Effects of Exogenous Spermidine on Root Metabolism of Cucumber Seedlings under Salt Stress by GC-MS

1
Collaborative Innovation Center for Improving Quality and Increase Profits of Protected Vegetables in Shanxi, College of Horticulture, Shanxi Agricultural University, Taigu 030801, China
2
Collaborative Innovation Center of Sustainable Forestry in Southern China, College of Forestry, Nanjing Forestry University, Nanjing 210037, China
3
Department of Public Order, Nanjing Forest Police College, Nanjing 210023, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2020, 10(4), 459; https://doi.org/10.3390/agronomy10040459
Submission received: 20 February 2020
/
Revised: 21 March 2020
/
Accepted: 23 March 2020
/
Published: 26 March 2020
(This article belongs to the Special Issue Effects of Abiotic Stresses and Their Control on Quality of Horticultural Products)
Abstract
:To investigate the effects of exogenous spermidine (Spd) on metabolism changes under salt stress in cucumber roots, a gas chromatography-mass spectrometry (GC-MS) was performed. The results showed that most of the 142 metabolites responded to salt stress or exogenous Spd treatment. Salt stress reduced carbon consumption, resulted in the transformation of glycolysis and the tricarboxylic acid (TCA) cycle to the pentose phosphate pathway (PPP), and meanwhile increased salicylic acid (SA) and ethylene synthesis, and, thus, inhibited the growth of seedlings. However, exogenous Spd further improved the utilization of carbon, the energy-saving pattern of amino acid accumulation, and the control of hydroxyl radicals. In conclusion, Spd could promote energy metabolism and inhibit SA and ethylene synthesis in favor of root growth that contributes to higher salt tolerance. This study provides insight that may facilitate a better understanding of the salt resistance by Spd in cucumber seedlings.
1. Introduction
Agricultural production has been seriously affected by soil salinity because the salt level has been a major limiting factor for plant growth, and affects a large terrestrial area in the world [1]. At present, nearly 20% of the irrigated land in the world is affected by salt, and more than 50% of the cultivated land is expected to be seriously salinized by 2050 [2,3]. High salt concentration in soil often causes osmotic stress (e.g., water deficit and nutrient imbalance), ion toxicity, and secondary stress (e.g., oxidative stress), leading to “physiological drought”, cellular damage, and growth reduction, and even death of plants [4,5]. Accordingly, a number of pathways are affected by salt stress, including energy metabolism, secondary metabolism, phytohormone synthesis, and signal transduction. Plants have also developed many biochemical and molecular mechanisms for the detrimental effects of salt stress through activation of cascades of molecular networks involved in signal transduction, the utilization of carbon, and the synthesis of compatible solutes and the control of hydroxyl radicals [4,6]. To cope with high salt concentrations, new technologies (e.g., proteomics and metabolomics) and exogenous growth regulators (e.g., polyamines (PAs) and γ-aminobutyric acid (GABA)) have been used [7,8].
In recent years, following genomics, transcriptomics, and proteomics, metabolomics as an original tool to obtain a comprehensive perspective of the mechanisms have been widely and rapidly used in salt stress studies [2,7]. Metabolites can be considered as the end products of gene expression, and represent the amount of all the levels of regulation in between gene and enzyme [2,9]. Correspondingly, linking metabolomics changes in the biochemical pathway with the enzymes involved can effectively assist in the identification of gene functions. Therefore, metabolomics can be a powerful method for studying the principle of plant response to salt stress. When plants are damaged by salt stress, metabolomics studies are performed to expound variations in metabolic pathways and networks, and to look for key genes in plant metabolic pathways [9]—then applying exogenous materials and other means to analyze and evaluate the metabolites before and after treatment, providing a basis for improving plant resistance and analyzing gene functions. However, this is a challenging task due to the known number of metabolites in plants being estimated to exceed 200.000, and the massive molecules with different structures and chemical properties [2,10]. Accordingly, many targeted techniques and different analytical approaches have been designed and used for a specific genotype or sample, allowing us to make insight into the multiple physiological processes in responses to stress conditions [7,11].
Polyamines (PAs) are aliphatic nitrogenous groups with low molecular mass commonly found in plants [8]. In general, common natural PAs include the higher PAs, spermine (Spm) and spermidine (Spd), and their diamine obligate precursor putrescine (Put) [12]. In plants, PAs play an important role in response to abiotic stress due to their polycationic nature [13]. For instance, PAs can protect negatively charged macromolecules, such as DNA, RNA, and certain proteins [14]. Moreover, recent studies have shown that PAs have diverse functions and roles in response to plant stress [15,16]. For instance, PAs can act as direct hydroxyl radical scavenging and influence antioxidant activity at molecular and gene expression level. In addition, PAs participate in signal transductions and form a complex network with nitric oxide, abscisic acid, ethylene, and Ca2+ in response to stress conditions [14,16]. According to this complex network from the cell to the entire plant, changes in their metabolism may cause a shift in a series of cellular metabolism pathways, such as energy metabolism and plant hormone synthesis [15]. Furthermore, compared to other plant hormones, PAs are often present in high concentrations. Hence, high PAs concentrations have been linked to salt stress tolerance in many plant species [16]. As a result, exogenous application of PAs has been widely used as a strategy to enhance salinity tolerance. Among the three major PAs, there is significant evidence suggesting that exogenous application of Spd could preserve the integrity of plant cell membrane, maintain cellular ionic balance minimize growth inhibition caused by salt stress, and reduce superoxide radical contents [8,13].
Cucumber (Cucumis sativus L.) is one of the most important vegetable crops worldwide, but it is highly sensitive to salinity because its roots are fragile, aerobic, and shallowly distributed, especially in the germination and early growth phases [17]. Serious secondary salinization in greenhouse soils causes large-scale physiological diseases of cucumber, reduces the yield and quality of cucumber, and leads to huge losses of economic value [18]. Recent studies in cucumber have shown that exogenous application of Spd could enhance seedlings stress tolerance [16,19]. Despite many studies on the functions and the roles of exogenous Spd in salt stress resistance, it remains unclear how exogenous Spd regulates cucumber growth and molecular responses. Therefore, our research used salt-sensitive cucumber cultivar ‘Jinchun No.2′ as material, and combined with the gas chromatography-mass spectrometry (GC-MS) technology to investigate metabolites change in cucumber roots. GC-MS is a powerful technique for the identification and quantification of metabolites, especially for analysis of distinctive tolerance mechanisms in different plant organs of horticultural crops under salt stress [14]. We identified specific metabolites whose abundances changed under salt stress with exogenous Spd using comparative metabolomics analysis. Combined with an analysis of important growth parameters, we revealed a close relationship between the differentially expressed metabolites and the observed physiological changes. Consequently, the metabolic mechanism of exogenous Spd responses to alleviate salt stress in cucumber seedling roots was proposed.
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
Cucumber (cv. Jinchun No. 2) seeds were soaked in hot water at the temperature of 55 °C for 15 min, then were transferred onto moist filter papers in the germination boxes in the dark at the temperature of 28 °C for 30 h. The germinated seedlings were sown in plastic trays (41 cm × 41 cm × 5 cm) containing quartz sand, and were cultivated in a greenhouse under natural light with a relative humidity of 60–75%, and a temperature of 26 ± 2 °C/day and 16 ± 2 °C/night at the Shanxi Agricultural University, China. When the cotyledons had expanded, seedlings were supplied with water containing a concentration of Hoagland’s nutrient solution every two days. After the three pieces of the euphylla were fully developed, 48 seedlings with uniform size were transplanted into plastic containers (51 cm × 33 cm × 20 cm) with aerated Hoagland’s nutrient solution. The hydroponic solutions were kept at the temperature of 22 ± 2 °C and were continuously aerated using an air pump at an interval of 30 min to maintain the dissolved oxygen at 8.0 ± 0.2 mg/L, and were renewed every 3 days.
After three days of pre-culture, the cucumber seedlings were subjected to three different treatments: (1) CK, control seedlings were grown in Hoagland’s solution; (2) S, seedlings were grown in Hoagland’s solution containing 75 mM NaCl; (3) SS, seedlings were grown in Hoagland’s solution containing 75 mM NaCl and 0.1 mM Spd. Each treatment had four replicates, and a total of 16 plants per treatment. After five days of treatment, five seedlings were collected for the determination of physiological parameters, and eight roots of seedlings per treatment were collected and frozen immediately in liquid nitrogen and stored at −80 °C to study the characteristic metabolites [10].
2.2. Determination of Plant Growth
The height (The distance between the cotyledon node to growing point) and stem diameter (below the cotyledon node 1 cm) of the seedlings were determined with a ruler and vernier caliper, respectively [20]. After cucumber seedlings were washed with sterile distilled water and the water wiped off with tissue paper, the maximum length, maximum width, and total area of the euphylla, and total length, surface area, volume, average diameter, and number of roots were measured using an Expression 1680 scanner (Epson, Sydney, Australia) and analyzed with WinRHIZO (Regent Instruments Ltd., Ontario, Canada). After fresh weights of shoots and roots were measured, shoots and roots were transplanted into a forced air oven at 105 °C for 15 min, and then placed at 75 °C for 3 days to determine their dry weights.
2.3. Metabolite Extraction
The exact mass of the root of the cucumber seedling sample (60 mg of fresh weight) was weighed and transferred into 1.5 mL centrifuge tubes. Then 360 μL of cold methanol and 40 μL of L-2-chloro-phenylalanine (0.3 mg/mL stock in methanol) were added and put into a Tissuelyser-48 (60 Hz) to grind into tissue homogenate for 2 min. After the tissue homogenate was extracted with an ultrasonic wave for 30 min, followed by adding 200 μL chloroform and 400 μL deionized water, it was then extracted with an ultrasonic wave for 30 min and centrifuged for 10 min at 14,000 rpm. Seven hundred microliters of supernatant was transferred into a glass centrifuge tube. After samples were rapidly volatile-dried by centrifuge enrichment, 80 μL methoxylamine pyridine solution (15 mg/mL) was added, vortexed for 2 min, and incubated for 90 min at 37 °C. Each sample was obtained by adding 80 µL bis(trimethylsilyl)trifluoroacetamide (BSTFA) derivatization reagent (containing 1% trimethylchlorosilane (TMCS)) and 20 µL n-Hexane, vortexed for 2 min, and incubated for 40 min at 40 °C. Finally, samples were placed for 30 min at room temperature and waited for the GC-MS metabolism analysis.
2.4. GC-MS Analysis and QC
Each 1 μL aliquot of the derivatized solution was injected in splitless mode into the Agilent 7890A-5975C GC-MS system (Agilent, Palo Alto, CA, USA). Separation was carried out on a non-polar DB-5 capillary column (30 m × 250 μm I.D., J&W Scientific, Folsom, CA, USA), with high purity helium as the carrier gas at a constant flow rate of 1.0 mL/min. The GC temperature programming began at 60 °C, followed by 8 °C/min oven temperature ramps to 125 °C, 4 °C/min to 210 °C, 5 °C/min to 270 °C, and 10 °C/min to 305 °C, and a final 3 min maintenance at 305 °C. The electron impact (EI) ion source was held at 260 °C with a filament bias of −70 V. Full scan mode (m/z 50-600) was used, with an acquisition rate of 20 spectrum/s in the MS setting. The quaility control (QC) sample was prepared by mixing aliquots of all samples to obtain a pooled sample and then analyzed using the same method as the analytic samples. The QCs were injected at regular intervals (every eight samples) throughout the analytical run to provide a set of data from which repeatability can be assessed.
2.5. Statistical Analysis
The acquired MS data from GC-MS were analyzed by ChromaTOF software (v 4.34, LECO, St Joseph, MI, USA). Briefly, after alignment with the Statistic Compare component, the CSV file was obtained with three-dimension data sets, including sample information, retention time-m/z, and peak intensities. The detectable peaks of urine samples in GC-MS were 716 in total, and the internal standard was used for data quality control (reproducibility). After internal standards and any known pseudo positive peaks, such as peaks caused by noise, column bleed, and the BSTFA derivatization procedure, were removed from the data set, and the peaks from the same metabolite were combined, the detectable metabolites of urine samples in GC-MS were 142 in total. The data set was normalized using the sum intensity of the peaks in each sample.
The data sets resulting from GC-MS were separately imported into the SIMCA-P+ 14.0 software package (Umetrics, Umeå, Sweden). Principle component analysis (PCA) and (orthogonal) partial least-squares-discriminant analysis (O)PLS-DA were carried out to visualize the metabolic alterations among experimental groups, after mean centering and unit variance scaling. Variable importance in the projection (VIP) ranks the overall contribution of each variable to the (O)PLS-DA model, and those variables with VIP > 1.0 are considered relevant for group discrimination.
In this study, the default 7-round cross-validation was applied with 1/seventh of the samples being excluded from the mathematical model in each round, to guard against overfitting. Significant differences of metabolites between treatment were tested using one-sample or independent sample Student’s t-test analysis on SPSS19 software, and the growth indices data were statistically analyzed using Duncan’s new multiple range test at the 0.05 level of significance.
3. Results
3.1. Growth Parameters of Cucumber Seedlings
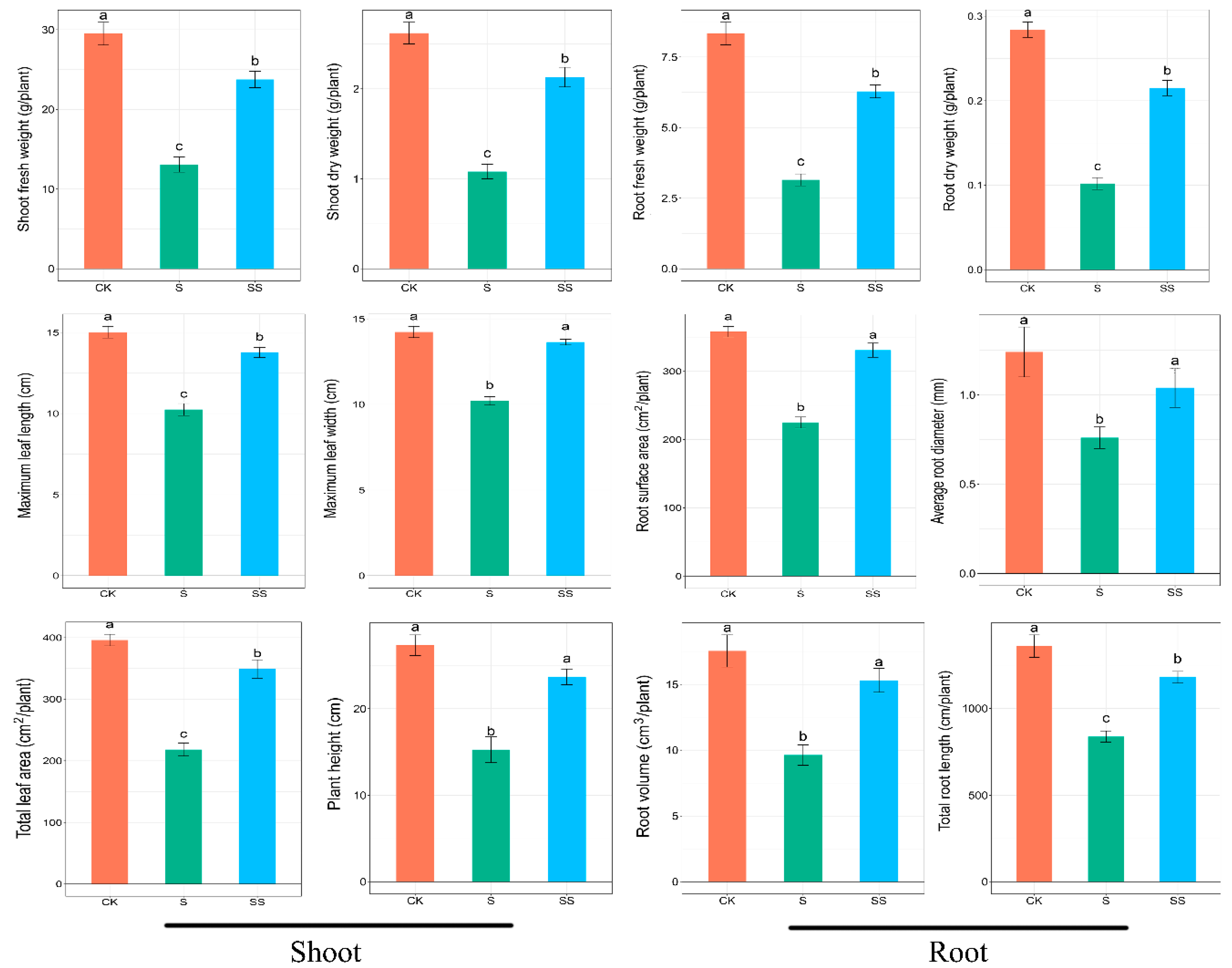
After five days of treatment, the salt stress significantly decreased plant growth, while exogenous Spd alleviated the growth inhibition by salinity (Figure 1). Compared with the control, the shoot fresh weight, shoot dry weight, maximum leaf length, maximum leaf width, total leaf area, and plant height of salt treatment were reduced by 55.7%, 63.7%, 31.8%, 28.2%, 45.1%, 44.2%, respectively. The shoot fresh weight, shoot dry weight, maximum leaf length, maximum leaf width, total leaf area, and plant height of Spd-treated were increased by 81.7%, 113.33%, 34.2%, 33.6%, 60.1%, 55.2%, respectively, compared to salt stress (Figure 2). In addition, the fresh weight, dry weight, surface area, average diameter, volume, and total length of roots were significantly suppressed by salt stress, while these indicators were significantly increased by exogenous Spd under salt stress. However, the surface area, average diameter, and volume of roots in Spd-treated did not significantly change as compared with the control (Figure 2). Thus, these results clearly showed that salt stress inhibited the growth of cucumber seedlings, and exogenous Spd could alleviate these damages.
3.2. Metabolic Changes in Cucumber Seedlings
To reveal the physiological mechanism of cucumber seedlings by Spd under the salt stress, quantitative and qualitative root metabolites analysis of 24 samples were carried out. The total ion chromatogram (TIC) of 24 samples is shown in Figure S1. An obvious chromatographic difference between sample groups revealed that the retention time was reproducible and stable, implying that the metabolomic analysis was reliable. The metabolite profiles showed a significant difference in all treatments. A total of 142 kinds of metabolites were identified, and their relative concentrations were determined (Table 1).
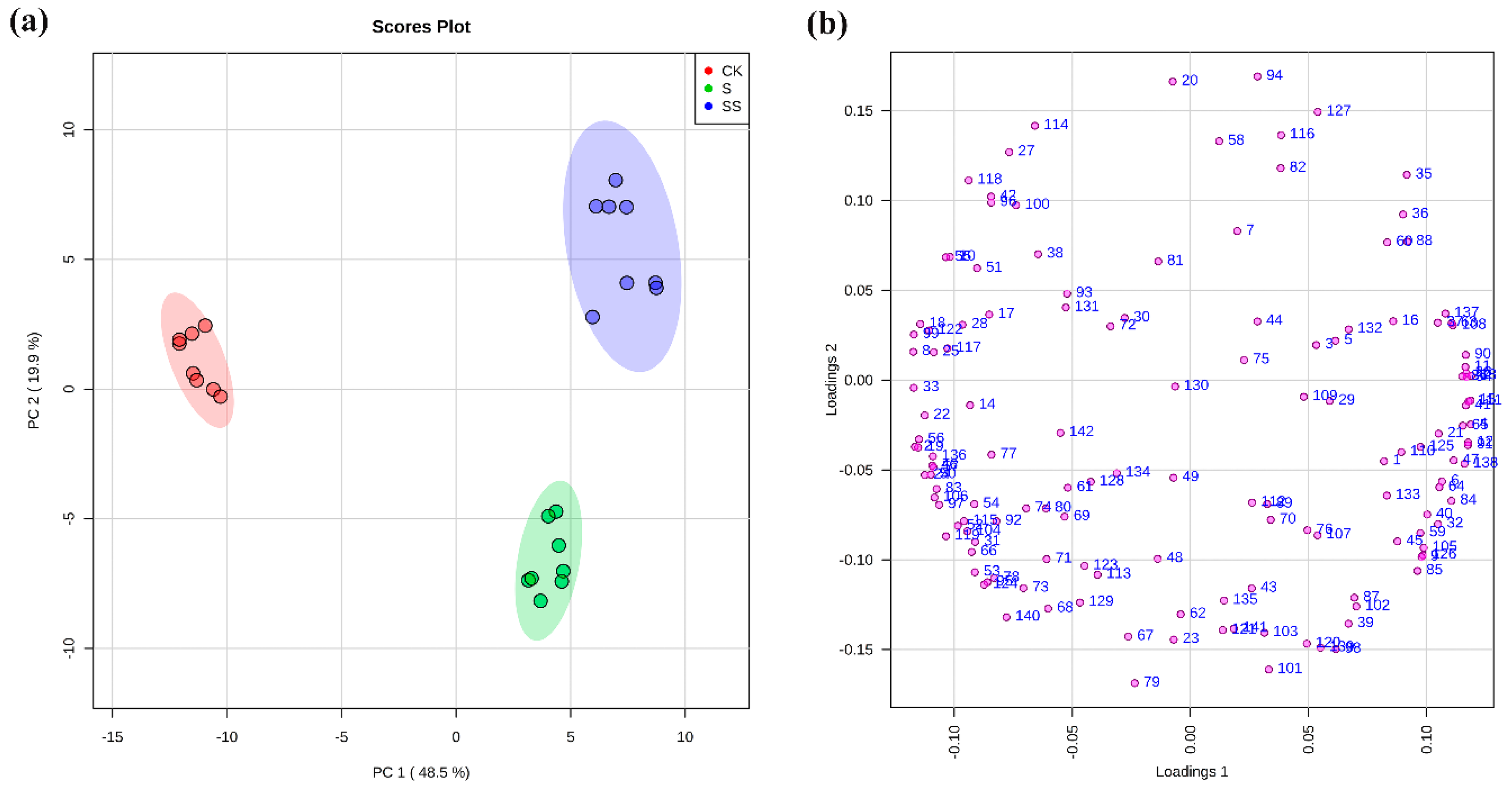
Based on the PCA and Heatmap analysis results (Figure 3a and Figure S2), a separation of samples under control, salt stress with or without Spd in roots was observed. The samples of different treatments were separated by the first principal component (PC1) and the second principal component (PC2), representing 48.5% and 19.9% of the total variation (Figure 3a), respectively. Each number in the loading plot (Figure 3b) represented a variable. Metabolites contributing to the separation between different treatments could be obviously identified due to the most of the variables that were away from the coordinate center. The samples of control (CK) and salt stress with or without Spd (S and SS) were clearly separated by PC1, whereas PC2 distinguished the samples of presence (SS) and absence (S) of Spd under salt stress. The contribution of metabolites in roots of cucumber seedlings for PC1 came from a number of metabolites with the most discrimination power dominated by sugars and their intermediate metabolites (e.g., xylose, succinic acid, 3-PGA, galactinol, erythrose-4-phosphate, ribulose-5-phosphate glucose-6-phosphate, and trehalose-6-phosphate), and amino acids (e.g., glycine, citrulline, and asparagine), while organic acids (e.g., azelaic acid, aminooxyacetic acid (AOAA), and glucoheptonic acid), some amino acids (e.g., valine, cysteine, and threonine), nucleotide metabolic pathway (e.g., hypoxanthine, thymine, and uridine) and polyamines (e.g., spermidine) were the dominate metabolites contributing to PC2 (Figure 3b and Table S1).
3.3. Differentially Metabolites under Salt Stress with or without Spd
To investigate the metabolic differences by salt stress and exogenous Spd, we compared the content of metabolites between the control and salt stress with or without Spd treatment (Table 1, Figure 4).
3.3.1. Sugar Metabolic Changes
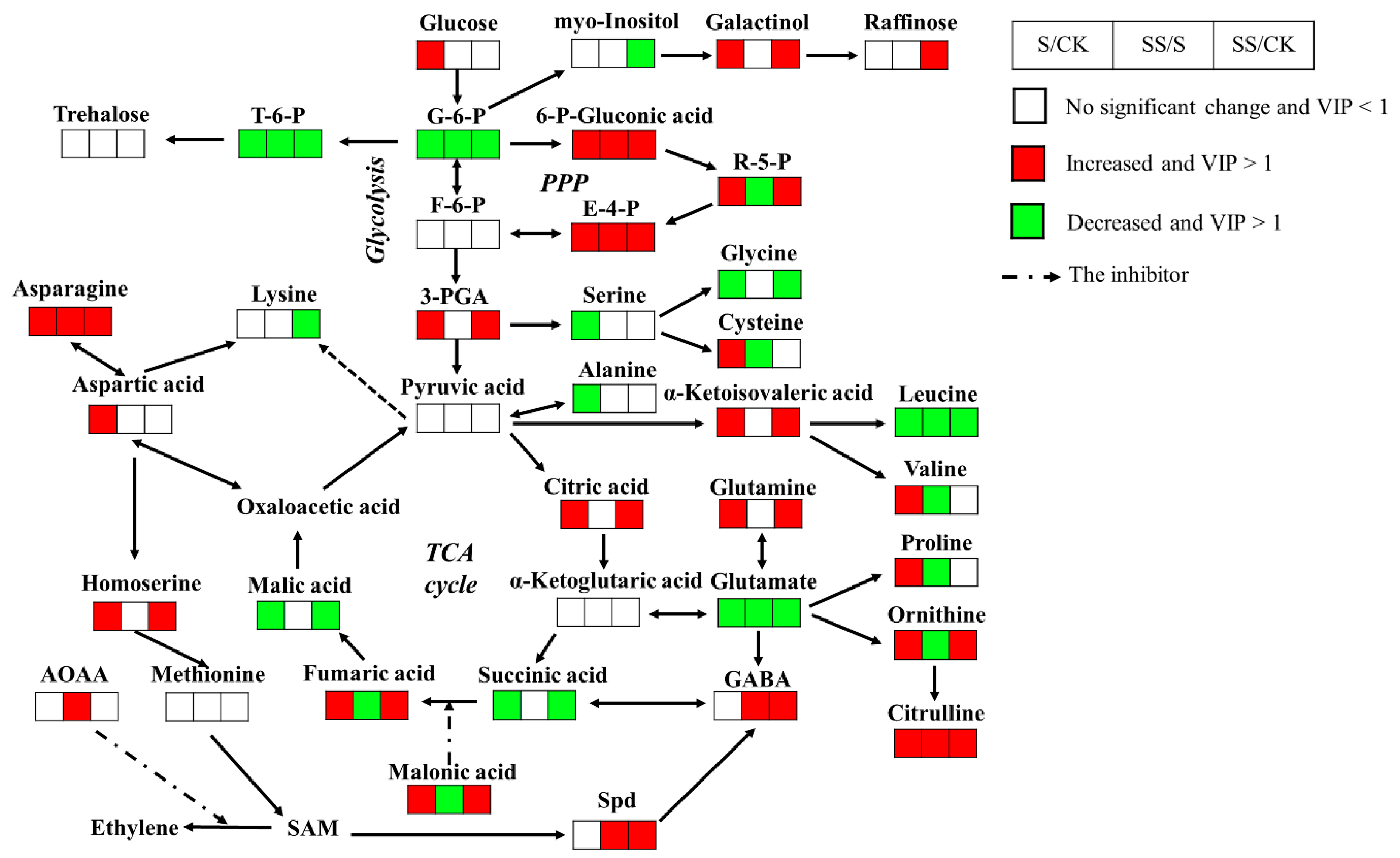
A large number of detected metabolites involved in primary metabolic pathways were found under salt stress. Compared with the control, salt stress increased the levels of 3-Phosphoglycerate(3-PGA), citric acid, and fumaric acid, that are involved in the glycolytic pathway and the tricarboxylic acid (TCA) cycle, and the levels of 6-phosphogluconic acid, ribulose-5-phosphate, and erythrose-4-phosphate, that are involved in the pentose phosphate pathway (PPP) (p < 0.001 and VIP > 1; Figure 4). Meanwhile, salt stress significantly decreased the levels of glucose-6-phosphate, succinic acid, and malic acid, which participated in the glycolysis and TCA cycle (p < 0.001 and VIP > 1; Figure 4). With exogenous Spd, the levels of glucose-6-phosphate, fumaric acid, and ribulose-5-phosphate were significantly decreased compared with salt stress, whereas the amount of 6-phosphogluconic acid and erythrose-4-phosphate were increased (p < 0.01 and VIP > 1; Figure 4). Although cucumber is a raffinose family oligosaccharides (RFOs) translocation plant, the level of raffinose was below the detection limit of GC-MS. With exogenous Spd, the levels of raffinose and galactinol were detected higher than the control, while myo-inositol was significantly decreased (p < 0.001 and VIP > 1; Figure 4). In addition, only galactinol was significantly increased without Spd under salt stress compared with the control (p < 0.001 and VIP > 1; Figure 4).
Most of the levels of the soluble sugars in roots were decreased under salt stress compared with the control. Only three soluble sugars, galactose, altrose, and levoglucosan, showed significantly increased levels after salt treatment (p < 0.01 and VIP > 1; Table 1). In contrast, the levels of sugar derivatives showed an opposite accumulation trend to soluble sugars after salt treatment. Sugar phosphate esters (e.g., glucose-1-phosphate), some sugar acids (e.g., saccharic acid, galactonic acid, and threonic acid), and sugar alcohols (e.g., threitol) were significantly increased under salt stress compared with the control (p < 0.001 and VIP > 1; Table 1). Only the level of trehalose-6-phosphate (T-6-P) decreased dramatically in both the presence and absence of Spd under salt stress as did ribitol (p < 0.001 and VIP > 1; Figure 4). Exogenous Spd had a certain impact on sugars and their derivatives compared with salt stress, such as erythrose, altrose, maltotriose, glucuronic acid, gluconic acid, and lactobionic acid were significantly increased; meanwhile, xylose, sedoheptulose, glucoheptose, talose, melibiose, T-6-P, lactobionic acid. 2-deoxy-D-galactose, and ribitol were significantly decreased (p < 0.001 and VIP > 1; Table 1, Figure 4).
3.3.2. Organic Acids Changes
High salinity caused a significant reduction in carboxylic acids content, including maleic acid, glyceric acid, 3-hydroxypropionic acid, 3-hydroxybutyric acid, and 2-hydroxybutanoic acid (p < 0.001 and VIP > 1; Table 1). In contrast, α-ketoisovaleric acid and 2-ketobutyric acid, which were confirmed in the amino acid metabolism pathway, and malonic acid, an inhibitor of the amber deoxy enzyme, were dramatically enhanced in salt stress (p < 0.001 and VIP > 1; Table 1). Meanwhile, compared with salt stress, exogenous Spd significantly improved the contents of some carboxylic acids (e.g., 3-hydroxypropionic acid, AOAA, benzoic acid, and pipecolinic acid), polyphenols (e.g., glycerol), and simple lipids (e.g., 1-monopalmitin and 2-monopalmitin) (p < 0.01 and VIP > 1; Table 1, Figure 4). By contrast, the contents of some carboxylic acids (e.g., maleic acid, glycolic acid, 2-furoic acid, lactic acid, 3-hydroxybutyric acid, 2-hydroxybutanoic acid, malonic acid, and pyrrole-2-carboxylic acid), polyphenols (e.g., gallic acid, pyrogallol, and 1,2,4-benzenetriol), alcohols (cuminic alcohol and dodecanol), and fatty acids (e.g., azelaic acid and heptadecanoic acid) were significantly downgraded by Spd under salt stress (p < 0.01 and VIP > 1; Table 1, Figure 4).
3.3.3. Amino Acids Changes
The results showed that most of the glutamate family (glutamine, ornithine, and citrulline) were upregulated; only glutamate was downregulated when cucumber seedlings were exposed to salt stress (p < 0.001 and VIP > 1; Figure 4). Among the aspartate family, asparagine and homoserine were significantly increased in roots of cucumber seedlings under salt stress (p < 0.001 and VIP > 1; Figure 4). Moreover, the levels of leucine, glycine, and α-aminoadipic acid were significantly decreased when cucumber seedlings were exposed to high salinity, whereas oxoproline, N-carbamylglutamate (NCG), cysteinylglycine, and maleimide were significantly increased (p < 0.01 and VIP > 1; Table 1, Figure 4). Furthermore, compared with the control, the levels of proline, aspartic acid, valine, and cysteine showed accumulation trends under salt stress, while alanine, threonine, and serine were significantly reduced (p < 0.001 and VIP > 1; Table 1, Figure 4). Compared with salt stress, exogenous Spd caused a significant increase in the levels of citrulline, GABA, asparagine, threonine, and NCG. By contrast, with exogenous Spd, the levels of glutamate, ornithine, proline, soleucine, leucine, valine, cysteine, aminoadipic acid, and aromatic amino acids were significantly decreased (p < 0.001 and VIP > 1; Table 1, Figure 4).
3.3.4. Other Metabolites Changes
A majority of the contents of the nucleotide metabolic pathway in roots, such as adenine, guanosine, adenosine, inosine, and β-alanine, were decreased under salt stress, while only allantoic acid and urea were significantly increased (p < 0.01 and VIP > 1; Table 1). By contrast, catecholamine metabolism pathway (e.g., noradrenaline) and steroids (e.g., 22-ketocholesterol) were significantly increased under salt stress, whereas dehydroascorbic acid was reduced (p < 0.001 and VIP > 1; Table 1). Meanwhile, exogenous Spd significantly improved the contents of thymine, uridine, and adenosine, and significantly reduced the contents of uracil, adenine, inosine, hypoxanthine, dihydrouracil, and urea, respectively. By contrast, compared with salt stress, the contents of Spd, plant hormones (e.g., salicin), and noradrenaline were significantly increased, while the contents of L-dopa, dehydroascorbic acid, 22-ketocholesterol, 21-hydroxypregnenolone, and nicotinic acid were significantly downgraded by exogenous Spd under salt stress (p < 0.01 and VIP > 1; Table 1).
4. Discussion
Plant roots are the first parts of plants that encounter soil stress and respond to stress signals through different physiological or morphological changes, so the growth status of the root system should be paid more attention [21]. All indicators of the root system of cucumber seedlings with Spd were significantly higher than salt stress. The total length, surface area, average diameter, and volume of roots of cucumber seedlings with Spd even had no significant difference compared with the control. These results clearly suggested that the cucumber seedlings with Spd attributed to significant improvement in growth status under high salinity. The findings were consistent with existing research results [16,19,22].
Plant growth under salt stress was closely related to carbohydrate accumulation and energy metabolism. The studies have shown that changes in the contents of some sugars and their derivatives are a normal response of a plant to salt stress [23,24,25]. In the root, sugars were derived from gluconeogenesis, sugar transport, and polysaccharide degradation [26]. As Na+ stress restricted the entry of CO2 from the environment into mesophyll cells, the photosynthesis rate and carbon assimilation capacity of cucumber seedlings were inhibited under salt stress [16,18]. Our result suggested that the accumulation of sugars and certain intermediates was not a simple active response to salt stress, but rather combined complex effects of glycolysis, TCA cycle, PPP, and gluconeogenesis. We also found that glycolysis and the TCA cycle, and the accumulation of sugar were inhibited, whereas PPP and raffinose family oligosaccharides (RFOs) were enhanced in roots under salt. Moreover, exogenous Spd had a certain impact on sugars and their derivatives compared with salt stress, enhancing the PPP and TCA cycle, increasing the content of some sugar acids, and inhibiting some sugar-phosphate esters.
Central metabolism (glycolysis and TCA cycle) not only valuably provided energy in stress conditions but also provided carbon skeletons for biosynthesis [27]. Correspondingly, the levels of carbon skeletons influenced central metabolism. Other studies have shown that the accumulation of citric acid and 3-PGA can inhibit glycolysis by inhibiting the activity of phosphofructokinase (PFK), which is a rate-limiting enzyme of glycolysis [28]. In this study, salt stress caused an increase in levels of citric acid and 3-PGA, especially the level of 3-PGA, which was below the detection limit of GC-MS in the control, which suggested that glycolysis was downregulated in response to salt stress. This conclusion was supported by the result showing that glycolysis in cucumber root tissues was inhibited by salt stress [29]. In other studies, most TCA cycle intermediates and organic acids in salt-sensitive plant roots showed a decreasing trend after salt stress [30,31,32]. However, the TCA cycle had duplicity or ammphibic of catabolism and anabolism, and intermediates were involved in a large number of biosynthetic pathways. Meanwhile, some degradation pathways can also produce TCA cycle intermediates, such as the oxidation of odd fatty acids, the deamination and transamination of amino acid, and the decomposition of valine to produce succinyl-CoA [33,34]. Therefore, it was not possible to simply determine whether the TCA cycle was inhibited or promoted by a decrease or increase in the concentration of certain intermediates. In this study, the increase in citric acid and fumaric acid, and the decrease in succinic acid and malic acid only indicated a perturbation of the TCA cycle under S and SS. However, salt stress could significantly increase the concentration of malonic acid, which is a representative strong inhibitor of the TCA cycle [35], whereas Spd-treated could significantly reduce its concentration. This meant that exogenous Spd can reduce the inhibition of the TCA cycle in plant root tissue by salt stress, thereby enhancing salt tolerance.
Other research has shown that the carbon pathway of glycolysis may flow into PPP to provide NADPH for antioxidant effects when plants were subjected to salt stress [36]. In this study, the level of ribulose-5-phosphate in roots under control conditions was below the detection limit of GC-MS. However, in the roots after the salt treatment, PPP intermediates, such as 6-phosphogluconic acid, ribulose-5-phosphate, and erythrose-4-phosphate, significantly increased in response to salt stress. The accumulation of PPP intermediates was in agreement with the report that the PPP-mediated recycling of NADPH is an important antioxidant molecule against salt-induced oxidative stress [37,38]. Furthermore, in the roots after the Spd treatment, the levels of ribulose-5-phosphate and glucose-6-phosphate were more significantly decreased than without Spd under salt stress, whereas the levels of 6-phosphogluconic acid and erythrose-4-phosphate were increased. The depletion of glucose-6-phosphate and the accumulation of 6-phosphogluconic acid indicated an increased enzyme activity of glucose 6-phosphate dehydrogenase (G6PDH), a key enzyme of PPP, and generated a large amount of NADPH [39]. This conclusion was supported by the result showing the increase in G6PDH transcripts was positively correlated with the accumulation of Spd [40]. As ribulose-5-phosphate is the link between the oxidative and non-oxidative branches of PPP, the depletion of ribulose-5-phosphate and the accumulation of erythrose-4-phosphate indicated an increased flow of carbons to the non-oxidative part of PPP, in which part a large amount of sugar metabolism intermediates are produced [41]. Thus, exogenous Spd could coordinate the plant’s demand for a large amount of sugar metabolism intermediates and NADPH in response to salt stress.
Carbohydrates are not only the primary material of energy for plant metabolism, but can also be involved in osmotic adjustment and hydroxyl radical scavenging to promote the cucumber salt-tolerance. In addition to sucrose transport plants, there were some RFOs that were the main forms of photosynthetic products, such as cucumber, which are commonly referred to as RFOs translocation plant [42,43]. Taji et al. reported that the contents of galactinol and raffinose in Arabidopsis thaliana with gene of TsGolS significantly increase compared with wild type, thereby enhanced the tolerance of plants to salt stress [44]. In this study, raffinose in roots could not be observed without salt treatment (less than the detection limit), possibly due to less relevance under normal growth conditions [44]. After salt stress, the level of galactinol in roots was significantly increased. It is well-known that galactinol and raffinose are regarded as osmolytes that protect cellular structures and hold osmotic balance under salt treatment [45]. Galactinol and raffinose have been reported to have a more superior hydroxyl radical scavenging capacity than typical antioxidants (e.g., ascorbate and glutathione) [46]. The accumulation of galactinol indicated an increased tolerance to saline ions and an improved ability for hydroxyl radical scavenging. The enzyme involved in the first step of RFO biosynthesis was galactitol synthase (GolS), which catalyzes the conversion of myo-inositol to galactitol. GolS is considered as a key regulator of the RFO biosynthetic pathway because the activity of this enzyme will determine the accumulation level of RFOs [46]. In this study, exogenous Spd further promoted the depletion of myo-inositol and the accumulation of galactitol, suggesting that Spd might induce RFOs synthesis by increasing GolS to enhance salt tolerance. However, the mechanism by which exogenous Spd promotes RFOs accumulation to maintain osmotic balance and hydroxyl radical scavenging remains to be further elucidated.
Although the accumulation of trehalose in the plant has been reported as one of the key protection mechanisms against salt stress conditions [5,23,25], a similar trend was not observed in this study. In fact, except for resuscitation plants and trehalose metabolic mutants, trehalose content was very low in the angiosperms; thus, a role as protectant or carbon storage to participate directly in stress protection seems less plausible [47]. On the other hand, many studies have shown that the trehalose precursor T-6-P was an important signal molecule that regulates plant metabolism and gene expression [48,49]. Recent studies have shown that T-6-P inhibits the activity of the SnRK1 (i.e., sucrose non-fermenting-1-related protein kinase 1), which are sensors of energy availability and regulate more than 1000 genes related to biosynthesis, growth metabolism, and stress response [50,51]. Under salt stress, SnRK1 activity increased as a consequence of decreased T-6-P, thereby activating catabolic energy-producing pathways and inhibiting anabolic energy-consuming pathways to maintain energy homeostasis [52,53]. The recent study has suggested that lower T-6-P played a wider role in regulating the source–sink relationship of plant metabolism under stress conditions, which can improve photosynthesis [54]. In our study, the lowest concentration of T-6-P was in Spd-treated roots. This result was supported by the study showing that Spd could improve the photosynthetic capacity of cucumber to increase salt tolerance [16,55].
Based on the above results, we suggested that exogenous Spd could regulate the central metabolism by more effective carbon utilization and improve the accumulation of carbon assimilation production, thereby providing more material and energy to promote the cucumber salt-tolerance. In addition, recent studies have shown that nitrogen metabolism is strongly interconnected with carbon metabolism [56]. Therefore, it is important to ensure that there are enough carbon skeletons and energy during the assimilation of nitrogen and the synthesis of amino acids for plant growth and development [57,58]. However, due to the inhibition of energy metabolism, plants respond to high salinity levels by inhibiting protein synthesis, promoting protein degradation, and changing amino acids composition [16,22,30].
Under salt stress, the synthesis of amino acids in plants mainly depended on the availability of energy and carbon skeletons. Glutamate and aspartic acid could act as an immediate donor of amino groups for the synthesis of most other amino acids and linked primary metabolisms, such as amino acid metabolism, nucleotide metabolism, and glucose metabolism, through their TCA cycle precursors [33,57,58]. Glutamine and asparagine had a higher nitrogen-to-carbon ratio and could sequester more nitrogen using the limited carbon skeletons in response to environmental stresses [58,59,60]. This conclusion supported the result of glutamine and asparagine accumulation under salt stress without Spd. In addition, the nitrogen transport of allantoic acid was energy efficient since the nitrogen-to-carbon ratio for allantoic acid in 1:1 compared with 2:5 for glutamine and 1:2 for asparagine [61]. Allantoic acid accumulation was associated with a number of traits involved in the tolerance of salt stress, through saving more carbon skeletons and salicylic acid (SA) signaling [54]. Based on those results, we suggested that the pattern of amino acid accumulation and nitrogen transport in cucumber under salt stress was closely related to the insufficient supply of photosynthetic products. Furthermore, asparagine accumulation and polyamine synthesis promoted each other in response to stress [62]. The further glutamate depletion and asparagine accumulation with Spd in this study suggested that Spd could further use the energy-saving pattern of amino acid accumulation and nitrogen transport in response to salt stress.
Meanwhile, glutamate could further synthesize citrulline, proline, ornithine, and GABA as active substances under salt stress [57,63,64], resulting in a further reduction in glutamate content in S and SS treatments. Citrulline was the most abundant in Cucurbitaceae plants and occurred as a primary carrier of organic nitrogen in the form of free amino acid [65,66]. Compared to mannitol, citrulline was identified as the most effective among compatible solutes in response to abiotic stress [67,68]. In this study, further exogenous Spd citrulline accumulation indicated that Spd-induced salt-tolerance was correlated with citrulline effectively protecting DNA and scavenging hydroxyl radicals.
Proline is generally considered to be an osmotic regulator, an active oxygen scavenger, and a protein structure in response to high salinity [30]. The major sources of proline in plants are the glutamate synthesis pathway and the ornithine synthesis pathway, both of which are activated to varying degrees under salt stress [64,69]. Activation of the ornithine synthesis pathway in seedlings played a vital role in improving plant salt tolerance [64]. This finding was consistent with our results that the levels of ornithine and proline without Spd under salt treatment were significantly higher than those of control. However, compared to without Spd, the level of proline was significantly decreased with Spd. One explanation for this observed discrepancy might be that the actual osmotic effect of proline was weak, and excessive accumulation of proline is related to the sensitivity of plant genotypes to salt stress [70,71]. This finding was consistent with the fact that our study used a salt-sensitive cucumber cultivar “Jinchun 2” as a material. In addition, NCG was an N-acetyl glutamate (NAG) analog, which has a wide range of biological functions in animals, such as participating in the urea cycle. The study has shown that NCG can be used as an arginine endogenous activator to promote the synthesis of arginine and citrulline by glutamine or proline and to stimulate nitric oxide (NO) and Spd to play positive effects in numerous metabolic cycles [72]. This might also be another reason for the further depletion glutamine and proline with exogenous Spd under salt stress. However, there are few reports on the role of NCG in plants, so the effect of exogenous Spd on NCG remains to be further elucidated.
Previous studies reported that GABA had a functional role in central carbon metabolism and membrane oxidation remediate in response to salt stress [73,74]. Recent research has shown that plant salt tolerance was closely related to promoting the accumulation of GABA and inhibiting the accumulation of glutamate [75]. The depletion of glutamate and the accumulation of GABA with Spd in this study suggested that Spd can effectively improve cucumber salt tolerance. This conclusion was supported by the result that Spd might be involved in GABA-remediated membrane oxidation injury under salinity stress [74]. Spd and GABA not only had similar functions but they could also promote to accumulate each other in response to stress. For example, GABA might promote the conversion of Put to Spd [76]. This might be due to plant salt tolerance being positively related to Spd levels but negatively related to Put levels [74]. This was verified in our study by the accumulation of Spd and depletion of Put in SS, although the decrease in Put was not obvious.
Our studies have highlighted the essential involvement of primary metabolites: sugars and their derivatives, glycolysis, PPP, and TCA cycle intermediates, and amino acids as direct markers of energy metabolism dysfunction as well as effectors of osmotic balance and hydroxyl radical scavenging. On the contrary, secondary metabolites were more specific of regulatory molecules and responded to salt stress conditions as antioxidants. It has been shown that the positive role of Spd was confirmed to ameliorate the salt stress effect by reducing SA content in cucumber [77]. This finding was in agreement with our result that the level of benzoic acid served as the precursor of SA was significantly increased to reduce the synthesis of SA in Spd treatment. On the other hand, plants converted excess free salicylic acid into bound salicylic acid (i.e., salicylic acid glucoside) to avoid damage [78]. It was consistent with the result that the level of salicin was significantly increased to reduce the toxicity to plant cells in Spd treatment. Numerous studies have shown that ethylene and PAs not only have opposite effects in regulating plant growth and development but also have antagonistic relationships [79]. In most cases, exogenous Spd could inhibit ethylene biosynthesis, which plays a role in reducing stress and delaying aging. Ethylene and Spd synthesis imposed competitive demands on the common precursor S-adenosylmethionine (SAM) [80], and Spd reduced ethylene synthesis by scavenging reactive oxygen species (ROS), which can promote the synthesis of ethylene [81]. Our results also found that exogenous Spd could significantly increase the concentration of AOAA served as the inhibitor of ethylene synthesis, thereby inhibiting ethylene synthesis. As ethylene mainly increased plant salt resistance by reducing growth [82], this might be one of the reasons why the growth indices of roots of cucumber seedlings with Spd were not significantly different from the control. Based on these findings, the Spd treatment might help alleviate the negative effect of some plant hormones on the growth of cucumber under salt stress.
5. Conclusions
In summary, the effect of salt stress and Spd treatment on cucumber seedlings was investigated at the metabolic level. Salt stress might lead to the transformation of glycolysis and the TCA cycle to PPP, and provide antioxidant effects for NADPH, and meanwhile reduce carbon consumption, regulate osmotic adjustment, increase SA and ethylene synthesis, and, thus, inhibit the growth of seedlings. While exogenous Spd further improved the utilization of carbon, the energy-saving pattern of amino acid accumulation and the control of hydroxyl radicals. Moreover, Spd could promote energy metabolism and inhibit SA and ethylene synthesis in favor of root growth that contributes to higher salt tolerance. This study provides insight that may facilitate a better understanding of the salt resistance by Spd in cucumber seedlings.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/10/4/459/s1, Figure S1: Total ion chromatogram (TIC) based on GC-MS for the cucumber seedlings root. Figure S2. Heatmap analysis combined with hierarchical cluster analysis (HCA) of the top 60 metabolites. Table S1: The contribution of metabolites in roots to the first principal component (PC1) and the second principal component (PC2).
Author Contributions
Conceptualization, B.L. (Bin Li); Data curation, B.L. (Bing Liu); Formal analysis, B.L. (Bing Liu) and X.P.; Funding acquisition, X.P., L.H. (Leiping Hou) and B.L. (Bin Li); Investigation, B.L. (Bin Li); Methodology, B.L. (Bing Liu)and B.L. (Bin Li); Project administration, L.H. (Leiping Hou) and B.L. (Bin Li); Resources, B.L. (Bing Liu) and L.H. (Lingjuan Han); Software, B.L. (Bing Liu) and X.P.; Supervision, X.P.; L.H. (Lingjuan Han) and L.H. (Leiping Hou); Validation, X.P. and L.H. (Lingjuan Han); Visualization, B.L. (Bing Liu) and X.P.; Writing—original draft, B.L. (Bing Liu) and X.P.; Writing—review and editing, B.L. (Bin Li). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Scientific and Technological Innovation Programs of Higher Education Institutions in Shanxi (2015144), the National Natural Science Foundation of China (31501806, 31870474), the Shanxi Province Key R&D Plan (201703D211001-04-03), the Science and Technology Innovation Foundation of Shanxi Agricultural University (2014YJ14), the Jinzhong Key Science and Technology Innovation Platform Project (P171002-1), and the Natural Science Foundation of Jiangsu Province (No. BK20151009).
Conflicts of Interest
There are no conflicts of interest related to this work.
References
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- D’Amelia, L.; Dell’Aversana, E.; Woodrow, P.; Ciarmiello, L.F.; Carillo, P. Metabolomics for crop improvement against salinity stress. In Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S.H., Suprasanna, P., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 267–287. [Google Scholar] [CrossRef]
- Mustafa, G.; Akhtar, M.S.; Abdullah, R. Global Concern for Salinity on Various Agro-Ecosystems. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Akhtar, M., Ed.; Springer: Singapore, 2019; pp. 1–19. [Google Scholar] [CrossRef]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Gilliham, M. Energy costs of salt tolerance in crop plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [Green Version]
- Arbona, V.; Manzi, M.; Ollas, C.D.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Baniasadi, F.; Saffari, V.R.; Moud, A.A.M. Physiological and growth responses of Calendula officinalis L. plants to the interaction effects of polyamines and salt stress. Sci. Hortic. 2018, 234, 312–317. [Google Scholar] [CrossRef]
- Jia, X.; Li, H.; Guo, H.; Wang, T.; Zhang, Z.; Qiao, Y.; Guo, Z. Metabolomics advances in horticultural crops. Chin. Agr. Sci. Bul. 2014, 30, 75–83. [Google Scholar] [CrossRef]
- Kim, H.K.; Verpoorte, R. Sample preparation for plant metabolomics. Phytochem. Anal. 2010, 21, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Jiménez-Bremont, J.F.; Ruiz, O.A.; Rodríguez-Kessler, M. Modulation of spermidine and spermine levels in maize seedlings subjected to long-term salt stress. Plant Physiol. Biochem. 2007, 45, 812–821. [Google Scholar] [CrossRef]
- López-Gómez, M.; Hidalgo-Castellanos, J.; Muñoz-Sánchez, J.R.; Marín-Peña, A.J.; Lluch, C.; Herrera-Cervera, J.A. Polyamines contribute to salinity tolerance in the symbiosis Medicago truncatula-Sinorhizobium meliloti by preventing oxidative damage. Plant Physiol. Biochem. 2017, 116, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.; Shan, X.; Li, B.; Shu, S.; Sun, J.; Guo, S. Comparative proteomic analysis reveals the positive effect of exogenous spermidine on photosynthesis and salinity tolerance in cucumber seedlings. Plant Cell Rep. 2016, 35, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Yuan, Y.; Shu, S.; Sun, J.; Guo, S.; Yuan, R.; Tang, Y. Effects of exogenous putrescine on glycolysis and Krebs cycle metabolism in cucumber leaves subjected to salt stress. Plant Growth Regul. 2016, 79, 319–330. [Google Scholar] [CrossRef]
- Zhou, H.; Guo, S.; Shao, H.; Chen, X.; Wei, B.; Sun, J. Effects of iso-smotic Ca(NO3)2 and NaCl stress on growth and physiological characteristics of cucumber seedlings. Acta Ecol. Sin. 2014, 34, 1880–1890. [Google Scholar] [CrossRef]
- Wu, J.; Shu, S.; Li, C.; Sun, J.; Guo, S. Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol. Biochem. 2018, 128, 152–162. [Google Scholar] [CrossRef]
- Han, B.; Xu, G.; Guo, S.; He, C.; Sun, Y.; Gao, W.; Shi, L. Growth and physiological metabolism of cucumber seedlings in response to different concentrations of salt stress. Jiangsu J. Agr. Sci. 2014, 30, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Haling, R.E.; Richardson, A.E.; Culvenor, R.A.; Lambers, H.; Simpson, R.J. Root morphology, root-hair development and rhizosheath formation on perennial grass seedlings is influenced by soil acidity. Plant Soil 2010, 335, 457–468. [Google Scholar] [CrossRef]
- Li, B.; He, L.; Guo, S.; Li, J.; Yang, Y.; Yan, B.; Li, J. Proteomics reveal cucumber Spd-responses under normal condition and salt stress. Plant Physiol. Biochem. 2013, 67, 7–14. [Google Scholar] [CrossRef]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Zhao, L. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.H.; Newbigin, E.D.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 4089–4103. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Williams, T.C.R.; Poolman, M.G.; Howden, A.J.M.; Schwarzlander, M.; Fell, D.A.; Ratcliffe, R.G. A genome-scale metabolic model accurately predicts fluxes in central carbon metabolism under stress conditions. Plant Physiol. 2010, 154, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.F.; Tomlinson, J.D.; Caldwell, R.A. Effect of salts on the activity of carrot phosphofructokinase. Plant Physiol. 1980, 66, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Shu, S.; Du, J.; Yuan, Y.; Xing, W.; Guo, S.; Sun, J. Proteome analysis of roots in cucumber seedlings under iso-osmotic NaCl and Ca (NO3)2 stresses. Plant Mol. Biol. Rep. 2016, 34, 303–317. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Guo, R.; Jiao, Y.; Jin, X.; Zhang, H.; Shi, L. Comparison of salt tolerance in Soja based on metabolomics of seedling roots. Front. Plant Sci. 2017, 8, 1101. [Google Scholar] [CrossRef]
- Shen, Q.; Yu, J.; Fu, L.; Wu, L.; Dai, F.; Jiang, L.; Zhang, G. Ionomic, metabolomic and proteomic analyses reveal molecular mechanisms of root adaption to salt stress in Tibetan wild barley. Plant Physiol. Biochem. 2018, 123, 319–330. [Google Scholar] [CrossRef]
- Chandler, A.M. Amino acid metabolism. In Biochemistry, 2nd ed.; Briggs, T., Chandler, A.M., Eds.; Springer: New York, NY, USA, 1995; pp. 80–103. [Google Scholar] [CrossRef]
- Yu, J.; Si, Y. Metabolic carbon fluxes and biosynthesis of polyhydroxyalkanoates in Ralstonia eutropha on short chain fatty acids. Biotechnol. Prog. 2004, 20, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Lei, L.; Liu, B.; Chen, W.; Zhang, R.; Wang, L.; Lin, Z. Effects of different inhibitors such as malonic acid, Na3PO4 and HgCl2 on uptake of different forms of antimony in rice plant. Plant Soil 2019, 445, 259–271. [Google Scholar] [CrossRef]
- Huan, L.; Xie, X.; Zheng, Z.; Sun, F.; Wu, S.; Li, M.; Wang, G. Positive correlation between PSI response and oxidative pentose phosphate pathway activity during salt stress in an intertidal macroalga. Plant Cell Physiol. 2014, 55, 1395–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orwat, J.; Sarkar, D.; Osorno, J.; Shetty, K. Improved salinity resilience in black bean by seed elicitation using organic compounds. Agron. J. 2017, 109, 1991–2003. [Google Scholar] [CrossRef]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; Gomez-Rodriguez, M.V.; Chaki, M.; Pedrajas, J.R.; Barroso, J.B. The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ. 2006, 29, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.S.; Cha, Y.H.; Kim, H.S.; Kim, N.H.; Yook, J.I. The pentose phosphate pathway as a potential target for cancer therapy. Biomol. Ther. 2018, 26, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.K.; Fatima, T.; Topuz, M.; Bernadec, A.; Sicher, R.; Handa, A.K.; Mattoo, A.K. Pathogenesis-related protein 1b1 (PR1b1) is a major tomato fruit protein responsive to chilling temperature and upregulated in high polyamine transgenic genotypes. Front. Plant Sci. 2016, 7, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, N.J.; Schaewen, A.V. The oxidative pentose phosphate pathway: Structure and organisation. Curr. Opin. Plant Biol. 2003, 6, 236–246. [Google Scholar] [CrossRef]
- Handley, L.W.; Pharr, D.M.; McFeeters, R.F. Carbohydrate changes during maturation of cucumber fruit: Implications for sugar metabolism and transport. Plant Physiol. 1983, 72, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Irving, D.E.; Hurst, P.L.; Ragg, J.S. Changes in carbohydrates and carbohydrate metabolizing enzymes during the development, maturation, and ripening of buttercup squash (Cucurbita maxima D. Delica’). J. Am. Soc. Hortic. Sci. 1997, 122, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Shinozaki, K. Important roles of drought- and cold- inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, T.B.; Budzinski, I.G.; Marur, C.J.; Petkowicz, C.L.; Pereira, L.F.; Vieira, L.G. Expression of three galactinol synthase isoforms in Coffea arabica L. and accumulation of raffinose and stachyose in response to abiotic stresses. Plant Physiol. Biochem. 2011, 49, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Chakraborty, M.; Saha, J.; Gupta, B.; Gupta, K. Polyamines: Osmoprotectants in plant abiotic stress adaptation. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Iqbal, N., Nazar, R., Eds.; Springer: New Delhi, India, 2016; pp. 97–127. [Google Scholar] [CrossRef]
- Fernandez, O.; Béthencourt, L.; Quero, A.; Sangwan, R.S.; Clément, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, L.E.; Paul, M.J.; Wingler, A. How do sugars regulate plant growth and development? New insight into the role of trehalose-6-phosphate. Mol. Plant. 2013, 6, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Tsai, A.Y.L. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: The emerging picture. Front. Plant Sci. 2014, 5, 119. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK and SNF1: Snuffing out stress. Cell Metab. 2007, 6, 339–340. [Google Scholar] [CrossRef] [Green Version]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938. [Google Scholar] [CrossRef]
- Emanuelle, S.; Doblin, M.S.; Stapleton, D.I.; Bacic, A.; Gooley, P.R. Molecular insights into the enigmatic metabolic regulator, SnRK. Trends Plant Sci. 2016, 21, 341–353. [Google Scholar] [CrossRef]
- Oszvald, M.; Primavesi, L.F.; Griffiths, C.A.; Cohn, J.; Basu, S.S.; Nuccio, M.L.; Paul, M.J. Trehalose 6-phosphate regulates photosynthesis and assimilate partitioning in reproductive tissue. Plant Physiol. 2018, 176, 2623–2638. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Chen, L.; Lu, W.; Sun, J.; Guo, S.; Yuan, Y.; Li, J. Effects of exogenous spermidine on photosynthetic capacity and expression of Calvin cycle genes in salt-stressed cucumber seedlings. J. Plant Res. 2014, 127, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shu, S.; An, Y.; Zhou, H.; Guo, S.; Sun, J. Influence of exogenous spermidine on carbon–nitrogen metabolism under Ca(NO3)2 stress in cucumber root. Plant Growth Regul. 2017, 81, 103–115. [Google Scholar] [CrossRef]
- Joshi, V.; Fernie, A.R. Citrulline metabolism in plants. Amino Acids 2017, 49, 1543–1559. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S.; Funck, D.; Trovato, M.; Forlani, G. Amino acids of the glutamate family: Functions beyond primary metabolism. Front. Plant Sci. 2016, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, D.H.; Pieckenstain, F.L.; Szymanski, J.; Erban, A.; Bromke, M.; Hannah, M.A.; Udvardi, M.K. Comparative functional genomics of salt stress in related model and cultivated plants identifies and overcomes limitations to translational genomics. PLoS ONE 2011, 6, e17094. [Google Scholar] [CrossRef]
- Sicher, R.C.; Timlin, D.; Bailey, B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration. J. Plant Physiol. 2012, 169, 686–695. [Google Scholar] [CrossRef]
- Triplett, E.W.; Blevins, D.G.; Randall, D.D. Allantoic acid synthesis in soybean root nodule cytosol via xanthine dehydrogenase. Plant Physiol. 1980, 65, 1203–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, R.M.; Bhattacharya, S.; Bavaria, M.N.; Viar, M.J.; Johnson, L.R. Spermidine, a sensor for antizyme 1 expression regulates intracellular polyamine homeostasis. Amino Acids 2014, 46, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; He, S.; Zhai, H.; Wang, L.; Zhao, Y.; Wang, B.; Liu, Q. Overexpression of IbP5CR enhances salt tolerance in transgenic sweet potato. Plant Cell Tissue Organ. Cult. 2014, 117, 1–16. [Google Scholar] [CrossRef]
- López-Gómez, M.; Hidalgo-Castellanos, J.; Iribarne, C.; Lluch, C. Proline accumulation has prevalence over polyamines in nodules of Medicago sativa in symbiosis with Sinorhizobium meliloti during the initial response to salinity. Plant Soil 2014, 374, 149–159. [Google Scholar] [CrossRef]
- Ludwig, R.A. Arabidopsis chloroplasts dissimilate L-arginine and L-citrulline for use as N source. Plant Physiol. 1993, 101, 429–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, D.E.; Madore, M.A. Patterns of assimilate production and translocation in muskmelon (Cucumis melo L.): II. Low temperature effects. Plant Physiol. 1992, 99, 966–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akashi, K.; Miyake, C.; Yokota, A. Citrulline, a novel compatible solute in drought-tolerant wild watermelon leaves, is an efficient hydroxyl radical scavenger. FEBS Lett. 2001, 508, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Massange-Sanchez, J.A.; Palmeros-Suarez, P.A.; Espitia-Rangel, E.; Rodriguez-Arevalo, I.; Sanchez-Segura, L.; Martinez-Gallardo, N.A.; Delano-Frier, J.P. Overexpression of grain amaranth (Amaranthus hypochondriacus) AhERF or AhDOF transcription factors in Arabidopsis thaliana increases water deficit-and salt-stress tolerance, respectively, via contrasting stress-amelioration mechanisms. PLoS ONE 2016, 11, e0164280. [Google Scholar] [CrossRef] [PubMed]
- Roosens, N.H.; Thu, T.T.; Iskandar, H.M.; Jacobs, M. Isolation of the ornithine-δ-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant Physiol. 1998, 117, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Campos, M.K.F.; de Carvalho, K.; de Souza, F.S.; Marur, C.J.; Pereira, L.F.P.; Filho, J.C.; Vieira, L.G.E. Drought tolerance and antioxidant enzymatic activity in transgenic ‘Swingle’ citrumelo plants over-accumulating proline. Environ. Exp. Bot. 2011, 72, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Ghars, M.A.; Parre, E.; Debez, A.; Bordenave, M.; Richard, L.; Leport, L.; Abdelly, C. Comparative salt tolerance analysis between Arabidopsis thaliana and Thellungiella halophila, with special emphasis on K+/Na+ selectivity and proline accumulation. J. Plant Physiol. 2008, 165, 588–599. [Google Scholar] [CrossRef]
- Gessler, P.; Buchal, P.; Schwenk, H.U.; Wermuth, B. Favourable long-term outcome after immediate treatment of neonatal hyperammonemia due to N-acetylglutamate synthase deficiency. Eur. J. Pediatr. 2010, 169, 197–199. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Xu, J.; Liu, T.; Yang, S.; Jin, X.; Qu, F.; Huang, N.; Hu, X. Polyamines are involved in GABA-regulated salinity-alkalinity stress tolerance in muskmelon. Environ. Exp. Bot. 2019, 164, 181–189. [Google Scholar] [CrossRef]
- Jin, X.; Liu, T.; Xu, J.; Gao, Z.; Hu, X. Exogenous GABA enhances muskmelon tolerance to salinity-alkalinity stress by regulating redox balance and chlorophyll biosynthesis. BMC Plant Biol. 2019, 19, 48. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, Z.; Xu, W.; Li, J.; Zhao, N.; Zhou, Y. Application of γ-aminobutyric acid demonstrates a protective role of polyamine and GABA metabolism in muskmelon seedlings under Ca(NO3)2 stress. Plant Physiol. Biochem. 2015, 92, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Lee, I.J. Regulation of salicylic acid, jasmonic acid and fatty acids in cucumber (Cucumis sativus L.) by spermidine promotes plant growth against salt stress. Acta Physiol. Plant. 2013, 35, 3315–3322. [Google Scholar] [CrossRef]
- Hao, J.H.; Wang, X.L.; Dong, C.J.; Zhang, Z.G.; Shang, Q.M. Salicylic acid induces stomatal closure by modulating endogenous hormone levels in cucumber cotyledons. Russ. J. Plant Physiol. 2011, 58, 906. [Google Scholar] [CrossRef]
- Feng, H.Y.; Wang, Z.M.; Kong, F.N.; Zhang, M.J.; Zhou, S.L. Roles of carbohydrate supply and ethylene, polyamines in maize kernel set. J. Integr. Plant Biol. 2011, 53, 388–398. [Google Scholar] [CrossRef]
- Hagège, D.; Kevers, C.; Geuns, J.; Gaspar, T. Ethylene production and polyamine content of fully habituated sugarbeet calli. J. Plant Physiol. 1994, 143, 722–725. [Google Scholar] [CrossRef]
- Kacperska, A.; Kubacka-Zębalska, M. Formation of stress ethylene depends both on ACC synthesis and on the activity of free radical-generating system. Physiol. Plant. 1989, 77, 231–237. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Glick, B.R.; Chauhan, P.S.; Yim, W.; Sa, T. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1-carboxylic acid deaminase activity. Plant Physiol. Biochem. 2011, 49, 427–434. [Google Scholar] [CrossRef]
Figure 1.
Effect of spermidine (Spd) on plant growth of cucumber seedlings under salt stress after 5-day treatment. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd.
Figure 1.
Effect of spermidine (Spd) on plant growth of cucumber seedlings under salt stress after 5-day treatment. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd.

Figure 2.
Effect of Spd on growth parameters of cucumber seedlings under salt stress. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd. Data are expressed as the mean ± standard error. Different letters indicate significant differences at p < 0.05, according to Duncan’s new multiple range test.
Figure 2.
Effect of Spd on growth parameters of cucumber seedlings under salt stress. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd. Data are expressed as the mean ± standard error. Different letters indicate significant differences at p < 0.05, according to Duncan’s new multiple range test.

Figure 3.
Principle components analysis (PCA) on metabolite profiles of the roots in cucumber seedlings under normal condition and salt stress with or without Spd. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd; (a) Scores plot; (b) Loading plot. Each number in loadings plot corresponds to a metabolite and its contribution to the principal components, as shown in Table S1.
Figure 3.
Principle components analysis (PCA) on metabolite profiles of the roots in cucumber seedlings under normal condition and salt stress with or without Spd. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd; (a) Scores plot; (b) Loading plot. Each number in loadings plot corresponds to a metabolite and its contribution to the principal components, as shown in Table S1.

Figure 4.
Proposed metabolic network changes in the roots of cucumber seedlings obtained through (orthogonal) partial least-squares-discriminant analysis ((O)PLS-DA) under normal condition and salt stress with or without Spd. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd; Significant difference at p < 0.05.
Figure 4.
Proposed metabolic network changes in the roots of cucumber seedlings obtained through (orthogonal) partial least-squares-discriminant analysis ((O)PLS-DA) under normal condition and salt stress with or without Spd. CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd; Significant difference at p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Root other metabolites responsive to salt stress and Spd (Metabolites not shown in Figure 4).
Table 1.
Root other metabolites responsive to salt stress and Spd (Metabolites not shown in Figure 4).
| Metabolite Names | Relative Concentration | Fold Changes | VIP | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CK | S | SS | Log2 (S/CK) | Log2 (SS/S) | Log2 (SS/CK) | S/CK | SS/S | SS/CK | |
| Simple sugars | |||||||||
| Fructose | 15.74 ± 0.61 | 11.86 ± 0.69 | 11.95 ± 0.76 | −0.41 ** | 0.01 | −0.40 ** | 0.95 | 0.02 | 0.9 |
| Mannose | 132.87 ± 6.19 | 56.64 ± 3.17 | 55.38 ± 3.63 | −1.23 *** | −0.03 | −1.26 *** | 1.21 | 0.13 | 1.21 |
| Xylose | 21.64 ± 0.67 | 8.44 ± 0.58 | 5.25 ± 0.3 | −1.36 *** | −0.68 *** | −2.04 *** | 1.22 | 1.2 | 1.26 |
| Erythrose | 5.66 ± 0.32 | 2.14 ± 0.22 | 8.61 ± 0.08 | −1.41 *** | 2.01 *** | 0.61 *** | 1.16 | 1.44 | 1.14 |
| Galactose | 31.89 ± 1.98 | 154.64 ± 17.37 | 125.09 ± 6.81 | 2.28 *** | −0.31 | 1.97 *** | 1.13 | 0.25 | 1.25 |
| Sedoheptulose | 101.88 ± 6.41 | 45.73 ± 2.47 | 34.28 ± 1.29 | −1.16 *** | −0.42 ** | −1.57 *** | 1.18 | 1.12 | 1.24 |
| Glucoheptose | 6.38 ± 0.34 | 8.03 ± 0.58 | 5.35 ± 0.49 | 0.33 * | −0.59 ** | −0.25 | 0.71 | 1.03 | 0.57 |
| Talose | 3.38 ± 0.12 | 2.09 ± 0.07 | 1.51 ± 0.03 | −0.69 *** | −0.47 *** | −1.16 *** | 1.19 | 1.35 | 1.26 |
| Allose | 0.82 ± 0.04 | 0.39 ± 0.02 | 0.38 ± 0.03 | −1.07 *** | −0.04 | −1.11 *** | 1.18 | 0.22 | 1.15 |
| Altrose | 3.16 ± 0.25 | 11.29 ± 0.6 | 14.74 ± 0.77 | 1.84 *** | 0.38 ** | 2.22 *** | 1.22 | 1.03 | 1.25 |
| Oligoses | |||||||||
| Maltotriose | 3.83 ± 0.27 | 1.37 ± 0.04 | 2.61 ± 0.2 | −1.48 *** | 0.93 *** | −0.55 ** | 1.22 | 1.34 | 0.89 |
| Cellobiose | 9.14 ± 0.53 | 1.99 ± 0.27 | 1.69 ± 0.09 | −2.20 *** | −0.23 | −2.43 *** | 1.01 | 0.06 | 1.26 |
| Leucrose | 0.03 ± 0.01 | 0.04 ± 0 | 0.05 ± 0.01 | 0.58 | 0.2 | 0.78 | 0.55 | 0.24 | 0.61 |
| Melibiose | 12.94 ± 0.63 | 9.48 ± 0.91 | 4.89 ± 0.44 | −0.45 ** | −0.96 *** | −1.40 *** | 0.83 | 1.19 | 1.18 |
| Polysaccharide | |||||||||
| Levoglucosan | 4.04 ± 0.27 | 12.13 ± 0.77 | 9.15 ± 0.59 | 1.59 *** | −0.41 * | 1.18 *** | 1.22 | 0.96 | 1.18 |
| Sugar phosphate esters | |||||||||
| Glucose-1-P | 69.52 ± 6.18 | 357.65 ± 20.12 | 529.59 ± 81.17 | 2.36 *** | 0.57 * | 2.93 *** | 1.24 | 0.82 | 1.23 |
| Sugar acids | |||||||||
| Saccharic acid | 0.56 ± 0.03 | 0.94 ± 0.03 | 8.95 ± 0.42 | 0.76 *** | 3.25 *** | 4.00 *** | 1.13 | 1.49 | 1.27 |
| Glucuronic acid | 8.78 ± 0.38 | 9.7 ± 0.14 | 13.95 ± 0.42 | 0.14 * | 0.53 *** | 0.67 *** | 0.68 | 1.41 | 1.18 |
| Gluconic acid | 20.39 ± 0.84 | 25.3 ± 0.65 | 30.2 ± 0.9 | 0.31 *** | 0.26 ** | 0.57 *** | 0.99 | 1.16 | 1.16 |
| Gluconic lactone | 2.08 ± 0.07 | 1.05 ± 0.1 | 1.64 ± 0.28 | −0.98 *** | 0.64 | −0.35 | 1.13 | 0.58 | 0.55 |
| Glucoheptonic acid | 0.66 ± 0.04 | 1.32 ± 0.06 | 0.83 ± 0.07 | 1.00 *** | −0.67 *** | 0.33 | 1.17 | 1.19 | 0.65 |
| Galactonic acid | 5.68 ± 0.26 | 10 ± 0.35 | 8.8 ± 0.56 | 0.82 *** | −0.18 | 0.63 *** | 1.18 | 0.67 | 1.06 |
| Threonic acid | 14.82 ± 0.61 | 35.47 ± 2.16 | 39.15 ± 1.01 | 1.26 *** | 0.14 | 1.40 *** | 1.22 | 0.63 | 1.26 |
| Lactobionic acid | 14.07 ± 1.03 | 5.69 ± 0.38 | 8.69 ± 0.7 | −1.31 *** | 0.61 ** | −0.69 *** | 1.18 | 1.08 | 0.98 |
| Glucosaminic acid | 2.54 ± 0.07 | 3 ± 0.17 | 2.6 ± 0.16 | 0.24 * | 0.21 | 0.03 | 0.73 | 0.66 | 0.07 |
| Amino sugar | |||||||||
| N-acetyl-β-D-mannosamine | 1.08 ± 0.1 | 1.46 ± 0.22 | 1.83 ± 0.07 | 0.43 | 0.33 | 0.76 *** | 0.03 | 0.47 | 1 |
| Deoxy suger | |||||||||
| 2-Deoxy-D-galactose | 0.22 ± 0.02 | 0.41 ± 0.01 | 0.32 ± 0.02 | 0.89 *** | −0.36 ** | 0.53 ** | 1.12 | 1.12 | 0.92 |
| Sugar alcohols | |||||||||
| Ribitol | 5.53 ± 0.18 | 3.01 ± 0.18 | 2.03 ± 0.14 | −0.88 *** | −0.57 ** | −1.44 *** | 1.18 | 1.14 | 1.23 |
| Threitol | 0.93 ± 0.01 | 2.23 ± 0.15 | 2.11 ± 0.12 | 1.25 *** | −0.08 | 1.18 *** | 1.21 | 0.19 | 1.24 |
| Allo-inositol | 0.31 ± 0.02 | 0.34 ± 0.02 | 0.28 ± 0.02 | 0.13 | −0.31 * | −0.18 | 0.36 | 0.83 | 0.49 |
| 1,5-Anhydroglucitol | 1.84 ± 0.42 | 1.92 ± 0.04 | 1.84 ± 0.1 | 0.06 | −0.06 | 0 | 0.33 | 0.41 | 0.21 |
| Carboxylic acids | |||||||||
| Maleic acid | 0.6 ± 0.02 | 0.29 ± 0.03 | 0.18 ± 0.02 | −1.06 *** | −0.70 ** | −1.77 *** | 1.15 | 1.11 | 1.24 |
| Glyceric acid | 14.17 ± 0.35 | 10.99 ± 0.33 | 11.57 ± 0.43 | −0.37 *** | 0.07 | −0.29 *** | 1.11 | 0.41 | 1 |
| Glycolic acid | 0.89 ± 0.07 | 0.55 ± 0.07 | 0.25 ± 0.02 | −0.7 ** | −1.14 *** | −1.83 *** | 0.9 | 1.24 | 1.22 |
| 2-Furoic acid | 2.17 ± 0.56 | 0.39 ± 0.09 | 0 | −2.46 *** | OS ** | OS ** | 0.99 | 1.37 | 1.23 |
| Lactic acid | 65.74 ± 4.89 | 48.74 ± 3.45 | 32.82 ± 2.03 | −0.43 * | −0.58 ** | −1.00 *** | 0.76 | 1.08 | 1.14 |
| 3-Hydroxypropionic acid | 0.74 ± 0.01 | 0.44 ± 0.01 | 0.51 ± 0.02 | −0.76 *** | 0.23 ** | −0.53 *** | 1.25 | 1.13 | 1.2 |
| 3-Hydroxybutyric acid | 2.21 ± 0.11 | 0.9 ± 0.05 | 0.62 ± 0.04 | −1.30 *** | −0.54 *** | −1.83 *** | 1.22 | 1.17 | 1.25 |
| 2-Hydroxybutanoic acid | 363.26 ± 17.41 | 240.05 ± 6.36 | 185.64 ± 5.56 | −0.60 *** | −0.37 *** | −0.97 *** | 1.13 | 1.3 | 1.22 |
| Benzoic acid | 0.72 ± 0.04 | 0.98 ± 0.13 | 1.64 ± 0.1 | 0.44 | 0.75 ** | 1.18 *** | 0.58 | 1.11 | 1.2 |
| Quinoline-4-carboxylic acid | 0.21 ± 0.01 | 0.19 ± 0.01 | 0.12 ± 0.01 | −0.19 | −0.61 | −0.79 | 0.42 | 0.78 | 0.66 |
| Pyrrole-2-carboxylic acid | 0.26 ± 0.04 | 0.34 ± 0.03 | 0.19 ± 0.02 | 0.39 | −0.81 *** | −0.42 | 0.59 | 1.25 | 0.41 |
| Pipecolinic acid | 15.85 ± 0.49 | 21.37 ± 0.74 | 25.23 ± 0.24 | 0.43 *** | 0.24 *** | 0.67 *** | 1.1 | 1.18 | 1.24 |
| 2-Ketobutyric acid | 1.67 ± 0.08 | 4.75 ± 0.38 | 4.76 ± 0.1 | 1.51 *** | 0 | 1.51 *** | 1.21 | 0.1 | 1.26 |
| Polyphenols | |||||||||
| Gallic acid | 7.11 ± 0.39 | 5.53 ± 0.35 | 3.6 ± 0.18 | −0.36 * | −0.62 *** | −0.98 *** | 0.81 | 1.22 | 1.19 |
| Pyrogallol | 0.13 ± 0 | 0.16 ± 0.01 | 0.11 ± 0.01 | 0.3 * | −0.54 ** | −0.24 * | 0.74 | 1.09 | 0.73 |
| 1,2,4-Benzenetriol | 0.17 ± 0.01 | 0.18 ± 0.01 | 0.1 ± 0.01 | 0.04 | −0.87 ** | −0.83 *** | 0.07 | 1.15 | 1 |
| Neohesperidin | 2.95 ± 0.17 | 2.08 ± 0.1 | 1.58 ± 0.17 | −0.51 *** | −0.4 | −0.9 * | 0.98 | 0.79 | 0.72 |
| Alcohols | |||||||||
| Acetol | 0.55 ± 0.08 | 0.82 ± 0.08 | 0.57 ± 0.04 | 0.57 | −0.52 * | 0.05 | 0.63 | 0.87 | 0.18 |
| Cuminic alcohol | 0.56 ± 0.1 | 0.36 ± 0.01 | 0.33 ± 0.09 | −0.64 | −0.14 ** | −0.78 ** | 0.63 | 1.03 | 0.92 |
| Cyclohexane-1,2-diol | 2.09 ± 0.06 | 0.97 ± 0.25 | 2.73 ± 0.17 | −1.10 * | 1.49 | 0.38 | 0.86 | 0.25 | 0.25 |
| Dodecanol | 0.56 ± 0.04 | 0.52 ± 0.04 | 0.34 ± 0.02 | −0.11 | −0.60 *** | −0.71 *** | 0.26 | 1.17 | 1.09 |
| Diglycerol | 4.29 ± 0.03 | 1.46 ± 0.06 | 0.84 ± 0.11 | −1.56 *** | −0.79 * | −2.34 ** | 1.26 | 0.86 | 0.87 |
| Glycerol | 522.34 ± 77.62 | 390.08 ± 12.75 | 475.12 ± 21.25 | −0.42 | 0.28 *** | −0.13 | 0.19 | 1.03 | 0.26 |
| Fatty acids | |||||||||
| Stearic acid | 0.25 ± 0.01 | 0.32 ± 0.02 | 0.28 ± 0.01 | 0.34 ** | −0.21 | 0.14 | 0.83 | 0.65 | 0.43 |
| Palmitic acid | 10.3 ± 0.86 | 5.81 ± 0.64 | 4.48 ± 0.86 | −0.83 *** | −0.37 | −1.20 ** | 0.97 | 0.56 | 0.96 |
| Heptadecanoic acid | 1.13 ± 0.25 | 0.43 ± 0.12 | 0.05 ± 0.02 | −1.38 ** | −3.08 *** | −4.46 *** | 0.89 | 1.3 | 1.13 |
| Azelaic acid | 0.22 ± 0.01 | 0.26 ± 0.01 | 0.17 ± 0.01 | 0.25 * | −0.60 *** | −0.34 ** | 0.8 | 1.37 | 0.96 |
| Simple lipids | |||||||||
| Monostearin | 0.64 ± 0 | 0.3 ± 0.03 | 0.56 ± 0.07 | −1.06 *** | 0.87 | −0.19 * | 1.13 | 0.68 | 0.76 |
| 1-Monopalmitin | 2.5 ± 0.33 | 1.47 ± 0.1 | 2.27 ± 0.16 | −0.77 | 0.63 ** | −0.14 | 0.47 | 1.11 | 0.04 |
| 2-Monopalmitin | 0.79 ± 0.09 | 0.7 ± 0.05 | 1.18 ± 0.11 | −0.18 | 0.75 ** | 0.57 * | 0.25 | 1.13 | 0.78 |
| Other aspartate family | |||||||||
| Threonine | 2.36 ± 0.04 | 1.77 ± 0.04 | 3.42 ± 0.03 | −0.41 *** | 0.95 *** | 0.54 *** | 1.21 | 1.48 | 1.26 |
| Isoleucine | 33.33 ± 0.54 | 30.55 ± 1.28 | 23.53 ± 0.61 | −0.13 | −0.38 *** | −0.50 *** | 0.61 | 1.21 | 1.22 |
| Aromatic amino acids | |||||||||
| Tryptophan | 8.75 ± 0.36 | 12.96 ± 0.49 | 10.2 ± 0.43 | 0.57 *** | −0.35 ** | 0.22 * | 1.13 | 1.13 | 0.74 |
| Phenylalanine | 0.83 ± 0.05 | 1.43 ± 0.08 | 0.86 ± 0.1 | 0.79 *** | −0.73 ** | 0.06 | 1.1 | 1.13 | 0.03 |
| Tyrosine | 161.38 ± 8 | 125.23 ± 6.4 | 92.65 ± 3.99 | −0.37 ** | −0.43 ** | −0.80 *** | 0.88 | 1.14 | 1.17 |
| Other amino acids | |||||||||
| Oxoproline | 224.1 ± 16.39 | 615.38 ± 17.81 | 428.64 ± 20.67 | 1.46 *** | −0.52 *** | 0.94 *** | 1.22 | 1.27 | 1.13 |
| α-Aminoadipic acid | 11.92 ± 0.44 | 8.1 ± 0.17 | 5.48 ± 0.15 | −0.56 *** | −0.56 *** | −1.12 *** | 1.17 | 1.42 | 1.25 |
| Creatine degr | 5.88 ± 0.36 | 7.06 ± 0.21 | 6.46 ± 0.29 | 0.27 * | −0.13 | 0.14 | 0.74 | 0.64 | 0.42 |
| N-Carbamylglutamate | 3.61 ± 0.31 | 6.87 ± 0.31 | 9.74 ± 0.14 | 0.93 *** | 0.50 *** | 1.43 *** | 1.1 | 1.33 | 1.21 |
| Cycloleucine | 0.33 ± 0.02 | 0.48 ± 0.06 | 0.53 ± 0.11 | 0.55 | 0.16 | 0.71 | 0.59 | 0.05 | 0.51 |
| Cysteinylglycine | 0 | 1.05 ± 0.27 | 0.51 ± 0.09 | OS ** | −1.04 | OS ** | 1.11 | 0.42 | 0.99 |
| Maleimide | 0.1 ± 0 | 0.23 ± 0.01 | 0.25 ± 0.01 | 1.20 *** | 0.15 | 1.35 *** | 1.26 | 0.62 | 1.25 |
| Oxamide | 0.68 ± 0.01 | 6.66 ± 0.36 | 6.14 ± 0.23 | 3.30 *** | −0.12 | 3.18 | 1.27 | 0.58 | 0.02 |
| Nucleotide metabolic pathway | |||||||||
| Uracil | 4.28 ± 0.47 | 4.36 ± 0.44 | 2.94 ± 0.15 | 0.03 | −0.57 ** | −0.54 * | 0.08 | 1.07 | 0.75 |
| Thymine | 0.24 ± 0.01 | 0.11 ± 0 | 0.19 ± 0 | −1.13 *** | 0.84 *** | −0.30 ** | 1.23 | 1.46 | 0.92 |
| Adenine | 3.22 ± 0.1 | 2.59 ± 0.08 | 2 ± 0.09 | −0.31 *** | −0.38 ** | −0.69 *** | 1.01 | 1.16 | 1.17 |
| Uridine | 4.03 ± 0.52 | 2.98 ± 0.34 | 7.76 ± 0.35 | −0.44 | 1.38 *** | 0.95 ** | 0.42 | 1.3 | 0.96 |
| Guanosine | 12.33 ± 1.66 | 4.06 ± 0.25 | 3.79 ± 0.38 | −1.60 *** | −0.1 | −1.70 *** | 1.11 | 0.3 | 1.1 |
| Adenosine | 97.24 ± 8 | 15.08 ± 0.51 | 35.96 ± 1.01 | −2.69 *** | 1.25 *** | −1.44 *** | 1.25 | 1.47 | 1.22 |
| Inosine | 1.37 ± 0.09 | 0.83 ± 0.05 | 0.42 ± 0.02 | −0.72 ** | −1.00 *** | −1.72 *** | 1.07 | 1.37 | 1.24 |
| Hypoxanthine | 0 | 0.1 ± 0.02 | 0.06 | OS *** | OS * | OS | 1.24 | 1.29 | 1.28 |
| Carbamoyl-aspartic acid | 2.46 ± 0.16 | 3.8 ± 0.22 | 2.26 ± 0.23 | 0.63 *** | −0.75 ** | −0.12 | 1.01 | 1 | 0.27 |
| β-Alanine | 5.64 ± 0.13 | 3.96 ± 0.09 | 4.05 ± 0.06 | −0.51 *** | 0.03 | −0.48 *** | 1.21 | 0.34 | 1.22 |
| 3-Aminoisobutyric acid | 45.2 ± 2.04 | 48.34 ± 1.87 | 27.57 ± 5.41 | 0.1 | −0.81 * | −0.71 * | 0.38 | 0.91 | 0.73 |
| 5,6-Dihydrouracil | 41.11 ± 2.4 | 33.48 ± 1.93 | 19.5 ± 1.2 | −0.30 * | −0.78 *** | −1.08 *** | 0.7 | 1.28 | 1.17 |
| Allantoic acid | 0 | 3.22 ± 0.12 | 6.3 ± 0.04 | OS *** | 0.97 | OS *** | 1.27 | 0.37 | 0.99 |
| Urea | 10.22 ± 0.32 | 20.86 ± 0.89 | 16.08 ± 0.67 | 1.03 *** | −0.38 ** | 0.65 *** | 1.23 | 1.15 | 1.18 |
| Polyamines | |||||||||
| Putrescine | 28.26 ± 2.26 | 25.54 ± 1.62 | 23.03 ± 1.74 | −0.15 | −0.15 | −0.31 | 0.29 | 0.41 | 0.55 |
| Hormones and precursors | |||||||||
| Salicin | 2.73 ± 1.62 | 4.45 ± 0.84 | 6.71 ± 0.93 | 0.71 | 0.59 * | 1.3 * | 0.51 | 1.2 | 0.97 |
| Melatonin | 0.79 ± 0.01 | 0.82 ± 0.02 | 1.49 ± 0.03 | 0.06 | 0.86 | 0.92 | 0.5 | 0.06 | 0.09 |
| α-Ecdysone | 20.49 ± 1.18 | 16.73 ± 0.61 | 17.87 ± 0.99 | −0.29 * | 0.1 | −0.2 | 0.74 | 0.33 | 0.5 |
| Sphingosine | |||||||||
| Phytosphingosine | 0.8 ± 0.09 | 1.61 ± 0.28 | 2.74 ± 0.56 | 1.01 | 0.77 | 1.78 ** | 0.51 | 0.55 | 0.96 |
| D-Erythro-sphingosine | 2.87 ± 0.33 | 7.94 ± 0.62 | 6.38 ± 0.85 | 1.47 *** | −0.32 | 1.15 *** | 1.15 | 0.51 | 0.84 |
| Dihydrosphingosine | 3.58 ± 0.17 | 3.63 ± 0.09 | 5.14 ± 0.15 | 0.02 | 0.5 | 0.52 | 0.12 | 0.46 | 0.41 |
| Catecholamine metabolism pathway | |||||||||
| L-dopa | 1.23 ± 0.07 | 1.75 ± 0.17 | 1.13 ± 0.07 | 0.51 * | −0.63 ** | −0.12 | 0.81 | 1.06 | 0.3 |
| Dehydroascorbic acid | 0.37 ± 0.02 | 0.22 ± 0.01 | 0.17 ± 0.01 | −0.75 *** | −0.41 *** | −1.16 *** | 1.11 | 1.21 | 1.21 |
| Noradrenaline | 9.82 ± 0.63 | 17.28 ± 1.15 | 26.68 ± 2.05 | 0.82 *** | 0.63 *** | 1.44 *** | 1.07 | 1.12 | 1.2 |
| Steroids | |||||||||
| 22-Ketocholesterol | 5.12 ± 0.28 | 132.18 ± 5.73 | 96.63 ± 3.18 | 4.69 *** | −0.45 *** | 4.24 *** | 1.27 | 1.24 | 1.28 |
| 21-Hydroxypregnenolone | 0.35 ± 0.01 | 0.5 ± 0.02 | 0.38 ± 0.01 | 0.51 *** | −0.40 *** | 0.11 | 1.12 | 1.2 | 0.63 |
| 7-Hydroxy-4-androstene-3,17-dione | 0.28 ± 0.02 | 0.19 ± 0.01 | 0 | −0.54 *** | OS *** | OS *** | 1.02 | 1.36 | 1.17 |
| Vitamin | |||||||||
| Nicotinic acid | 2.2 ± 0.04 | 3.03 ± 0.13 | 2.06 ± 0.15 | 0.46 *** | −0.56 ** | −0.1 | 1.12 | 1.14 | 0.35 |
| Nicotinamide | 0.95 ± 0.05 | 0.85 ± 0.11 | 0.72 ± 0.07 | −0.16 | −0.24 | −0.41 * | 0.4 | 0.44 | 0.8 |
CK = control, S = 75 mM NaCl, SS = 75 mM NaCl + 0.1 mM Spd. The relative contents of metabolites are the means of data from eight biological replicates using GC-MS. Data are expressed as the mean ± standard error of eight biological replicates. * Significant difference at p < 0.05. ** Significant difference at p < 0.01. *** Significant difference at p < 0.001. OS, one-sample t-test analysis. Bold numbers are VIP values > 1 and p-values < 0.05.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, B.; Peng, X.; Han, L.; Hou, L.; Li, B. Effects of Exogenous Spermidine on Root Metabolism of Cucumber Seedlings under Salt Stress by GC-MS. Agronomy 2020, 10, 459. https://doi.org/10.3390/agronomy10040459
AMA Style
Liu B, Peng X, Han L, Hou L, Li B. Effects of Exogenous Spermidine on Root Metabolism of Cucumber Seedlings under Salt Stress by GC-MS. Agronomy. 2020; 10(4):459. https://doi.org/10.3390/agronomy10040459
Chicago/Turabian StyleLiu, Bing, Xujian Peng, Lingjuan Han, Leiping Hou, and Bin Li. 2020. "Effects of Exogenous Spermidine on Root Metabolism of Cucumber Seedlings under Salt Stress by GC-MS" Agronomy 10, no. 4: 459. https://doi.org/10.3390/agronomy10040459
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.