Mating Disruption of Helicoverpa armigera (Lepidoptera: Noctuidae) on Processing Tomato: First Applications in Northern Italy



 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Mating Disruption Trials Planning
2.3. Pheromone Treatments
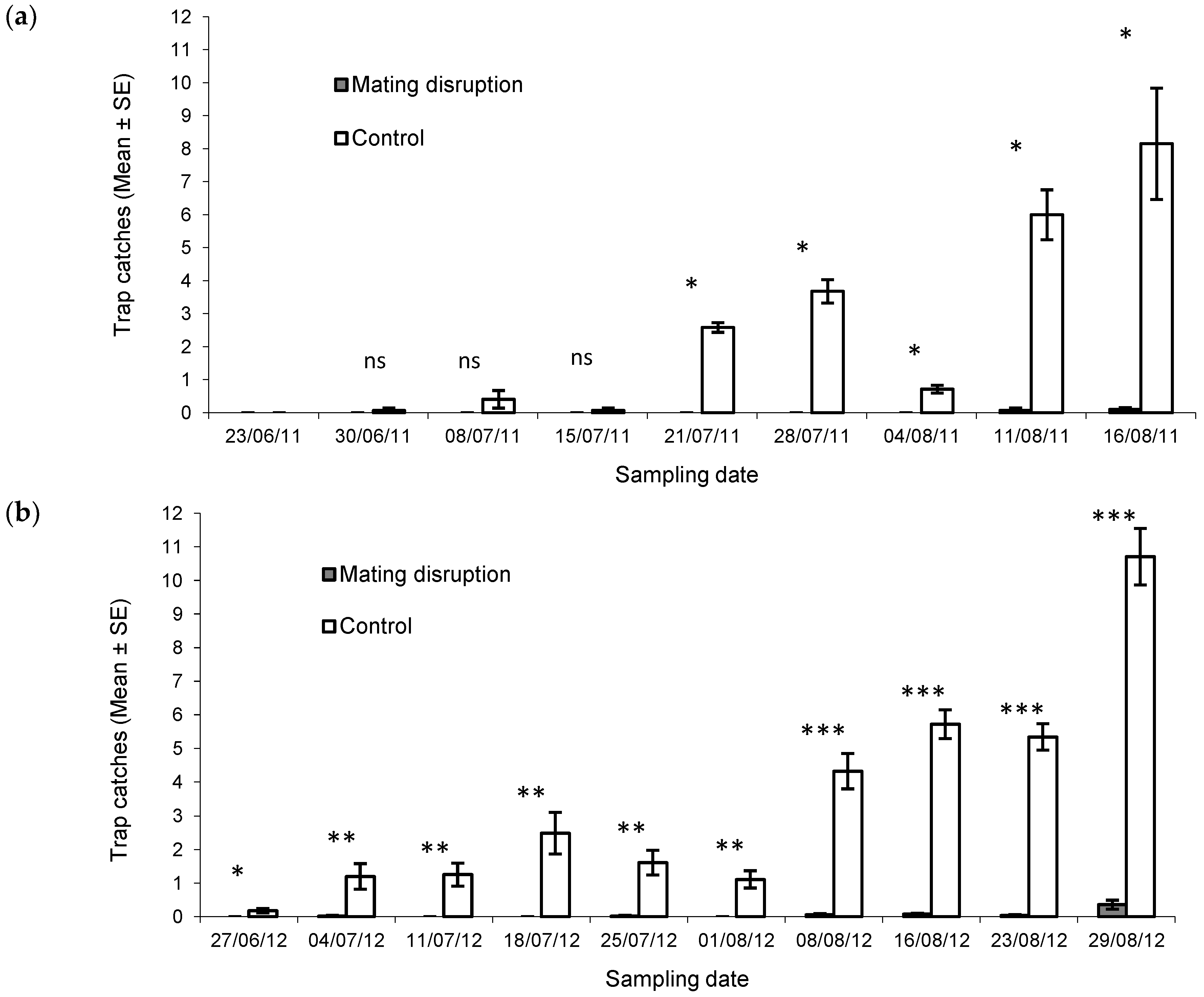
2.4. Male Capture Evaluation
2.5. Fruit Damage Estimation
2.6. Data Analysis
2.6.1. Male Capture Analysis
2.6.2. Fruit Damage Assessment
3. Results
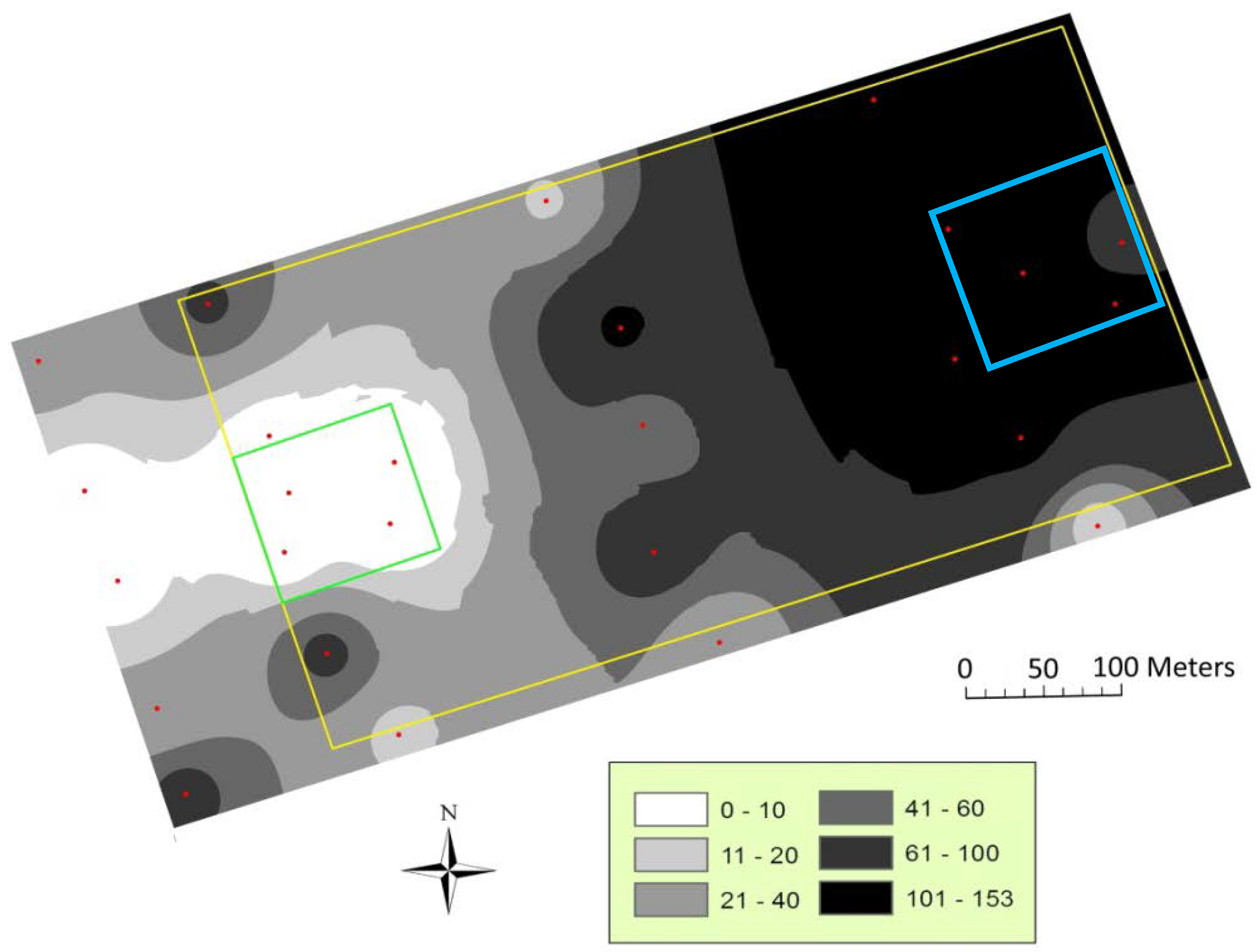
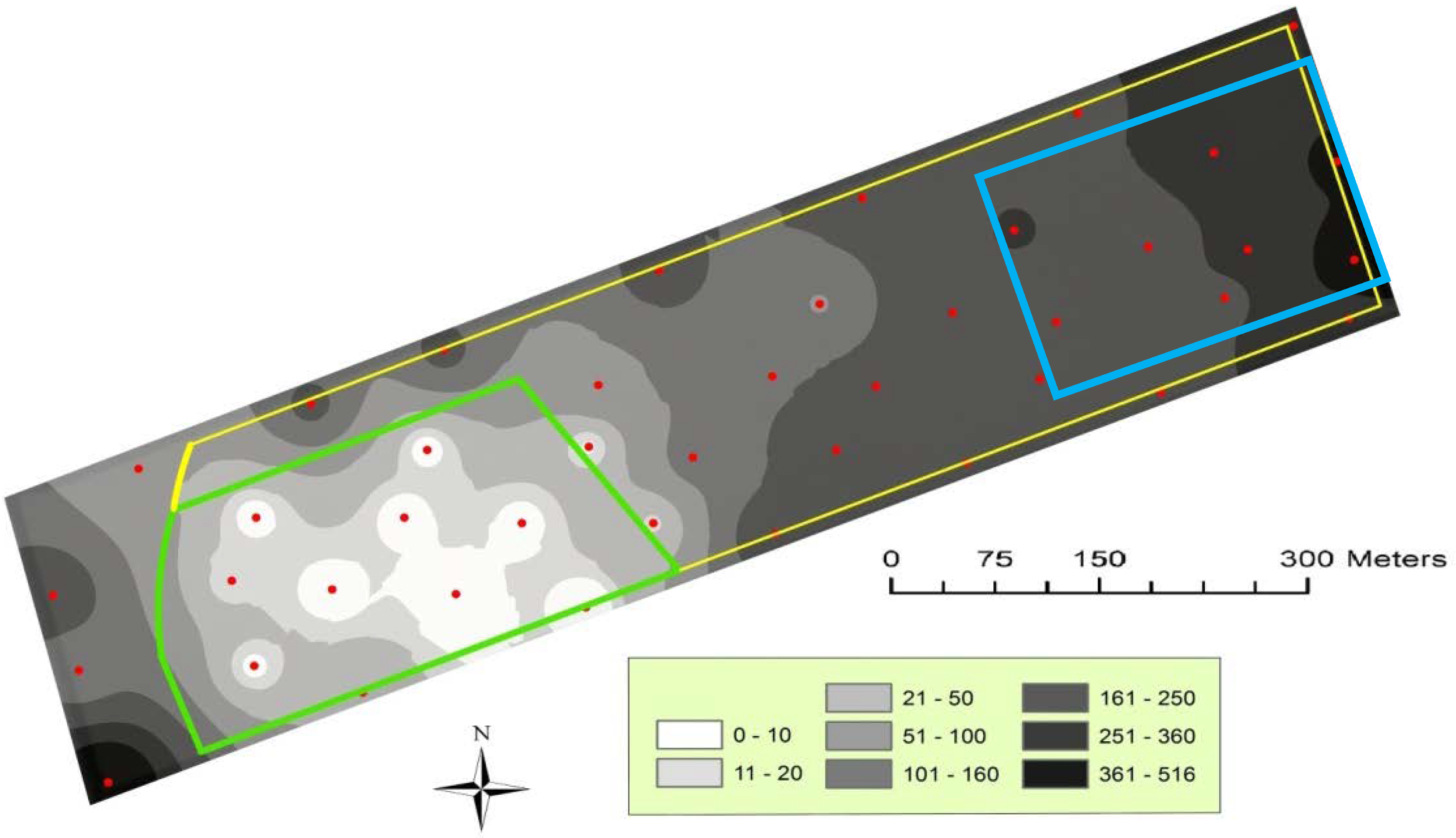
3.1. Mating Disruption Evaluation
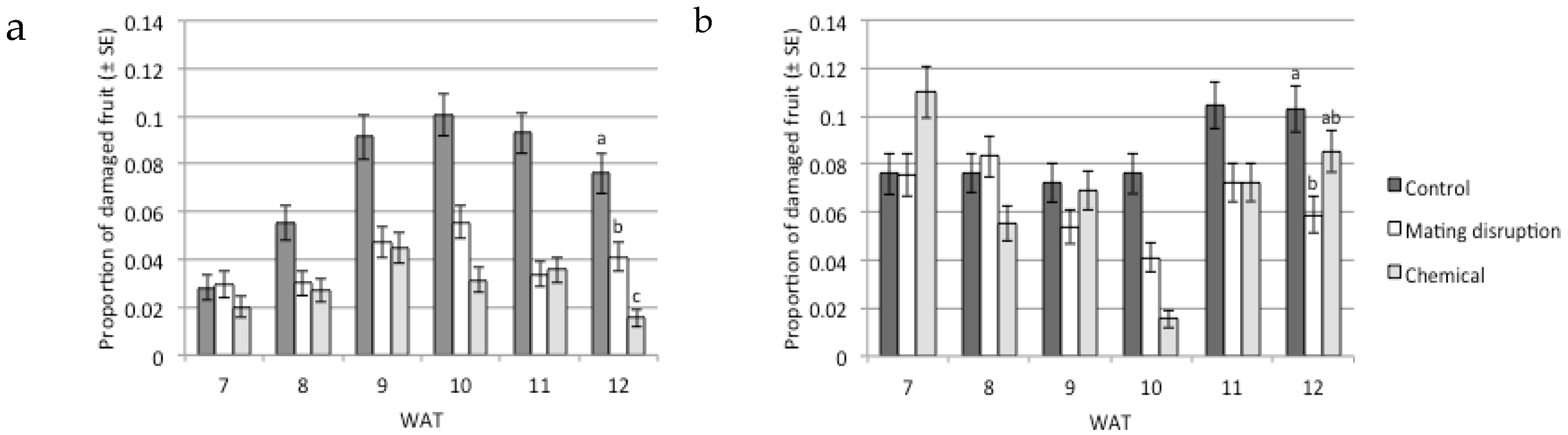
3.2. Fruit Damage Estimation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lammers, J.W.; Macleod, A. Report of a Pest Risk Analysis: Helicoverpa Armigera (Hübner, 1808). Available online: https://secure.fera.defra.gov.uk/phiw/riskRegister/downloadExternalPra.cfm?id=3879 (accessed on 2 July 2018).
- Cherry, A.; Cock, M.; van den Berg, H.; Kfir, R. Biological control of Helicoverpa armigera in Africa. In Biological Control in IPM Systems in Africa; Neuenschwander, P., Borgemeister, C., Langewald, J., Eds.; CAB International: Wallingford, UK, 2003; pp. 329–346. [Google Scholar]
- Sannino, L.; Espinosa, B. I Parassiti Animali Delle Solanacee; Edagricole: Milano, Italy, 2009; p. 290. [Google Scholar]
- EPPO European and Mediterranean Plant Protection Organization. Distribution Maps of Quarantine Pests. Helicoverpa armigera. Available online: https://gd.eppo.int/taxon/HELIAR/distribution (accessed on 2 July 2018).
- EPPO European and Mediterranean Plant Protection Organization. Datasheets on Quarantine Pests–Helicoverpa Armigera. Available online: https://gd.eppo.int/download/doc/132_datasheet_HELIAR.pdf (accessed on 2 July 2018).
- (Istat) Istituto Nazionale di Statistica. Census Database. 2018. Available online: http://dati-censimentoagricoltura.istat.it (accessed on 2 July 2018).
- Buès, R.; Bouvier, J.C.; Boudinhon, L. Insecticide resistance and mechanisms of resistance to selected strains of Helicoverpa armigera (Lepidoptera: Noctuidae) in the south of France. Crop Prot. 2005, 24, 814–820. [Google Scholar] [CrossRef]
- Mitchell, R.E. Pheromones: As the glamour and glitter fade: The real work begins. Fla. Entomol. 1986, 69, 132–139. [Google Scholar] [CrossRef]
- Birch, M.C.; Haynes, K.F. Insect Pheromones, 1st ed.; Studies in Biology No. 147; Hodder Arnold, H & S.: London, UK, 1982; p. 58. [Google Scholar]
- Kehat, M.; Dunkelblum, E.; Gothilf, S. Mating disruption of the cotton leafworm, Spodoptera littoralis (Lepidoptera: Noctuidae), by release of sex pheromone from widely separated Hercon-laminated dispensers. Environ. Entomol. 1983, 12, 1265–1269. [Google Scholar] [CrossRef]
- De Souza, K.; McVeigh, L.J.; Downham, M.C.A.; Smith, J.; Moawad, G.M. Mating-disruption of Spodoptera littoralis in Egypt. WPRS Bull. 1993, 16, 212. [Google Scholar]
- Mitchell, E.R.; Kehat, M.; McLaughin, J.R. Suppression of mating by beet armyworm (Noctuidae: Lepidoptera) in cotton with pheromone. J. Agri. Entomol. 1997, 14, 17–28. [Google Scholar]
- Wakamura, S.; Takai, M. Communication disruption for control of the beet armyworm, Spodoptera exigua (Hübner), with synthetic sex pheromone. Jpn. Agri. Res. Q. 1995, 29, 125. [Google Scholar]
- Kerns, D.L. Mating disruption of beet armyworm (Lepidoptera: Noctuidae) in vegetables by a synthetic pheromone. Crop Prot. 2000, 19, 327–334. [Google Scholar] [CrossRef]
- Rama, F.; Reggiori, F.; Albertini, A. Control of Spodoptera littoralis (Bsdv.) by biodegradable, low-dosage, slow-release pheromone dispensers. IOBC/Wprs Bull. 2011, 72, 59–66. [Google Scholar]
- Lanzoni, A.; Bazzocchi, G.G.; Reggiori, F.; Rama, F.; Sannino, L.; Maini, S.; Burgio, G. Spodoptera littoralis male capture suppression in processing spinach using two kinds of synthetic sex-pheromone dispensers. Bull. Insectol. 2012, 65, 311–318. [Google Scholar]
- Kehat, M.; Dunkelblum, E. Sex pheromones: Achievements in monitoring and mating disruption of cotton pests in Israel. Arch. Insect Biochem. Physiol. 1993, 22, 425–431. [Google Scholar] [CrossRef]
- Kehat, M.; Anshelevich, L.; Gordon, D.; Harel, M. Evaluation of Shin-Etsu twist-tie rope dispensers by the mating table technique for disrupting mating of the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae), and the pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelechiidae). Bull. Entomol. Res. 1989, 88, 141–148. [Google Scholar] [CrossRef]
- Chamberlain, D.J.; Brown, N.J.; Jones, O.T.; Casagrande, E. Field evaluation of a slow release pheromone formulation to control the American bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. Bull. Entomol. Res. 2000, 90, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex Pheromone Aerosol Devices for Mating Disruption: Challenges for a Brighter Future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, R.E.; Borth, P.W.; Tollefson, J.J. Stochastic simulation for characterizing ecological spatial patterns and appraising risk. Ecol. Appl. 1993, 3, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, A.; Trematerra, P. Geostatistical tools for the study of insect spatial distribution: Practical implications in the integrated management of orchard and vineyard pests. Plant Protect. Sci. 2014, 50, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Isaaks, E.H.; Srivastava, R.M. An Introduction to Applied Geostatistics; Oxford University Press: New York, NY, USA, 1989; p. 592. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5nd ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010; p. 944. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics. A Biometrical Approach, 3rd ed.; WCB McGraw-Hill: New York, NY, USA, 1997; p. 666. [Google Scholar]
- Sharpe, D. Your Chi-Square test is statistically significant: Now what? Pract. Assess. Res. Eval. 2015, 20, 1–10. [Google Scholar]
- Garcia Moral, F.J. Analysis of the spatio—Temporal distribution of Helicoverpa armigera Hb. in a tomato Field using a stochastic approach. Biosyst. Eng. 2011, 93, 253–259. [Google Scholar] [CrossRef]
- Ge, S.K.; Carruthers, R.I.; Ma, Z.F.; Zhang, G.X.; Li, D.M. Spatial heterogeneity and population risk analysis of cotton bollworm, Helicoverpa armigera, in China. Insect Sci. 2005, 12, 255–261. [Google Scholar] [CrossRef]
- Masetti, A.; Butturini, A.; Lanzoni, A.; De Luigi, V.; Burgio, G. Area-wide monitoring of potato tuberworm (Phthorimaea operculella) by pheromone trapping in Northern Italy: Phenology, spatial distribution and relationships between catches and tuber damage. Agric. For. Entomol. 2015, 17, 138–145. [Google Scholar] [CrossRef]
- De Luigi, V.; Furlan, L.; Palmieri, S.; Vettorazzo, M.; Zanini, G.; Edwards, C.R.; Burgio, G. Results of WCR monitoring plans and evaluation of an eradication program using GIS and Indicator Kriging. J. Appl. Entomol. 2011, 135, 38–46. [Google Scholar] [CrossRef]
- Dradi, D.; ASTRA Innovazione e Sviluppo, Cesena, Italy. Personal communication, 2012.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Day | Commercial Name | Active Ingredient | Dose kg c.p./ha |
|---|---|---|---|---|
| 2011 | 04 July | Steward® | Indoxacarb (30 g/L) | 0.125 |
| 21 July | Affirm® | Emamectin Benzoate (0.95%) | 1.5 | |
| 2012 | 19 July | Affirm® | Emamectin Benzoate (0.95%) | 1.5 |
| 10 August | Steward® | Indoxacarb (30 g/L) | 0.125 |
| Year | Weeks after Dispenser Position | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | Total | |
| 2011 | NA | NA | NA | - | 100 | 100 | 100 | 100 | 100 | 100 | 98.8 | 98.8 | 99.1 |
| 2012 | 100 | 98.5 | 100 | 100 | 98.9 | 100 | 98.8 | 98.6 | 99.3 | 96.7 | NA 1 | NA 1 | 98.4 |
| Year | Map | R | p | Mean Prediction Error |
|---|---|---|---|---|
| 2011 | July, 21 | 0.80 | <0.001 | 0.01 |
| July, 28 | 0.75 | <0.001 | −0.74 | |
| August, 4 | 0.24 | >0.05 | −0.01 | |
| August, 11 | 0.45 | <0.05 | −0.44 | |
| August, 16 | 0.45 | <0.05 | 0.34 | |
| Total catches | 0.71 | <0.001 | −0.91 | |
| 2012 | August, 1 | 0.42 | <0.05 | −0.62 |
| August, 8 | 0.81 | <0.001 | −0.99 | |
| August, 16 | 0.69 | <0.001 | −2.88 | |
| August, 23 | 0.57 | <0.001 | −1.79 | |
| August, 29 | 0.70 | <0.001 | −3.67 | |
| Total catches | 0.73 | <0.001 | −13.6 |
| Effect | χ2 | d.f. | p |
|---|---|---|---|
| Treatments * Damage | 103.2 | 2 | <0.01 |
| Years * Damage | 80.1 | 1 | <0.01 |
| Date * Damage | 77.3 | 5 | <0.01 |
| Treatments * Years * Damage | 66.7 | 2 | <0.01 |
| Treatments * Date * Damage | 48.5 | 10 | <0.01 |
| Years * Date * Damage | 39.7 | 5 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgio, G.; Ravaglia, F.; Maini, S.; Bazzocchi, G.G.; Masetti, A.; Lanzoni, A. Mating Disruption of Helicoverpa armigera (Lepidoptera: Noctuidae) on Processing Tomato: First Applications in Northern Italy. Insects 2020, 11, 206. https://doi.org/10.3390/insects11040206
Burgio G, Ravaglia F, Maini S, Bazzocchi GG, Masetti A, Lanzoni A. Mating Disruption of Helicoverpa armigera (Lepidoptera: Noctuidae) on Processing Tomato: First Applications in Northern Italy. Insects. 2020; 11(4):206. https://doi.org/10.3390/insects11040206
Chicago/Turabian StyleBurgio, Giovanni, Fabio Ravaglia, Stefano Maini, Giovanni Giorgio Bazzocchi, Antonio Masetti, and Alberto Lanzoni. 2020. "Mating Disruption of Helicoverpa armigera (Lepidoptera: Noctuidae) on Processing Tomato: First Applications in Northern Italy" Insects 11, no. 4: 206. https://doi.org/10.3390/insects11040206