The Tomato spotted wilt virus (TSWV) Genome is Differentially Targeted in TSWV-Infected Tomato (Solanum lycopersicum) with or without Sw-5 Gene

 , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Virus and Plant Inoculations
2.2. DAS-ELISA
2.3. RNA Isolation
2.4. Deep Sequencing
2.5. Bioinformatic Analysis
2.6. Primer Design
2.7. RT-qPCR of TSWV Genes
2.8. Data Analysis
3. Results
3.1. Disease Symptoms of Sw-5(−) and Sw-5(+) Genotypes
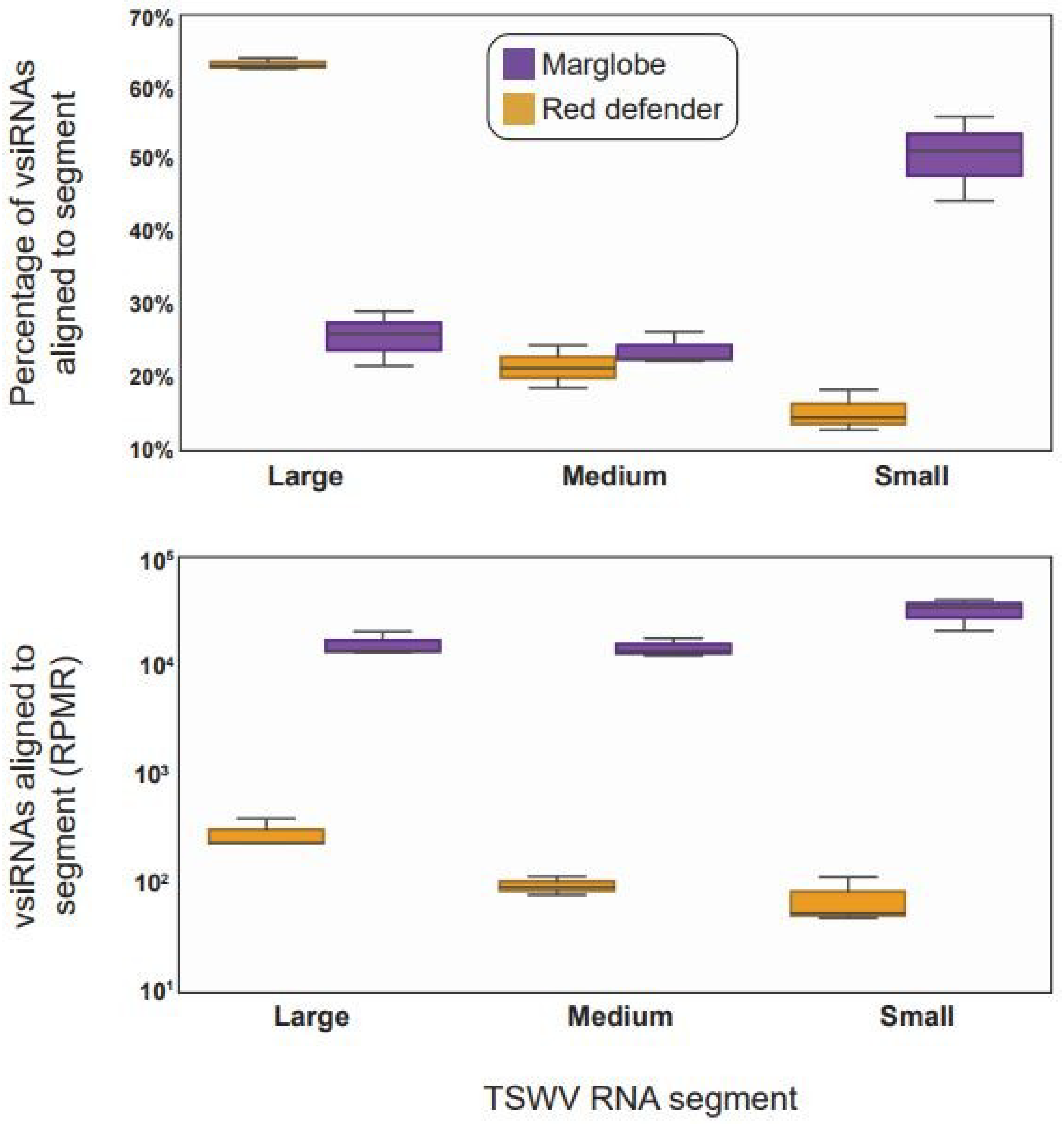
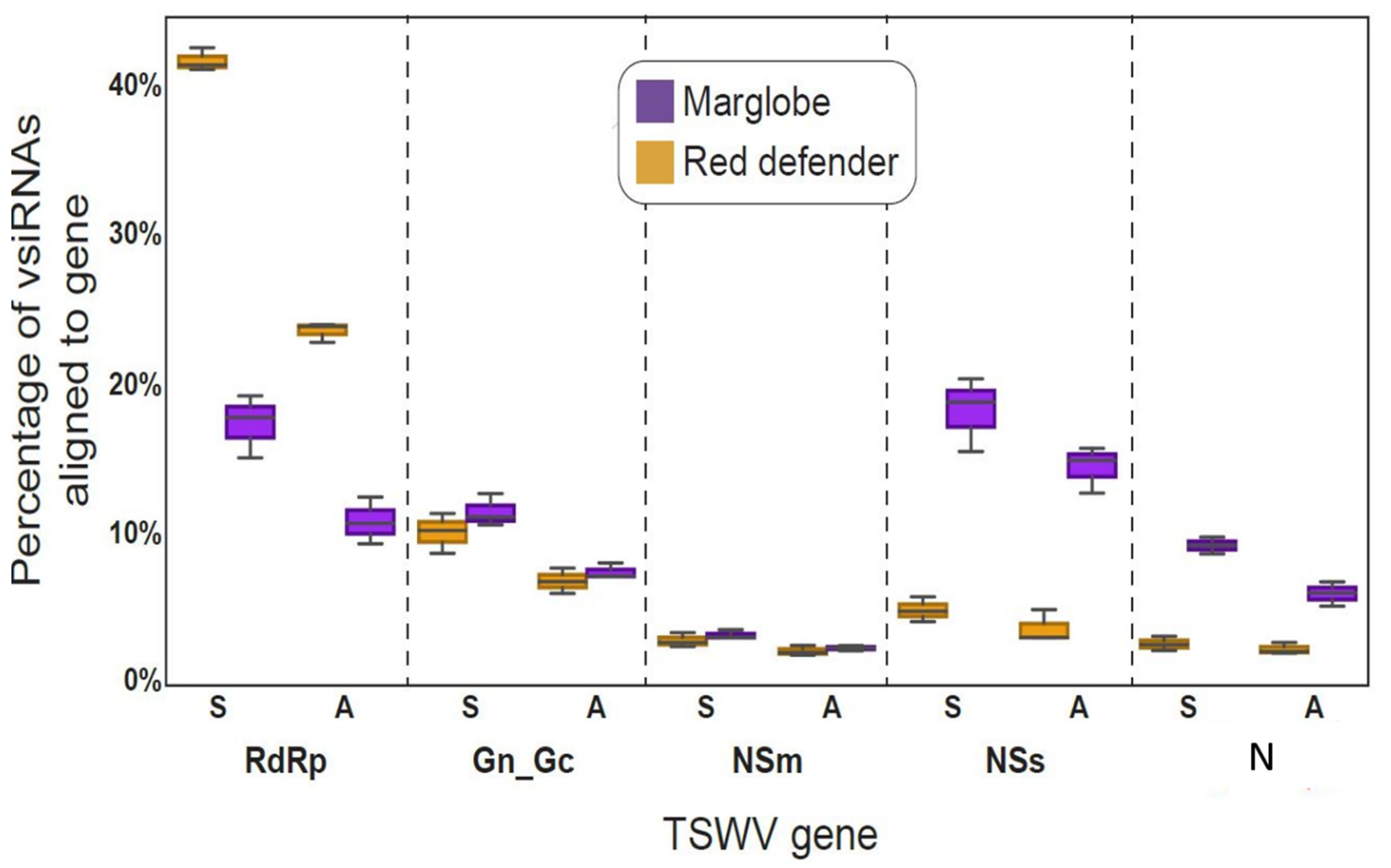
3.2. Profiles of vsiRNAs
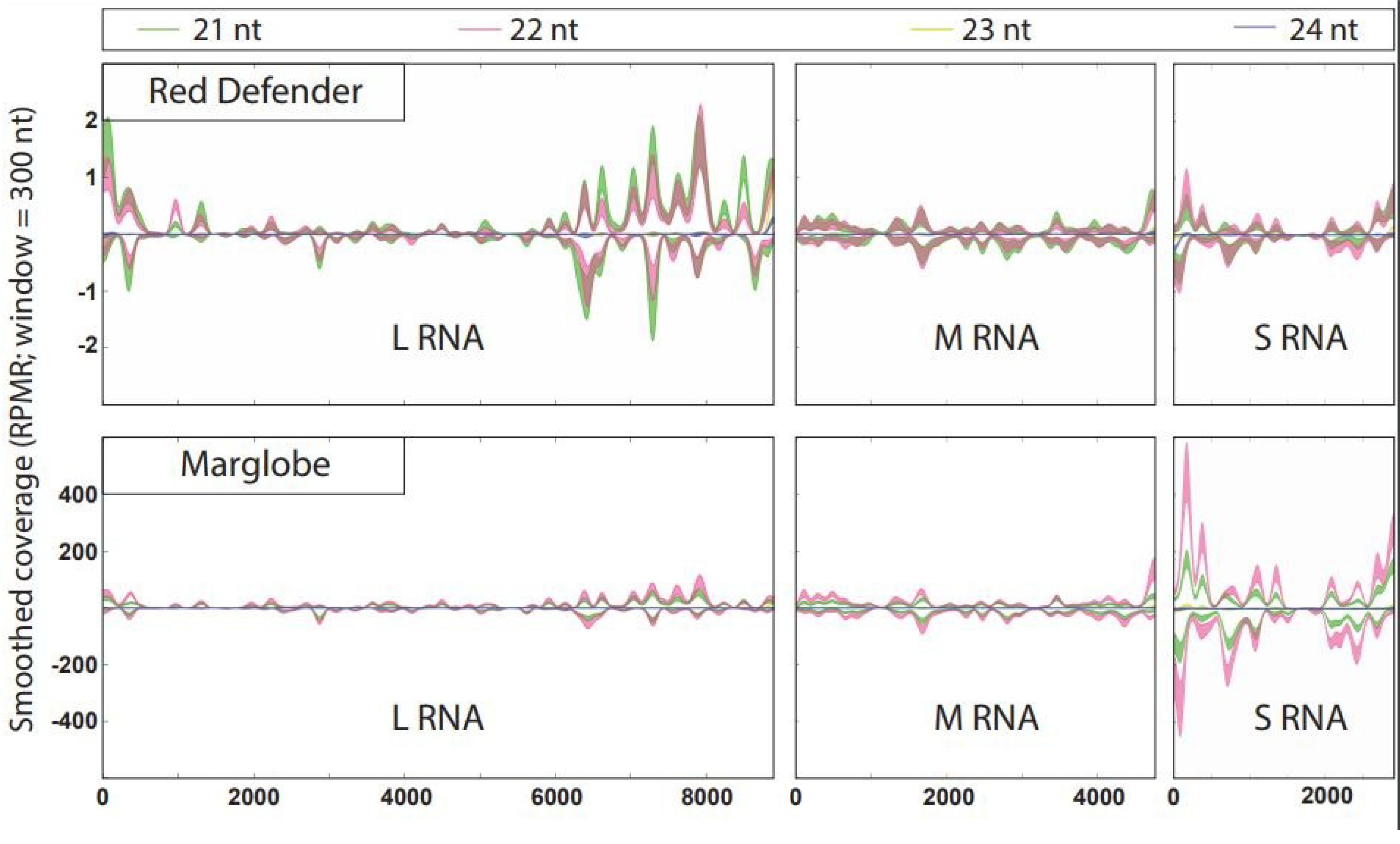
3.3. Hotspots in TSWV RNAs
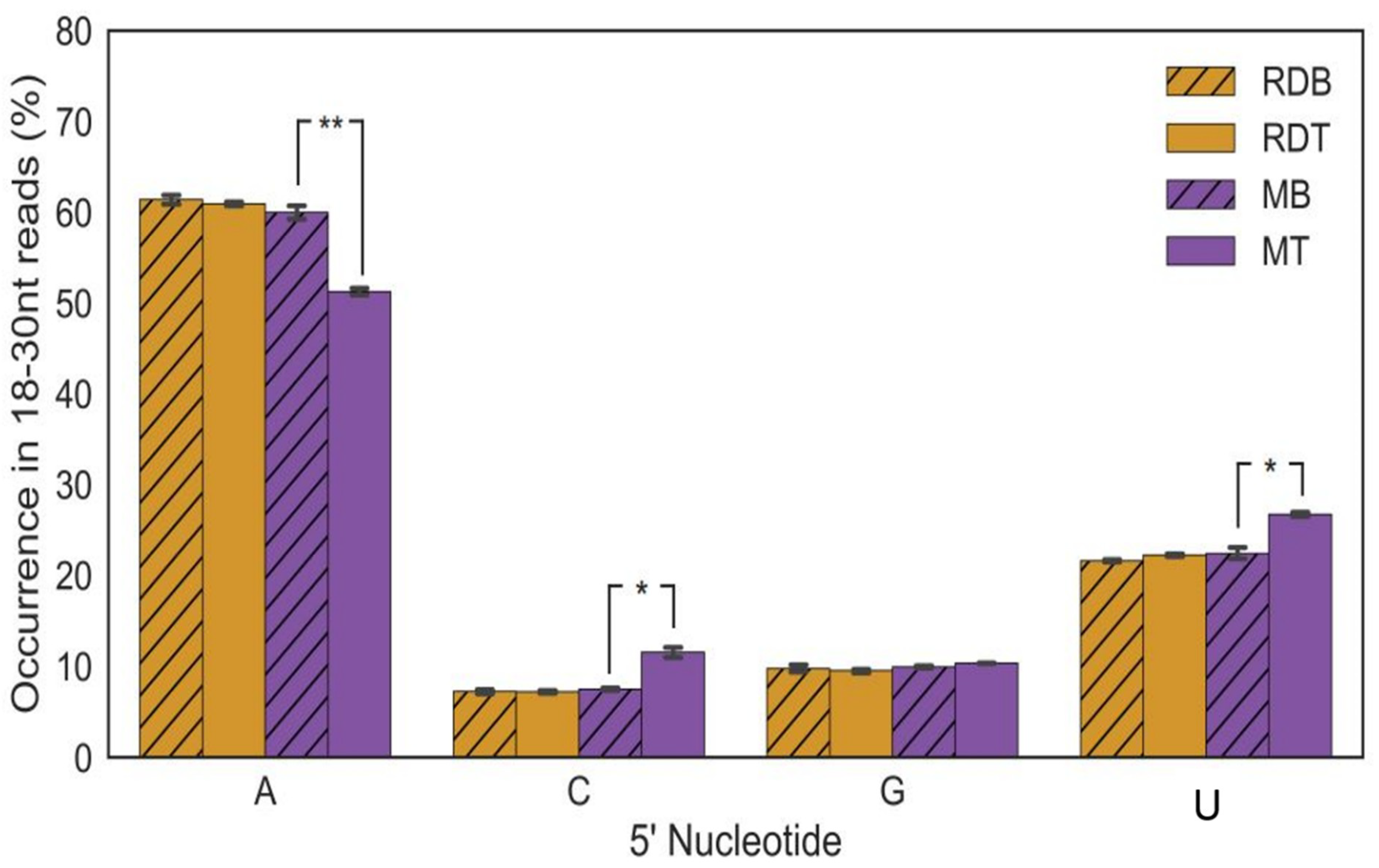
3.4. siRNAs 5′ Terminal Nucleotide-Strand Polarity
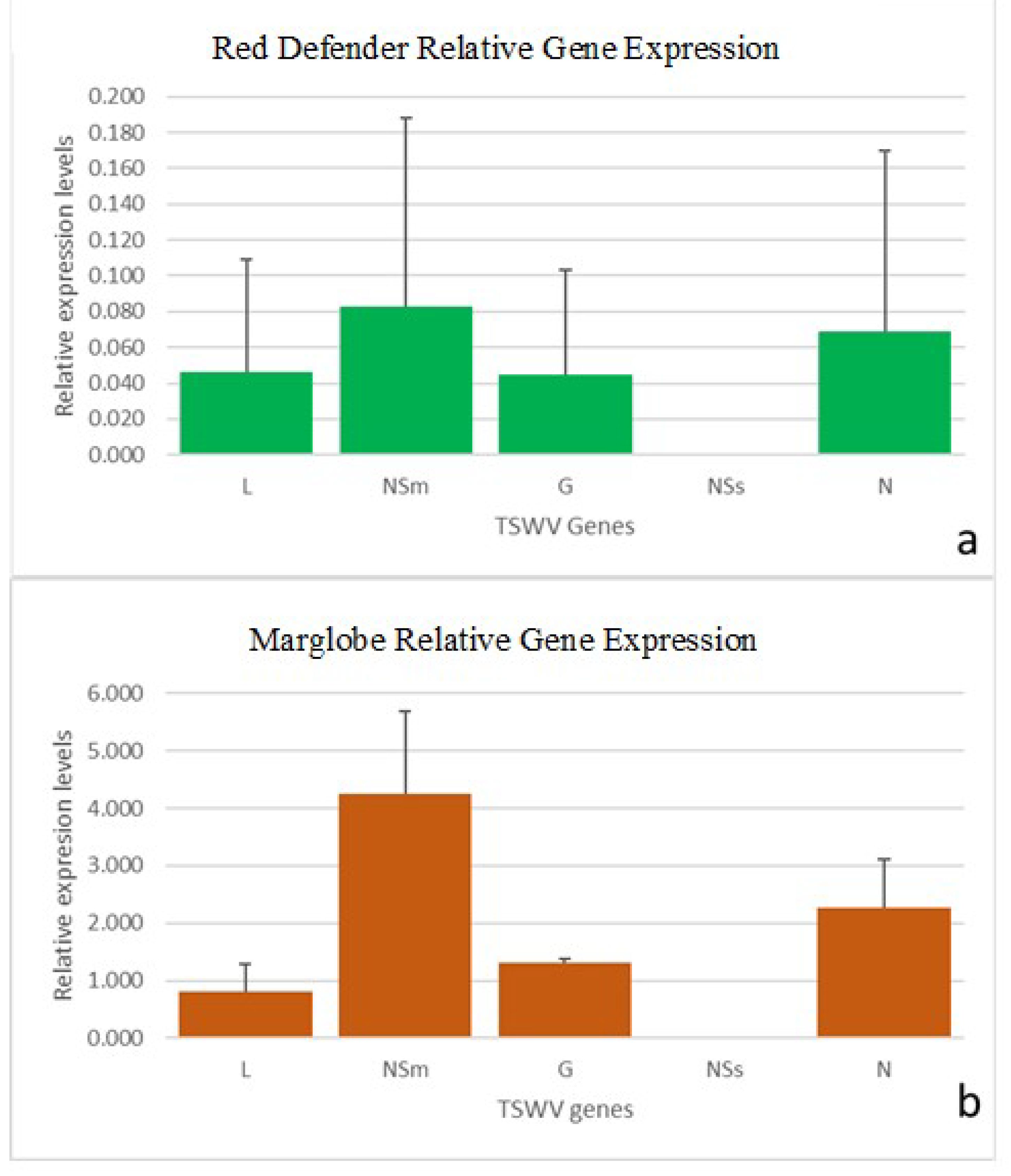
3.5. RT-qPCR of TSWV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Alkhovsky, S.V.; Bào, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotenberg, D.; Whitfield, A.E. Molecular interactions between Tospoviruses and Thrips vectors. Curr. Opin. Virol. 2018, 33, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Resende, R.; Pappu, H.R. Encylopedia of Virology, 4th ed.; Orthotospoviruses (Tospoviridae); Zuckeman, M., Bamford, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; in press. [Google Scholar]
- Pappu, H.R.; Whitfield, A.E.; de Oliveira, A.S. Encylopedia of Virology, 4th ed.; Tomato Spotted Wilt Virus (Tospoviridae); Zuckeman, M., Bamford, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; in press. [Google Scholar]
- Oliver, J.E.; Whitfield, A.E. The Genus Tospovirus: Emerging Bunyaviruses that Threaten Food Security. Annu. Rev. Virol. 2016, 3, 101–124. [Google Scholar] [CrossRef]
- Pappu, H.R.; Jones, R.A.C.; Jain, R.K. Global status of Tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 1412, 219–236. [Google Scholar] [CrossRef]
- Walter, C.T.; Barr, J.N. Recent advances in the molecular and cellular biology of bunyaviruses. J. Gen. Virol. 2011, 92, 2467–2484. [Google Scholar] [CrossRef]
- Kormelink, R.J.M. The Bunyaviridae: Molecular and Cellular Biology; The Molecular Biology of Tospoviruses and Resistance Strategies; Elliott, R.M., Plyusin, A., Eds.; Caister Academic Press: Norfolk, UK, 2011; pp. 163–191. [Google Scholar]
- Prins, M.; Goldbach, R. The emerging problem of Tospovirus infection and nonconventional methods of control. Trends Microbiol. 1998, 6, 31–35. [Google Scholar] [CrossRef]
- Stevens, M.R.; Scott, S.J.; Gergerich, R.C. Inheritance of a gene for resistance to Tomato spotted wilt virus (TSWV) from Lycopersicon peruvianum Mill. Euphytica 1991, 59, 9–17. [Google Scholar] [CrossRef]
- Dianese, É.C.; Fonseca, M.E.N.; Inoue-Nagata, A.K.; Resende, R.O.; Boiteux, L.S. Search in Solanum (section Lycopersicon) germplasm for sources of broad-spectrum resistance to four Tospovirus species. Euphytica 2011, 180, 307–319. [Google Scholar] [CrossRef]
- De Oliveira, A.S.; Boiteux, L.S.; Kormelink, R.; Resende, R.O. The Sw-5 Gene Cluster: Tomato Breeding and Research Toward Orthotospovirus Disease Control. Front. Plant Sci. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- De Oliveira, A.S.; Koolhaas, I.; Boiteux, L.S.; Caldararu, O.F.; Petrescu, A.J.; Resende, R.O.; Kormelink, R. Cell death triggering and effector recognition by Sw-5 SD-CNL proteins from resistant and susceptible tomato isolines to Tomato spotted wilt virus. Mol. Plant Pathol. 2016, 17, 1442–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallwass, M.; de Oliveira, A.S.; de Campos Dianese, E.; Lohuis, D.; Boiteux, L.S.; Inoue-Nagata, A.K.; Resende, R.O.; Kormelink, R. The Tomato spotted wilt virus cell-to-cell movement protein (NSM ) triggers a hypersensitive response in Sw-5-containing resistant tomato lines and in Nicotiana benthamiana transformed with the functional Sw-5b resistance gene copy. Mol. Plant Pathol. 2014, 15, 871–880. [Google Scholar] [CrossRef]
- Hoffmann, K.; Qiu, W.P.; Moyer, J.W. Overcoming Host- and Pathogen-Mediated Resistance in Tomato and Tobacco Maps to the M RNA of Tomato spotted wilt virus. Mol. Plant Microbe Interact. 2001, 14, 242–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Goldbach, R.; Bucher, E.; Prins, M. Resistance mechanisms to plant viruses: An overview. Virus Res. 2003, 92, 207–212. [Google Scholar] [CrossRef]
- Donaire, L.; Barajas, D.; Martinez-Garcia, B.; Martinez-Priego, L.; Pagan, I.; Llave, C. Structural and Genetic Requirements for the Biogenesis of Tobacco rattle virus-Derived Small Interfering RNAs. J. Virol. 2008, 82, 5167–5177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenllado, F.; Llave, C.; Díaz-Ruíz, J.R. RNA interference as a new biotechnological tool for the control of virus diseases in plants. Virus Res. 2004, 102, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small rnas. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.M.; Pruss, G.J.; Vance, V.B. Platn viral suppressors of RNA silencing. Virus Res. 2004, 102, 97–108. [Google Scholar] [CrossRef]
- Wang, M.B.; Masuta, C.; Smith, N.A.; Shimura, H. RNA Silencing and Plant Viral Diseases. Mol. Plant Microbe Interact. 2012, 25, 1275–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wu, Z.; Li, Y.; Wu, J. Biogenesis, function, and applications of virus-derived small RNAs in plants. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, C.; Frizzi, A.; Kao, J.; Jia, L.; Huang, M.; Zhang, Y.; Huang, S. Using small RNA sequences to diagnose, sequence, and investigate the infectivity characteristics of vegetable-infecting viruses. Arch. Virol. 2011, 156, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Hedil, M.; Hassani-Mehraban, A.; Lohuis, D.; Kormelink, R. Analysis of the A-U rich hairpin from the intergenic region of Tospovirus S RNA as target and inducer of RNA silencing. PLoS ONE 2014, 9, e106027. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Miozzi, .L.; Rosa, C.; Axtell, M.J.; Pappu, H.R.; Turina, M. Small RNA profiles of wild-type and silencing suppressor-deficient Tomato spotted wilt virus infected Nicotiana benthamiana. Virus Res. 2015, 208, 30–38. [Google Scholar] [CrossRef]
- Mitter, N.; Koundal, V.; Williams, S.; Pappu, H.R. Differential expression of Tomato spotted wilt virus-derived viral small rnas in infected commercial and experimental host plants. PLoS ONE 2013, 8, e76276. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Shrestha, A.; Peters, J.R.; Carroll, B.J.; Srinivasan, R.; Pappu, H.R.; Mitter, N. The Tomato spotted wilt virus genome is processed differentially in its plant host arachis hypogaea and its thrips vector Frankliniella fusca. Front. Plant Sci. 2016, 7, 1349. [Google Scholar] [CrossRef] [Green Version]
- Margaria, P.; Miozzi, L.; Ciuffo, M.; Pappu, H.R.; Turina, M. The first complete genome sequences of two distinct European Tomato spotted wilt virus isolates. Arch. Virol. 2014, 160, 591–595. [Google Scholar] [CrossRef]
- Margaria, P.; Miozzi, L.; Ciuffo, M.; Rosa, C.; Axtell, M.J.; Pappu, H.R.; Turina, M. Comparison of small RNA profiles in Nicotiana benthamiana and Solanum lycopersicum infected by polygonum ringspot Tospovirus reveals host-specific responses to viral infection. Virus Res. 2016, 211, 38–45. [Google Scholar] [CrossRef]
- Naveed, K.; Mitter, N.; Harper, A.; Dhingra, A.; Pappu, H.R. Comparative analysis of virus-specific small RNA profiles of three biologically distinct strains of Potato virus Y in infected potato (Solanum tuberosum) cv. Russet Burbank. Virus Res. 2014, 191, 153–160. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Williams, S.; Kappagantu, M.; Mitter, N.; Pappu, H.R. Transcriptome-wide identification of host genes targeted by Tomato spotted wilt virus-derived small interfering RNAs. Virus Res. 2017, 238, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S.J.; Boden, M.; Mitter, N.; Carroll, B.J. SCRAM: A pipeline for fast index-free small RNA read alignment and visualization. Bioinformatics 2018, 34, 2670–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Educ. 2011, 39, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinforma. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotenberg, D.; Krishna Kumar, N.K.; Ullman, D.E.; Montero-Astúa, M.; Willis, D.K.; German, T.L.; Whitfield, A.E. Variation in Tomato spotted wilt virus titer in Frankliniella occidentalis and its association with frequency of transmission. Phytopathology 2009, 99, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badillo-Vargas, I.E.; Rotenberg, D.; Schneweis, D.J.; Hiromasa, Y.; Tomich, J.M.; Whitfield, A.E. Proteomic Analysis of Frankliniella occidentalis and differentially expressed proteins in response to Tomato spotted wilt virus infection. J. Virol. 2012, 86, 8793–8809. [Google Scholar] [CrossRef] [Green Version]
- Mascia, T.; Santovito, E.; Gallitelli, D.; Cillo, F. Evaluation of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in infected tomato plants. Mol. Plant Pathol. 2010, 11, 805–816. [Google Scholar] [CrossRef]
- Nachappa, P.; Margolies, D.C.; Nechols, J.R.; Whitfield, A.E.; Rotenberg, D. Tomato Spotted wilt virus benefits a non-vector arthropod, tetranychus urticae, by modulating different plant responses in tomato. PLoS ONE 2013, 8, e75909. [Google Scholar] [CrossRef] [Green Version]
- Bouché, N.; Lauressergues, D.; Gasciolli, V.; Vaucheret, H. An antagonistic function for Arabidopsis DCL2 in development and a new function for DCL4 in generating viral siRNAs. EMBO J. 2006, 25, 3347–3356. [Google Scholar] [CrossRef] [Green Version]
- Donaire, L.; Wang, Y.; Gonzalez-Ibeas, D.; Mayer, K.F.; Aranda, M.A.; Llave, C. Deep-sequencing of plant viral small RNAs reveals effective and widespread targeting of viral genomes. Virology 2009, 392, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.F.; Romanel, E.A.; Andrade, R.R.; Farinelli, L.; Østerås, M.; Deluen, C.; Corrêa, R.L.; Schrago, C.E.; Vaslin, M.F. Profile of small interfering RNAs from cotton plants infected with the polerovirus Cotton leafroll dwarf virus. BMC Mol. Biol. 2011, 12, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargason, J.M.; Szittya, G.; Burgyán, J.; Tanaka Hall, T.M. Size selective recognition of sirna by an rna silencing suppressor. Cell 2003, 115, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Ogwok, E.; Ilyas, M.; Alicai, T.; Rey, M.E.C.; Taylor, N.J. Comparative analysis of virus-derived small RNAs within cassava (Manihot esculenta Crantz) infected with cassava brown streak viruses. Virus Res. 2016, 215, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusaro, A.F.; Matthew, L.; Smith, N.A.; Curtin, S.J.; Dedic-Hagan, J.; Ellacott, G.A.; Watson, J.M.; Wang, M.B.; Brosnan, C.; Carroll, B.J.; et al. RNA interference-inducing hairpin RNAs in plants act through the viral defence pathway. EMBO Rep. 2006, 7, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhang, H.; Adams, M.J.; Yang, J.; Peng, J.; Antoniw, J.F.; Zhou, Y.; Chen, J. Characterization of siRNAs derived from rice stripe virus in infected rice plants by deep sequencing. Arch. Virol. 2010, 155, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Yang, G.S.; Chen, W.T.; Lin, R.M.; Ling, J.; Mao, Z.C.; Xie, B.Y. Characterization and function of Tomato yellow leaf curl virus-derived small RNAs generated in tolerant and susceptible tomato varieties. J. Integr. Agric. 2016, 15, 1785–1797. [Google Scholar] [CrossRef] [Green Version]
- Spanò, R.; Mascia, T.; Kormelink, R.; Gallitelli, D. Grafting on a non-transgenic tolerant tomato variety confers resistance to the infection of a sw5-breaking strain of Tomato spotted wilt virus via RNA silencing. PLoS ONE 2015, 10, e0141319. [Google Scholar] [CrossRef] [Green Version]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R.; Baulcombe, D.C. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2010, 328, 872–875. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sw-5(+) Mock | Sw-5(+) TSWV | Sw-5(−) Mock | Sw-5(−) TSWV | |
|---|---|---|---|---|
| Total sequences * | 30,796,029.33 | 29,959,930.67 | 29,738,796.33 | 29,284,469.33 |
| Total Nucleotides in data set * | 701,391,986.7 | 682,215,963.3 | 674,277,615.3 | 652,956,838.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olaya, C.; Fletcher, S.J.; Zhai, Y.; Peters, J.; Margaria, P.; Winter, S.; Mitter, N.; Pappu, H.R. The Tomato spotted wilt virus (TSWV) Genome is Differentially Targeted in TSWV-Infected Tomato (Solanum lycopersicum) with or without Sw-5 Gene. Viruses 2020, 12, 363. https://doi.org/10.3390/v12040363
Olaya C, Fletcher SJ, Zhai Y, Peters J, Margaria P, Winter S, Mitter N, Pappu HR. The Tomato spotted wilt virus (TSWV) Genome is Differentially Targeted in TSWV-Infected Tomato (Solanum lycopersicum) with or without Sw-5 Gene. Viruses. 2020; 12(4):363. https://doi.org/10.3390/v12040363
Chicago/Turabian StyleOlaya, Cristian, Stephen J. Fletcher, Ying Zhai, Jonathan Peters, Paolo Margaria, Stephan Winter, Neena Mitter, and Hanu R. Pappu. 2020. "The Tomato spotted wilt virus (TSWV) Genome is Differentially Targeted in TSWV-Infected Tomato (Solanum lycopersicum) with or without Sw-5 Gene" Viruses 12, no. 4: 363. https://doi.org/10.3390/v12040363