Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of GOS Intake on Water Holding Capacity and TEWL in UVB-Irradiated Hairless Mice
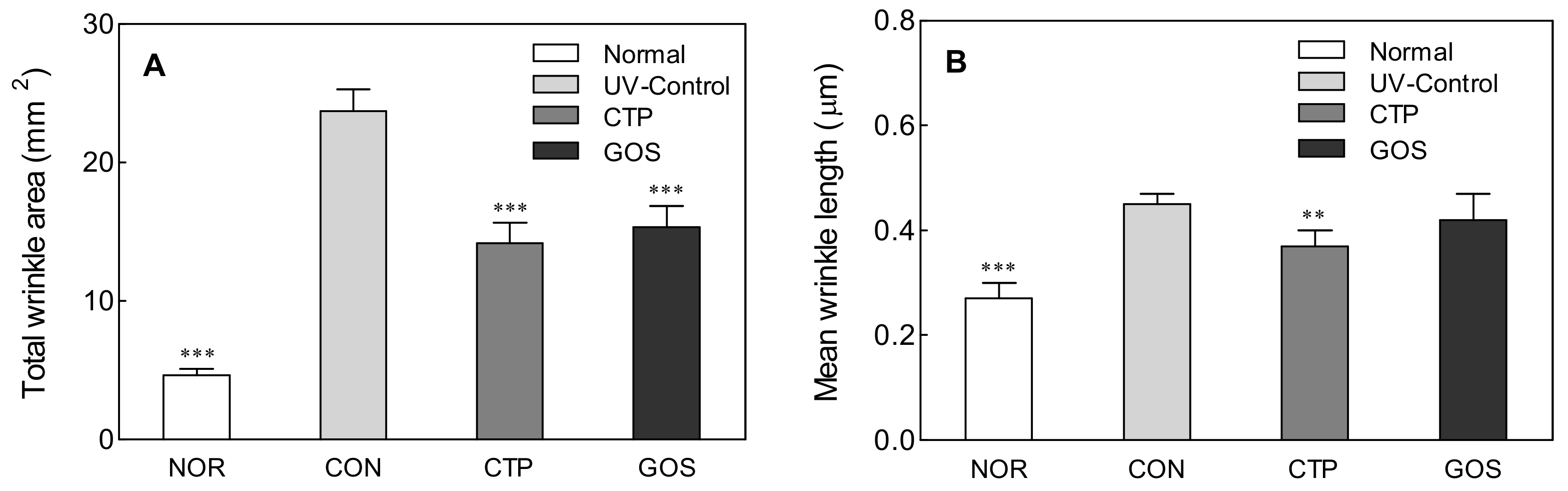
2.2. Effect of GOS Intake on Wrinkle Area and Mean Wrinkle Length in UVB-Irradiated Hairless Mice
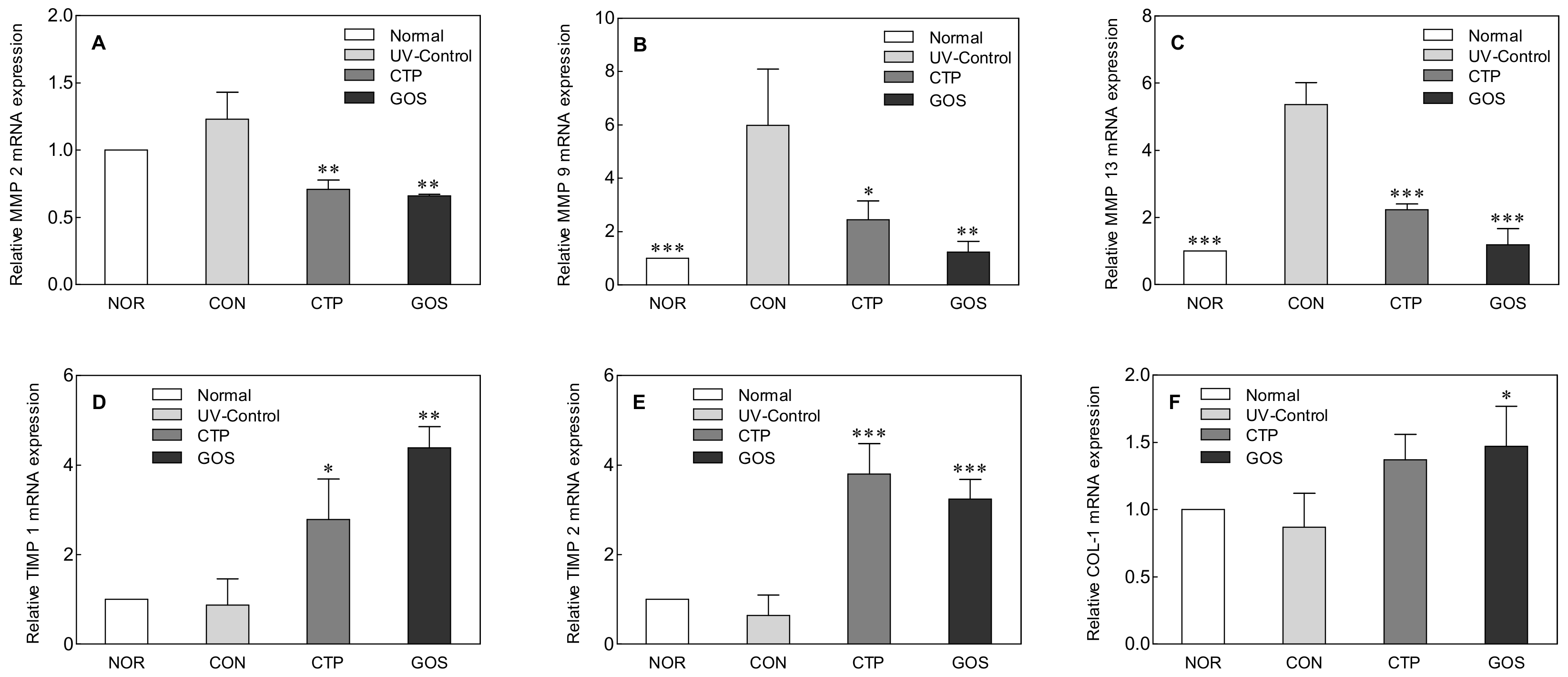
2.3. Effect of GOS Intake on MMPs, TIMP, and COL-1 Expression in UVB-Irradiated Hairless Mice
2.4. Effect of GOS Intake on Cytokine Levels in UVB-Irradiated Hairless Mice
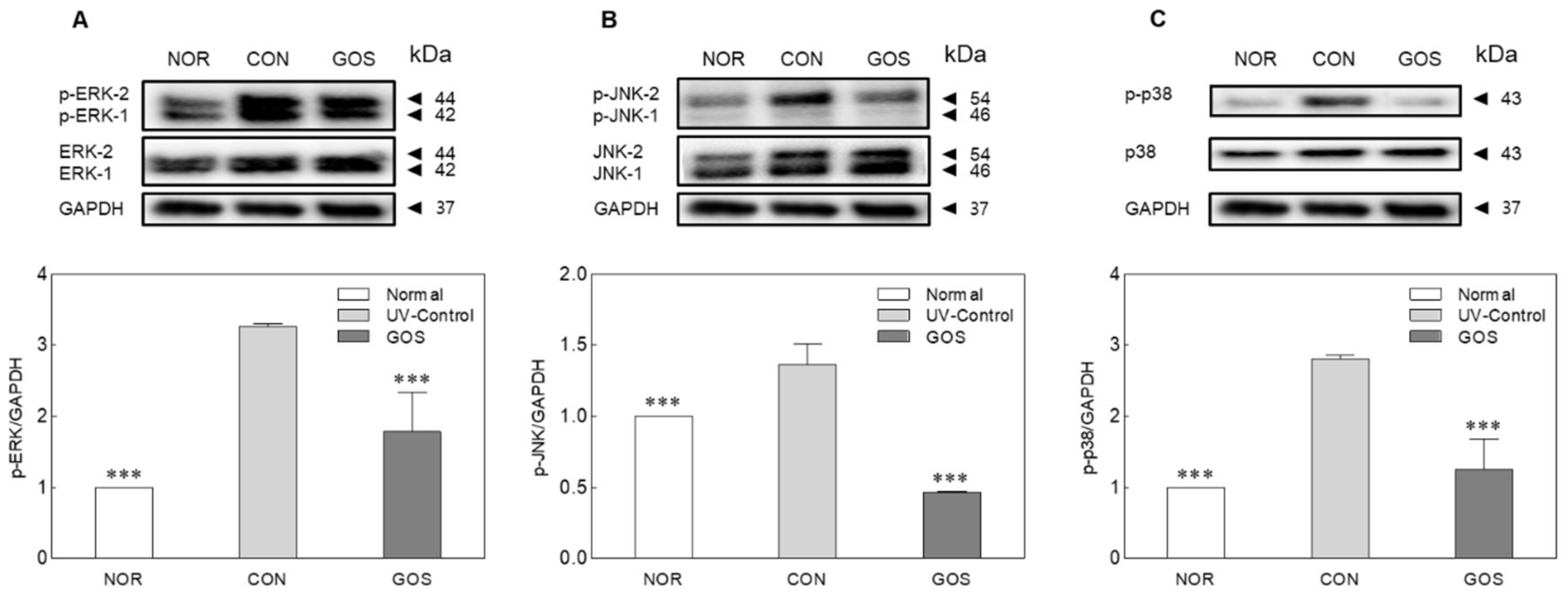
2.5. GOS Intake Prevents MAPK Phosphorylation in UVB-Irradiated Hairless Mice
3. Materials and Methods
3.1. Materials and Animals
3.2. UVB Exposure to Hairless Mice and Measurement of Skin Parameters
3.3. Gene Expression of Matrix Metalloproteinase and Collagen in Skin Tissue
3.4. Cytokine Assay for Skin Tissue
3.5. Western Blot Analysis
3.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maarouf, M.; Maarouf, C.; Yosipovitch, G.; Shi, V. The impact of stress on epidermal barrier function: An evidence-based review. Br. J. Dermatol. 2019, 181, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microb. 2018, 16, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, M.; Goldberg, D.; Aiello, A.; Larson, E.; Foxman, B. Skin microbiota: Microbial community structure and its potential association with health and disease. Infect. Genet. Evol. 2011, 11, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Takayama, H.; Morotomi, M.; Kuroshima, T.; Ueyama, S.; Matsumoto, K.; Kuroda, A.; Mutai, M. Effects of administration of TOS and Bifidobacterium breve 4006 on the human fecal flora. Bifidobacteria Microflora 1983, 2, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.; Lahtinen, S.; Tiihonen, K. The potential of probiotics and prebiotics for skin health. In Textbook of Aging Skin; Farage, M.A., Miller, K.W., Maibach, H.I., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2016; pp. 1299–1311. [Google Scholar]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Youn, S.Y.; Park, M.S.; Kim, H.G.; Baek, N.I.; Li, Z.; Ji, G.E. Synthesis of β-galactooligosaccharide using Bifidobacterial β-galactosidase purified from recombinant Escherichia coli. J. Microbiol. Biotechnol. 2017, 27, 1392–1400. [Google Scholar]
- Hong, K.-B.; Jeong, M.G.; Kim, J.H.; Park, Y.; Suh, H.J. Photoprotective effects of reinforced galactooligosaccharides supplementation against skin damage in hairless mice. FASEB J. 2014, 28, LB357. [Google Scholar]
- Hong, Y.H.; Chang, U.J.; Kim, Y.S.; Jung, E.Y.; Suh, H.J. Dietary galacto-oligosaccharides improve skin health: A randomized double blind clinical trial. Asia. Pac. J. Clin. Nutr. 2017, 26, 613–618. [Google Scholar]
- Tanabe, S.; Hochi, S. Oral administration of a galactooligosaccharide preparation inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. Int. J. Mol. Med. 2010, 25, 331–336. [Google Scholar] [CrossRef]
- Haratake, A.; Uchida, Y.; Schmuth, M.; Tanno, O.; Yasuda, R.; Epstein, J.H.; Elias, P.M.; Holleran, W.M. UVB-induced alterations in permeability barrier function: Roles for epidermal hyperproliferation and thymocyte-mediated response. J. Invest. Dermatol. 1997, 108, 769–775. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.J.; Chu, A.W.; Lu, Z.F.; Pan, M.H.; Che, D.F.; Zhou, X.J. Ultraviolet B-induced alterations of the skin barrier and epidermal calcium gradient. Exp. Dermatol. 2007, 16, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.H.; Chen, A.P.; Beno, D.W.A. Raf and mitogen-activated protein kinase regulate stellate cell collagen gene expression. J. Biol. Chem. 1996, 271, 11039–11042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.D.; Zhang, S.Q.; Zhang, L.; Li, B. Effect of orally administered collagen peptides from bovine bone on skin aging in chronologically aged mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, E.; Gomez, J.; Bilodeau, D. Beyond UV radiation: A skin under challenge. Int. J. Cosmet. Sci. 2013, 35, 224–232. [Google Scholar] [CrossRef]
- Zhan, J.Y.X.; Wang, X.F.; Liu, Y.H.; Zhang, Z.B.; Wang, L.; Chen, J.N.; Huang, S.; Zeng, H.F.; Lai, X.P. Andrographolide sodium bisulfate prevents uv-induced skin photoaging through inhibiting oxidative stress and inflammation. Mediators Inflamm. 2016, 2016, 3271451. [Google Scholar] [CrossRef] [Green Version]
- Inomata, S.; Matsunaga, Y.; Amano, S.; Takada, K.; Kobayashi, K.; Tsunenaga, M.; Nishiyama, T.; Kohno, Y.; Fukuda, M. Possible involvement of gelatinases in basement membrane damage and wrinkle formation in chronically ultraviolet B-exposed hairless mouse. J. Invest. Dermatol. 2003, 120, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Birkedalhansen, H. Proteolytic remodeling of extracellular-matrix. Curr. Opin. Cell Biol. 1995, 7, 728–735. [Google Scholar] [CrossRef]
- Presland, R.B. Function of filaggrin and caspase-14 in formation and maintenance of the epithelial barrier. Dermatol. Sinica 2009, 27, 1–14. [Google Scholar]
- Bellayr, I.H.; Mu, X.; Li, Y. Biochemical insights into the role of matrix metalloproteinases in regeneration: Challenges and recent developments. Future Med. Chem. 2009, 1, 1095–1111. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [Green Version]
- Quan, T.; Little, E.; Quan, H.; Qin, Z.; Voorhees, J.J.; Fisher, G.J. Elevated matrix metalloproteinases and collagen fragmentation in photodamaged human skin: Impact of altered extracellular matrix microenvironment on dermal fibroblast function. J. Invest. Dermatol. 2013, 133, 1362–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.F.; Gilliam, A.C.; Chen, G.F.; Tootell, E.; Cooper, K.D. In human skin, UVB initiates early induction of IL-10 over IL-12 preferentially in the expanding dermal monocytic/macrophagic population. J. Invest. Dermatol. 1998, 111, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, 1–63. [Google Scholar] [CrossRef] [Green Version]
- Delzenne, N.M.; Neyrinck, A.M.; Cani, P.D. Gut microbiota and metabolic disorders: How prebiotic can work? Br. J. Nutr. 2013, 109, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Erickson, K.L.; Hubbard, N.E. Probiotic immunomodulation in health and disease. J. Nutr. 2000, 130, 403–409. [Google Scholar] [CrossRef]
- Dow, W.F. The intestinal immune system. Gut 1989, 30, 1679–1685. [Google Scholar]
- Scharffetter-Kochanek, K.; Wlaschek, M.; Brenneisen, P.; Schauen, M.; Blaudschun, R.; Wenk, J. UV-induced reactive oxygen species in photocarcinogenesis and photoaging. Biol. Chem. 1997, 378, 1247–1257. [Google Scholar] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signalling. Oxid. Med. Cell. Longev. 2016, 2016, 1–18. [Google Scholar]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta Proteins Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Peus, D.; Vasa, R.A.; Beyerle, A.; Meves, A.; Krautmacher, C.; Pittelkow, M.R. UVB activates ERK1/2 and p38 signaling pathways via reactive oxygen species in cultured keratinocytes. J. Investig. Dermatol. 1999, 112, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suh, M.G.; Bae, G.Y.; Jo, K.; Kim, J.M.; Hong, K.-B.; Suh, H.J. Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway. Molecules 2020, 25, 1679. https://doi.org/10.3390/molecules25071679
Suh MG, Bae GY, Jo K, Kim JM, Hong K-B, Suh HJ. Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway. Molecules. 2020; 25(7):1679. https://doi.org/10.3390/molecules25071679
Chicago/Turabian StyleSuh, Min Geun, Gi Yeon Bae, Kyungae Jo, Jin Man Kim, Ki-Bae Hong, and Hyung Joo Suh. 2020. "Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway" Molecules 25, no. 7: 1679. https://doi.org/10.3390/molecules25071679