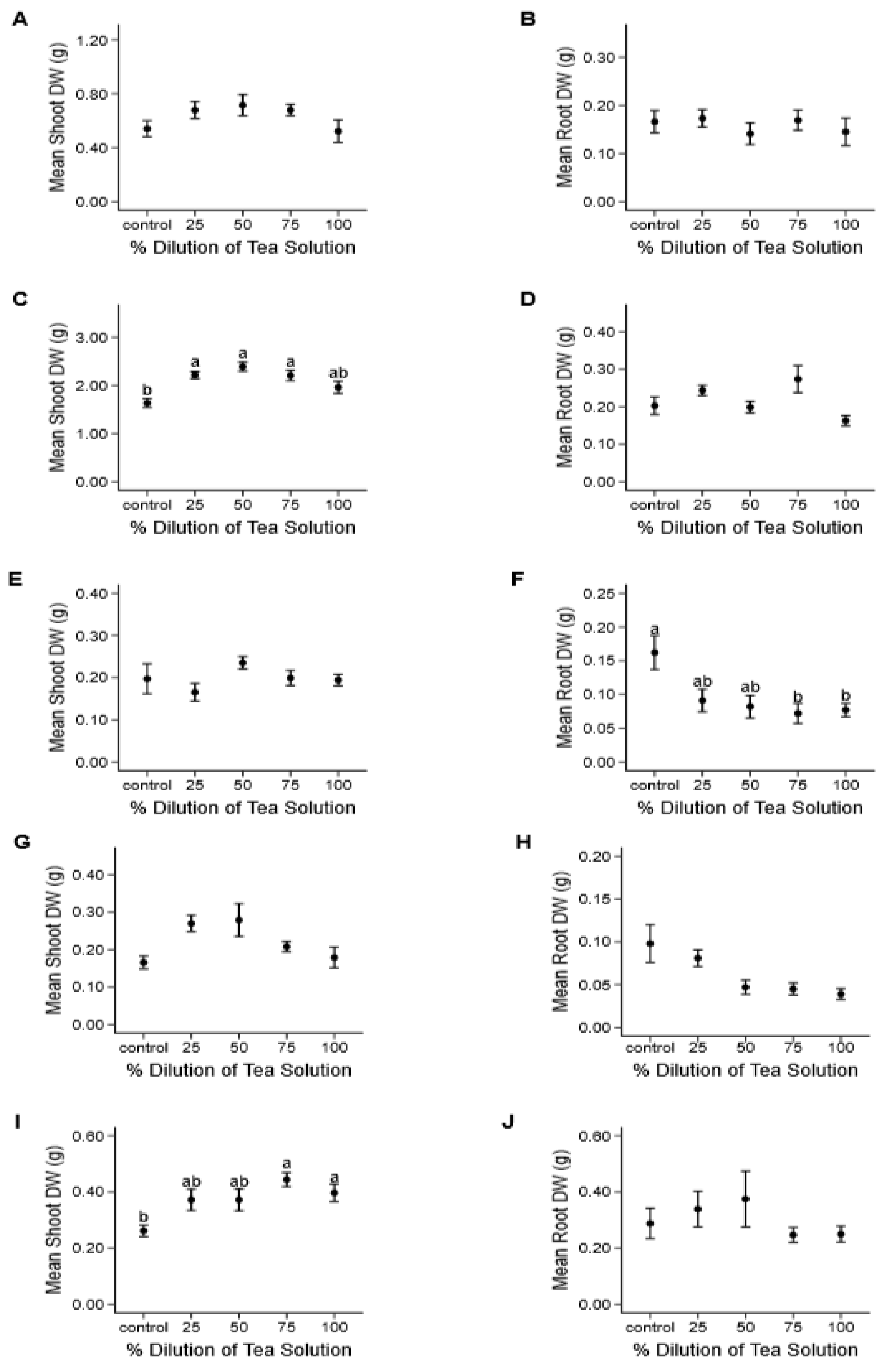
Shoot (Figure 2A,C,E,G,I) and root (Figure 2B,D,F,H,J) weights tended to vary slightly (< 1g) among tea dilutions, but most differences were not significant. Shoot weights of oilseed radish and ryegrass significantly increased (from control 1.63 ± 0.09g to 2.39 ± 0.09g in 50% dilution and from control 0.26 ± 0.02g to 0.44 ± 0.03g in 75% dilution, respectively), with increased concentrations of tea solutions suggesting a promoting effect (Figure 2C,I, respectively). Red clover root weight significantly decreased with increasing concentrations of tea solution (more than 50% in both 75 and 100% dilutions; Figure 2F).
3.2.2. Dried Ground Material
In general, dried ground material did not significantly affect shoot or root weight when compared to the control. Only the weights of red clover shoot and root and alfalfa root were significantly lower (up to 60%) than the control, when the plants were exposed to dried ground material (Table 1). On the contrary, annual ryegrass showed a significant increase in shoot weight when exposed to dried ground material (from 0.26 ± 0.02g in control to 0.58 ± 0.06g in treated). Similarly, oilseed radish root weight significantly increased (from control 0.20 ± 0.02 to 0.30 ± 0.04g in treated) when exposed to dried ground material (Table 1).
3.2.3. Fresh Material
Mortality of plants was observed in chicory (60%), red clover (40%) and alfalfa (30%) when exposed to 20 g of fresh material (Table 2). Shoots of chicory, oilseed radish and alfalfa showed significant decreases in weights when exposed to fresh material of A. hybridus incorporated in the soil (up to 58%, 47%, and 40% declines, respectively; Table 2). A similar decline (up to 48%) was also observed for oilseed radish and chicory roots, although for chicory the trend was not as obvious (a decline of 20% between control and 20g treatment), because of the mortality-caused loss of individuals (60%) in the highest concentration (Table 2).
4. Discussion
Plant allelopathy has the potential to play both positive and negative roles in agricultural settings, but surprisingly only a few studies have investigated allelopathic interactions of cover crops in perennial agroecosystems. In this study, we assessed the germination potential and plant growth, reflected by weight, of selected cover crop species exposed to various concentrations of A. hybridus tea solution and dried ground or fresh plant materials incorporated into the soil. Both germination and plant weight responses to A. hybridus vary among species, organs, and delivery-modes.
Germination potential decreased for all cover crop species exposed to A. hybridus tea extract treatments. These results agree with those obtained for A. retroflexus leaf extracts on garden cress (Lepidium sativum L.) [31] and maize (Zea mays L.) [32]. In our germination experiment, alfalfa and red clover were both negatively affected by increasing tea concentration. Slender amaranth (Amaranthus viridis L.) extract significantly decreases and delays the germination of mung bean (Vigna radiata (L.) R. Wilczek) [33]. Similarly, A. retroflexus extracts significantly inhibit the germination of soybean, common vetch (Vicia sativa L.), and pea, up to 58%, 75%, and 100%, respectively [34]. Our results indicate that A. hybridus is also detrimental to the germination of leguminous species. However, we only tested germination using tea solutions. It is possible that the plant material in the soil may have a different impact on germination from that of the solution. Germination of species, such as sunflower and soybean, are inhibited by A. retroflexus shoot residues [35]. Future experiments should examine how newly transplanted seedlings respond to A. hybridus. This may also reduce the mortality encountered for some species exposed to fresh materials in this study.
On one hand, A. hybridus tea extract has the potential to negatively impact the growth of cover crop species, as was shown in red clover root weight. Such negative impacts have been observed in other species, such as for root and shoot biomasses of red, white, and pinto beans (Phaseolus vulgaris L.) [13]. In the present work, the inhibitory effect was observed in the legumes, whereby the dry weights of the shoot and root of red clover and that of the root of alfalfa decreased when the A. hybridus was added as a dried ground material, and the dry weight of the alfalfa shoot weight declined when A. hybridus was added as fresh material. However, a few of our cover crop species did not respond to the presence of the different types of A. hybridus extracts. The lack of response of chicory to both tea solutions and dried ground material, but its significant decline in both root and shoot weights when exposed to fresh material, suggested that the active chemical ingredients in these materials might differ and had differential effect on the responses of plants. Singh et al. [36] found that the mode of delivery often changes the potency of allelochemicals and, as a result, the responses of exposed plants. It is important to note that both dead and live plant materials can be found at different times of the year in the fields, as soil is plowed on a regular basis in vineyards. Further studies would be needed to better understand the influence of the chemicals in dried versus fresh materials and their interactions with soil and plants.
On the other hand, A. hybridus may be beneficial to the growth of some cover crops. Oilseed radish and annual ryegrass shoot, for example, responded positively to the tea solution, while oilseed radish roots benefited from the addition of dried material. However, both shoot and root weights of oilseed radish declined when the plants were grown in the presence of fresh material. The existence of beneficial allelopathic effects of donor plant extracts, even at high concentrations, on receiver plant growth is well documented [37] and can support some of our findings. The type of materials therefore may play a role in affecting plant growth. Cabbage (Brassica oleracea L.) and annual sunflower biomass are not affected by fresh residues of Amaranthus palmeri, but dried residues lead to biomass decrease in both species [38]. A. retroflexus dry shoot residue incorporated in soil has an adverse effect on several species (i.e., cabbage, carrot, eggplant, pepper, and cauliflower). However, again, fresh residues are more potent [39].
The stimulatory effects observed in some of our species could be attributed to hormesis. Hormesis occurs when plant growth increases under low levels (concentrations) of exposure to otherwise toxic plant extracts [40,41]. This phenomenon is an evolutionary response mechanism that takes place when plants produce secondary metabolites as a response to environmental toxin exposure, a process that stimulates plant growth [42]. Indeed, in the case of oilseed radish treated with tea, we observed an increase in shoot weight at low concentrations, but no change in shoot weight when plants were exposed to higher concentrations. Hernández-Aro et al. [43] report that the shoot growth of radish (variety not indicated) is stimulated by a 50% concentration of organ extract from creeping-oxeye (Sphagneticola trilobata L.), relative to 100%. However, radish (var. Champion) shoot growth is unaffected by extract (tea) from prince’s feather (Amaranthus hypochondriacus L.) but decreases when the plant is grown in soil amended with even small amounts of A. hypochondriacus plant residue [43].
The conditions in which such experiments are conducted may also be important to consider. Our experiments were conducted under controlled conditions in the greenhouse. Red amaranth (Amaranthus cruentus L) extracts reduce tomato and cabbage seed germination in pot trials, but not in field trials [44]. This is likely due to confounding effects of other environmental conditions and potential interactions with other plant species and even soil microorganisms in the field. Our study, like others, emphasizes how weed allelochemical interactions with plants can be complex depending on species, weed material and environmental conditions.
Allelopathic pressure is not always delivered by a weed species to a crop. Quite the opposite, crop species can also have allelopathic effects on weed species. Wild radish soil residue inhibits yellow nutsedge (Cyperus esculentus L.), prickly sida (Sida spinosa L.) and sicklepod (Senna obtusifolia L.) growth, by up to 98% [45]. Furthermore, forage radish and black oat significantly reduce A. retroflexus biomass in field experiments [7]. Results such as these suggest that interactions between weeds and field cultivars are not simple cause-and-effect relationships, but they are rather highly complex interactions that change according to plant type, growth stage, organ residue present, and a combination of variables (i.e., microbes, soil nutrients, weather, etc. [19,26]), beyond that of just the plants themselves. Control studies that look at the reciprocal effects that each weed species and crop cultivar have on one another are therefore needed. Further in situ field analysis will also help determine site-specific parameters that may influence the allelopathic behaviour of plant communities in agroecosystems. Understanding how difficult-to-control weeds such as A. hybridus may affect the performance of cover crop species or how they in turn may be affected by cover crops is essential in developing sound agricultural weed management strategies. Studying these interactions through different types of extract and under controlled and field conditions may enhance our understanding of the mechanisms that affect plant responses. Such an understanding is essential to developing sustainable ways of providing food and materials for future generations.
Author Contributions
Conceptualization, H.V. and L.V.; methodology, H.V. and L.V.; validation, L.V.; formal analysis, H.V.; investigation, H.V.; writing—original draft preparation, H.V., F.C.G. and L.V.; writing—review and editing, F.C.G. and L.V.; supervision, F.C.G. and L.V.; funding acquisition, H.V., F.C.G. and L.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Ontario-China Research and Innovation Fund (OCRIF) grant number 3230, with additional funding provided by the Government of Ontario Graduate Scholarship program.
Acknowledgments
The authors would like to thank Ed Hughes (Beamsville, ON) for allowing us to retrieve plants from his vineyard and for supplying seeds, Canadian Comfort Alpacas (Fenwick, ON) for supplying seeds, Paulina Fedko, Alannah Mosimann, and Kasia Zgurzynski for their assistance with project implementation and maintenance, as well as Fiona Hunter (Brock University) and Kevin Stevens (Wilfrid Laurier University), for their valuable comments during the initial stages of the manuscript. The authors would also like to thank the two anonymous reviewers for their constructive comments which made the manuscript stronger.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1988; pp. 52–97. [Google Scholar]
- Bedoussac, L.; Journet, E.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Gitari, H.I.; Karanja, N.N.; Gachene, C.K.K.; Kamau, S.; Sharma, K.; Schulte-Geldermann, E. Nitrogen and phosphorous uptake by potato (Solanum tuberosum L.) and their use efficiency under potato-legume intercropping systems. Field Crops Res. 2018, 222, 78–84. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, A.; Wang, F.; Han, X.; Wang, D.; Li, S. Arbuscular mycorrhizal fungi and Rhizobium facilitate nitrogen uptake and transfer in soybean/maize intercropping system. Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razze, J.M.; Liburd, O.E.; Webb, S.E. Intercropping buckwheat with squash to reduce insect pests and disease incidence and increase yield. Agroecol. Sustain. Food 2016, 40, 863–891. [Google Scholar] [CrossRef]
- Adler, M.J.; Chase, C.A. Comparison of the allelopathic potential of leguminous summer cover crops: Cowpea, sunn hemp, and velvet bean. J. Hortic. Sci. Biotech. 2007, 42, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Gfeller, A.; Herrera, J.M.; Tschuy, F.; Wirth, J. Explanations for Amaranthus retroflexus growth suppression by cover crops. J. Crop Prot. 2018, 104, 11–20. [Google Scholar] [CrossRef]
- Florence, A.M.; Higley, L.G.; Drijber, R.A.; Francis, C.A.; Lindquist, J.L. Cover crop mixture diversity, biomass productivity, weed suppression, and stability. PLoS ONE 2019, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37, 1–17. [Google Scholar] [CrossRef]
- Duke, S.O. Proving allelopathy in crop-weed interactions. Weed Sci. 2015, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Rice, E. Allelopathy, 3rd ed.; Academic Press: Cambridge, MA, USA, 1984; pp. 1–4. [Google Scholar]
- Skinner, E.M.; Diaz-Perez, J.C.; Schomber, H.H.; Vencill, W. Allelopathic effects of sunnhemp (Crotalaria juncea L.) on germination of vegetables and weeds. Hort. Sci. 2012, 47, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Kalinova, S.; Golubinova, I.; Hristoskov, A.; Ilieva, A. Allelopathic effect of aqueous extract from root systems of Johnson grass on seed germination and initial development of soybean, pea and vetch. Ratar. Povrt. 2012, 49, 250–256. [Google Scholar] [CrossRef]
- Carvalho, M.S.S.; Andrade-Vieira, L.F.; Santos, F.E.; Correa, F.F.; Cardoso, M.G.; Vilela, L.R. Allelopathic potential and phytochemical screening of ethanolic extracts from five species of Amaranthus spp. in the plant model Lactuca sativa. Sci. Hortic. 2019, 245, 90–98. [Google Scholar] [CrossRef]
- Wazir, L.; Sadiq, M.; Baloch, M.; Awan, I.; Khan, E.; Shah, I. Application of bio-herbicide alternatives for chemical weed control in rice. Pak. J. Weed Sci. Res. 2011, 17, 245–252. [Google Scholar]
- Mushtaq, M.; Cheema, Z.; Khaliq, A. Effects of mixture of allelopathic plant aqueous extracts on Trianthema portulacastrum L. weed. Allelopath. J. 2010, 25, 205–212. [Google Scholar]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H. Growth and physiological changes in continuously cropped eggplant (Solanum melongena L.) upon relay intercropping with garlic (Allium sativum L.). Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Riaz, T.; Khan, S.N.; Javaid, A. Management of Fusarium corm rot of gladiolus (Gladiolus grandiflorus sect. Blandus cv. Aarti) by using leaves of allelopathic plants. Afr. J. Biotechnol. 2010, 9, 4681–4686. [Google Scholar]
- Kruse, M.; Strandberg, M.; Strandberg, B. Ecological Effects of Allelopathic Plants-a Review; NERI Technical Report No. 315; National Environmental Research Institute: Silkeborg, Denmark, 2000; p. 66. [Google Scholar]
- Yanyan, G.; Wang, Q.; Wang, L.; Liu, W.; Liu, X.; Huang, Y.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2018, 64, 708–717. [Google Scholar] [CrossRef]
- Ontario Ministry of Agriculture, Food, and Rural Affairs. Pigweeds (Redroot, Green and Smooth); Factsheet Order#:01-009; Queen’s Printer for Ontario: Toronto, ON, Canada, 2009.
- Costea, M.; Weaver, S.E.; Tardif, F.J. The biology of Canadian weeds. 130. Amaranthus retroflexus L., A. powellii S. Watson and A. hybridus L. Can. J. Plant Sci. 2004, 84, 631–668. [Google Scholar] [CrossRef]
- Bakhshayeshan-Agdam, H.; Salehi-Lisar, S.Y.; Motafakkerazad, R.; Talebpour, A.; Farsad, N. Allelopathic effects of redroot pigweed (Amaranthus retroflexus L.) on germination & growth of cucumber, alfalfa, common bean and bread wheat. Acta Agric. Slov. 2015, 105, 193–202. [Google Scholar] [CrossRef]
- Baličević, R.; Ravlić, M.; Čuk, P.; Šević, N. Allelopathic effect of three weed species on germination and growth of onion cultivars. In Proceedings of the 8th International Scientific/Professional Conference Agriculture in Nature and Environment Protection, Vukovar, Croatia, 1–3 June 2015. [Google Scholar]
- Shahrokhi, S.; Hejazi, S.N.; Khodabandeh, H.; Farboodi, M.; Faramarzi, A. Allelopathic effect of aqueous extracts of pigweed, Amaranthus retroflexus L. organs on germination and growth of five barley cultivars. In Proceedings of the 3rd International Conference on Chemical, Biological and Environmental Engineering, Chengdu, China, 23 September 2011; IACSIT Press: Singapore, 2011; Volume 20, pp. 80–84. [Google Scholar]
- De Souza, M.C.; de Carvalho, L.B.; da Costa Aguiar Alves, P.L.; Fidelis Giancotti, P.R. Allelopathy in pigweed (a review). Can. J. Plant Sci. 2011, 1, 5–12. [Google Scholar]
- Trucco, F.; Hager, A.; Tranel, P.J. Acetolactate synthase mutation conferring imidazolinone-specific herbicide resistance in Amaranthus hybridus. J. Plant Physiol. 2006, 163, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, A.I.; Pueyo, Y.; Giner, M.L.; Foronda, A.; Sanchez-Navarrete, P.; Saiz, H.; Alado, C.L. Evidence for chemical interference effect of an allelopathic plant on neighboring plant species: a field study. PLoS One 2018, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kader, M.A. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proc. R. Soc. New South Wales 2005, 138, 65–75. [Google Scholar]
- Lee, S.L.; Lee, D.K. What is the proper way to apply the multiple comparison test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Mlakar, S.G.; Jakop, M.; Bavec, M.; Bavec, F. Allelopathic effects of Amaranthus retroflexus and Amaranthus cruentus extracts on germination of garden cress. Afr. J. Agric. Res. 2012, 7, 1492–1497. [Google Scholar] [CrossRef]
- Konstantinović, B.; Blagojević, M.; Konstantinović, B.; Samardžić, N. Allelopathic effect of weed species Amaranthus retroflexus L. on maize seed germination. Rom. Agric. Res. 2014, 31, 315–321. [Google Scholar] [CrossRef]
- Sheeba, M.A.; Mahesh, B.; Devi Chinmayee, M.; Swapna, T.S.; Mini, I. The allelopathic potential of the weed Amaranthus viridis L. on germination and pigment profile of a pulse crop Vigna radiata (L.) R. Wilsczek. J. Aqu. Biol. Fish 2014, 2, 3344–3350. [Google Scholar]
- Marinov-Serafimov, P. Determination of allelopathic effect of some invasive weed species on germination and initial development of grain legume crops. Pestic. Phytomed. 2010, 25, 251–259. [Google Scholar] [CrossRef]
- Béres, I.; Kazinezi, G. Allelopathic effects of shoot extracts and residues of weeds on field crops. Allelopath. J. 2000, 7, 93–98. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathy in agroecosystems. J. Crop Prod. 2008, 4, 1–41. [Google Scholar] [CrossRef]
- Subtain, M.U.; Hussain, M.; Tabassam, M.A.R.; Ali, M.A.; Ali, M.; Mohsin, M.; Mubushar, M. Role of allelopathy in the growth promotion of plants. J. Agric. Sci. 2014, 2, 141–145. [Google Scholar] [CrossRef]
- Menges, R.M. Allelopathic effects of Palmer Amaranth (Amaranthus palmeri) and other plant residues in soil. Weed Sci. 1987, 35, 339–347. [Google Scholar] [CrossRef]
- Qasem, J.R. Allelopathic effects of Amaranthus retroflexus and Chenopodium murale on vegetable crops. Allelopath. J. 1995, 2, 49–66. [Google Scholar]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Chauhan, B.S. Can hormesis of plant-released phytotoxins be used to boost and sustain crop production? Crop Prot. 2017, 93, 69–76. [Google Scholar] [CrossRef]
- Southam, C.M.; Ehrlich, J. Effects of extract of western red-cedar heartwood on certain wood-decaying fungi in culture. Phytopathology 1943, 33, 517–524. [Google Scholar]
- Belz, R.G.; Hurle, K. Dose-response-a challenge for allelopathy? Nonlinearity Biol. Toxicol. Med. 2005, 3, 173–211. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Aro, M.; Hernández-Pérez, R.; Guillén-Sánchez, D.; Torres-Garcia, S. Allelopathic influence of residues from Sphagneticola trilobata on weeds and crops. Planta Daninha 2016, 33, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Prinsloo, G.; Plooy, C.P.D. The allelopathic effects of Amaranthus on seed germination, growth and development of vegetables. Biol. Agric. Hortic. 2018, 34, 268–279. [Google Scholar] [CrossRef]
- Norsworthy, J.K. Allelopathic potential of wild radish. Weed Technol. 2003, 17, 307–313. [Google Scholar] [CrossRef]
Figure 1.
Effects of A. hybridus extracts on mean germination index (± SE), after seven days of seed exposure. General linear models were performed on the five treatments and control for each individual species, including: (A) chicory, (B) oilseed radish, (C) red clover, (D) alfalfa, and (E) annual ryegrass. Significant differences among treatments are represented by different lowercase letters in (A) F(5, 54) =7.470, p = < 0.001, (B) F(5, 54) = 2.902, p = 0.035, (C) F(5, 54) = 9.348, p = < 0.001, (D) F(5, 54) = 3.010, p = 0.030, and (E) F(5, 54) = 4.335, p = 0.006.
Figure 1.
Effects of A. hybridus extracts on mean germination index (± SE), after seven days of seed exposure. General linear models were performed on the five treatments and control for each individual species, including: (A) chicory, (B) oilseed radish, (C) red clover, (D) alfalfa, and (E) annual ryegrass. Significant differences among treatments are represented by different lowercase letters in (A) F(5, 54) =7.470, p = < 0.001, (B) F(5, 54) = 2.902, p = 0.035, (C) F(5, 54) = 9.348, p = < 0.001, (D) F(5, 54) = 3.010, p = 0.030, and (E) F(5, 54) = 4.335, p = 0.006.


Figure 2.
Effects of A. hybridus organ extracts on mean individual shoot and root dry weight (± SE), after a 35-day growth period. General linear models were performed on the four extract treatments and control for chicory (A and B), oilseed radish (C and D), red clover (E and F), alfalfa (G and H), and annual ryegrass (I and J). Significant differences among treatments are represented by different lowercase letters in (C) oilseed radish shoot (F(4, 45) = 8.658, p = < 0.001), (F) red clover root (F (4, 45) = 4.593, p = 0.003), and (I) annual ryegrass shoot (F(4, 45) = 4.497, p = 0.004).
Figure 2.
Effects of A. hybridus organ extracts on mean individual shoot and root dry weight (± SE), after a 35-day growth period. General linear models were performed on the four extract treatments and control for chicory (A and B), oilseed radish (C and D), red clover (E and F), alfalfa (G and H), and annual ryegrass (I and J). Significant differences among treatments are represented by different lowercase letters in (C) oilseed radish shoot (F(4, 45) = 8.658, p = < 0.001), (F) red clover root (F (4, 45) = 4.593, p = 0.003), and (I) annual ryegrass shoot (F(4, 45) = 4.497, p = 0.004).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of A. hybridus dried ground plant material treatments on mean individual shoot and root dry weight (± SE), after a 35-day growth period. Independent samples t-tests on individual root and shoot dry weight were performed between the ground plant treatment and control (soil with no dried ground plant material added).
Table 1.
Effects of A. hybridus dried ground plant material treatments on mean individual shoot and root dry weight (± SE), after a 35-day growth period. Independent samples t-tests on individual root and shoot dry weight were performed between the ground plant treatment and control (soil with no dried ground plant material added).
| Species | Organ | Control (g) | Ground (g) | df | T | P |
|---|---|---|---|---|---|---|
| M | M | |||||
| Chicory | Shoot | 0.54 ± 0.06 | 0.73 ± 0.05 | 18 | 1.92 | 0.072 |
| Root | 0.17 ± 0.02 | 0.17 ± 0.03 | 18 | 0.17 | 0.866 | |
| Oilseed | Shoot | 1.63 ± 0.29 | 1.90 ± 0.38 | 18 | 1.77 | 0.095 |
| Radish | Root | 0.20 ± 0.07 | 0.30 ± 0.12 | 18 | 2.24 | 0.038 |
| Red | Shoot | 0.20 ± 0.04 | 0.10 ± 0.02 | 18 | 2.38 | 0.028 |
| Clover | Root | 0.16 ± 0.03 | 0.03 ± 0.01 | 18 | 4.80 | 0.001 |
| Alfalfa | Shoot | 0.17 ± 0.02 | 0.16 ± 0.04 | 18 | 0.10 | 0.924 |
| Root | 0.10 ± 0.02 | 0.04 ± 0.01 | 18 | 2.60 | 0.024 | |
| Annual | Shoot | 0.26 ± 0.02 | 0.58 ± 0.06 | 18 | 4.95 | <0.001 |
| Rye | Root | 0.29 ± 0.05 | 0.33 ± 0.03 | 18 | 0.68 | 0.506 |
abbreviations indicate mean (M), degrees of freedom (df), critical value of t (T) and calculated probability (p).
Table 2.
Effects of A. hybridus fresh trimmed plant material treatments on plant mortality rate and mean individual shoot and root dry weight (± SE), after a 35-day growth period. General linear models were performed on individual root and shoot dry weight, between the two plant material treatments and control. Significant differences among treatments are represented by lower case letters.
Table 2.
Effects of A. hybridus fresh trimmed plant material treatments on plant mortality rate and mean individual shoot and root dry weight (± SE), after a 35-day growth period. General linear models were performed on individual root and shoot dry weight, between the two plant material treatments and control. Significant differences among treatments are represented by lower case letters.
| Species | A. hybridus Treatment | Mortality Rate (%) | Shoot (g) | MS | F | P | Root (g) | MS | F | P |
|---|---|---|---|---|---|---|---|---|---|---|
| Chicory | 0g | 0 | 1.20 ± 0.63 a | 0.56 ± 0.04 a | ||||||
| 10g | 0 | 0.68 ± 0.09 b | 0.97 | 12.40 | < 0.001 | 0.29 ± 0.05 b | 0.19 | 7.87 | 0.003 | |
| 20g | 60 | 0.51 ± 0.21 b | 0.46 ± 0.12 ab | |||||||
| Oilseed | 0g | 0 | 2.99 ± 0.09 a | 0.88 ± 0.08 a | ||||||
| Radish | 10g | 0 | 2.01 ± 0.18 b | 5.27 | 37.05 | < 0.001 | 0.72 ± 0.06 ab | 0.32 | 8.79 | 0.001 |
| 20g | 0 | 1.57 ± 0.57 c | 0.52 ± 0.04 b | |||||||
| Red | 0g | 0 | 0.43 ± 0.05 | 0.29 ± 0.04 | ||||||
| Clover | 10g | 0 | 0.32 ± 0.04 | 0.03 | 2.09 | 0.147 | 0.23 ± 0.02 | 0.03 | 3.26 | 0.057 |
| 20g | 40 | 0.35 ± 0.04 | 0.18 ± 0.03 | |||||||
| Alfalfa | 0g | 0 | 0.59 ± 0.03 a | 0.27 ± 0.04 | ||||||
| 10g | 0 | 0.38 ± 0.06 b | 0.16 | 8.50 | 0.002 | 0.24 ± 0.02 | 0.01 | 1.18 | 0.325 | |
| 20g | 30 | 0.36 ± 0.03 b | 0.20 ± 0.02 | |||||||
| Annual | 0g | 0 | 0.81 ± 0.07 a | 0.50 ± 0.07 | ||||||
| Rye | 10g | 0 | 0.61 ± 0.09 b | 0.16 | 3.36 | 0.050 | 0.51 ± 0.07 | 0.06 | 1.14 | 0.334 |
| 20g | 0 | 0.57 ± 0.04 b | 0.38 ± 0.06 |
abbreviations indicate mean squares (MS), F ratio (F), and calculated probability (p).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
VanVolkenburg, H.; Guinel, F.C.; Vasseur, L. Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario. Agronomy 2020, 10, 529. https://doi.org/10.3390/agronomy10040529
AMA Style
VanVolkenburg H, Guinel FC, Vasseur L. Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario. Agronomy. 2020; 10(4):529. https://doi.org/10.3390/agronomy10040529
Chicago/Turabian StyleVanVolkenburg, Heather, Frédérique C. Guinel, and Liette Vasseur. 2020. "Impacts of Smooth Pigweed (Amaranthus hybridus) on Cover Crops in Southern Ontario" Agronomy 10, no. 4: 529. https://doi.org/10.3390/agronomy10040529
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.