
Abstract
Limited understanding of the effects of enhanced nitrogen (N) addition and grazing exclusion (E) on greenhouse gases fluxes (GHGs: CO2, CH4, and N2O) in grasslands constrains our ability to respond to the challenges of future climate change. In this study, we conducted a field experiment using a static closed opaque chamber to investigate the response of GHG fluxes to N addition (69 kg N ha−1 year−1 applied in 3 splits) and grazing exclusion in an alpine meadow on the Tibetan Plateau during the growing seasons from 2011 to 2013. Our results showed that winter grazing significantly raised soil temperature (ST), while grazing exclusion (E) had no effect on soil moisture (SM), and N fertilizer (F) had no effect on ST or SM. Aboveground biomass (AB) and root biomass (RB) were not significantly affected by E in 2011-2013 (p > 0.05), but F significantly affected AB and RB (p < 0.05). Compared with winter grazing, only E substantially reduced seasonal mean CO2 emissions (by about 20.1%) during the experimental period. E did not significantly directly affect CH4 uptake, whereas N addition reduced seasonal mean CH4 uptake by about 6.5%, and N addition changed seasonal average absorption of N2O into an emission source. CO2 flux is the major contributor to CO2 equivalent emissions in this area. Our results indicate that exclosure from livestock grazing might be a promising measure to reduce CO2 emissions, while enhanced N addition might reduce CH4 uptake and increase N2O emission in the alpine meadow under future climate change.
Similar content being viewed by others

Introduction
Increases in atmospheric nitrogen (N) deposition (Ndep) from human activities can strongly affect the exchange of greenhouse gases (GHG, i.e. CO2, CH4, and N2O) between terrestrial ecosystems and the atmosphere (Reay et al. 2008; Vitousek et al. 1997). Since the Industrial Revolution, the emission of reactive N into the atmosphere is estimated to have increased by nearly 11.47 times, from 15 Tg N year−1 in 1860 to 187 Tg N year−1 in 2005, and is expected to reach 200 Tg N year−1 by 2050 (Galloway et al. 2008). After America and Europe, China now has the third highest rates of N deposition, with significant increases also found on the Tibetan Plateau, and rates of N deposition are expected to continue to increase in the coming decades due to economic development (Galloway and Cowling 2002; Liu et al. 2013a, b). Meanwhile, some alpine grasslands on the Tibetan Plateau have become degraded in recent decades due to overgrazing driven by population growth and food demand (Wei et al. 2012). Exclosure from livestock grazing has been widely employed by China’s national and local authorities as a management practice aimed at restoring and protecting these fragile ecosystems (Luo et al. 2015; Zhang et al. 2012; Zou et al. 2014). Although some studies have explored the separate effects of Ndep and grazing exclusion on GHG fluxes (Chen et al. 2016; Hu et al. 2017; Liu et al. 2013a, b; Zhang et al. 2012), research on their combined effects on GHG fluxes in alpine meadows is scarce.
Previous studies have shown that compared with grazed plots, grazing exclusion significantly decreased soil respiration (Rs) by 23.6% over the growing season and by 21.4% annually, but grazing exclusion increased the temperature sensitivity (Q10) of seasonal and annual soil respiration by 6.5% and 14.2%, respectively (Chen et al. 2016). Some studies in the region suggest that grazing exclusion and N fertilization have no effect on CO2 emissions (Hu et al. 2017; Zhang et al. 2012), but that they can significantly reduce average CH4 uptake (Hu et al. 2017; Jiang et al. 2010). Grazing exclusion has little effect on N2O emissions (Hu et al. 2017), whereas N fertilizer significantly increases average N2O emission (Jiang et al. 2010). These inconsistent and even contradictory responses of GHG fluxes to Ndep and grazing exclusion could be due to different mechanisms. For example, Ndep directly increases soil N availability, which promotes plant productivity (Bala et al. 2013; LeBauer and Treseder 2008) and thus increases inputs of nutrients into soils through decomposition from increased root and litter biomass. However, grazing exclusion decreases soil temperature (Luo et al. 2010) and increases plant productivity through shifts in plant composition towards tall grasses with deep roots (Xiong et al. 2016; Zhang et al. 2015; Zhu et al. 2016b). Therefore, the lack of data on GHG fluxes under different Ndep rates combined with grazing exclosure limits our mechanistic understanding of the relationship between GHG fluxes and Ndep, rendering predictions of the net strength of grassland GHG sinks in the future highly uncertain. We hypothesized that (1) N fertilization and grazing exclosure would increase CO2 due to increased plant production; (2) N fertilization would increase N2O emission, but grazing exclosure would decrease it; and (3) N fertilization would decrease CH4 uptake due to changes in soil inorganic N, whereas grazing exclosure would increase CH4 uptake due to decreased soil compaction. Here, measurements and analysis of GHG fluxes were carried out in grazed and fenced areas of an alpine meadow on the Tibetan Plateau during the growing seasons from 2011 to 2013. The aims of the study were to (1) investigate the effects of grazing exclusion and experimental N addition on key GHG fluxes, and (2) identify which abiotic environmental factors (i.e., soil temperature and moisture) drive changes in GHG fluxes in the alpine region.
Materials and methods
Study site
This study was carried out at an alpine meadow at Haibei Alpine Meadow Ecosystem Research Station, Northwest Plateau Institute of Biology, Chinese Academy of Sciences (37° 36′ N, 101° 12′ E, and 3250 m above sea level) in Qinghai Province during the growing seasons (from June to September) of 2011–2013. The local climate is characterized by long, cold winters and short mild summers, with multi-year mean annual precipitation of 580 mm (from 1981 to 2012), of which 80% is concentrated in the growing season from June to September. Mean annual air temperature is 1.7 °C with monthly mean air temperature ranging from − 15 °C in January to 10 °C in July. The plant community of the natural alpine meadow at the experimental site is dominated by grasses and forbs, Lancea tibetica, Pedicularis Linn., Gentianopsis paludosa, Potentilla anserina L., Aster yunnanensis Franch., Gentiana straminea and Saussurea pulchra. Prior to the experiment, the experiment site was freely grazed by sheep during winter (from the end of September to May of the following year) each year with almost no litter biomass remaining aboveground. The soil has a clay-loam texture with an average depth of 0.65 m, and is classified as Mat-Gryic Cambisol (Chinese Soil Taxonomy Research Group 1995), corresponding to Gelic Cambisol (WRB 2006). Basic soil properties were as follows: total organic carbon (C) 55.8 g kg−1, potassium (K) 13.0 g kg−1, phosphorus (P) 0.70 g kg−1, nitrogen (N) 5.37 g kg−1, pH of 8.2 determined in distilled water, and bulk density in the 0-10 cm soil layer is 1.05 g cm−3 (Zhang et al. 2012).
Experiment design
The experiment site was surrounded by eight fenced squares of 4 × 4.5 m. Before exclusion of livestock from the freely-grazed meadow, vegetation and other environmental conditions inside and outside the fence were almost homogeneous. Outside of the fenced meadow, grassland was still freely grazed by local herders’ animals during winter, i.e. from the end of September to the end of May the following year. Eight plots (each of 4 m × 4.5 m dimension) were assigned to two treatments (i.e., N addition and control) with four replications inside the fence and four replications outside the fence. The treatments were laid out randomly. A completely randomized design was used for two exclosures and their fertilizer management practices with four replicate plots (4 m × 4.5 m) of four treatments as follows: grazing without N addition (G+-N0), grazing with N addition (G+-N+), no-grazing without N addition (G0-N0) and no-grazing with N addition (G0-N+). Local atmospheric N deposition is estimated to range from 8.7 to 13.8 kg N ha−1 year−1 (Lü and Tian 2007). The N fertilization experiment was initiated in May 2011 and the simulated future climate change scenario used about six times the amount of N deposition. N fertilization (69 kg N ha−1 year−1 as urea applied three times) was added on 20 June, 20 July and 20 August each year in 2011, 2012 and 2013, respectively.
Field sampling and measurements
GHG fluxes were measured using a static dark chamber and the gas chromatography method. A stainless steel square box [without a top or bottom, 0.4 m (length) × 0.4 m (width) × 0.08 m (height)] with a water groove to make the chamber airtight was placed 0.20 m away from the edge of each plot and inserted into the soil to a depth of 0.08 m. Four air samples were collected from 09:00 to 11:00 a.m., representing 1-day average flux as described in previous reports (Jiang et al. 2010; Lin et al. 2009). The gas samples were collected every 7–10 days from June to September during the growing seasons in 2011, 2012, and 2013. GHG fluxes were monitored on the first or second day after fertilizer application, and then monitored with the same frequency as prior to fertilization. For all observations, average gas fluxes and standard errors were calculated from four replicates. Seasonal GHG fluxes in 2011, 2012 and 2013 were estimated by calculating average fluxes over an experimental period with 16 flux observations in each period.
Air temperature (1.5 m above ground) data were recorded simultaneously using an automated weather station (AWS, Vaisala Corp., Holland), and precipitation was measured using a rain gauge. Soil temperature and soil water content data (0.05 m) were measured simultaneously to the air sampling process at each chamber using a digital temperature sensor (JM624 digital thermometer, Living–Jinming Ltd., China) and a Time Domain Reflectometer (JS-TDR300, Meridian Measurement, USA).
Peak aboveground biomass was estimated by clipping vegetation to the soil surface using a 0.5 × 0.5 m quadrat placed 0.2 m away from the plot edge in each plot in late August each year. At the center of each quadrat, two soil cores of 0–0.2 m depth were collected using a 0.08 m diameter soil auger. All samples with four replicates were taken on the same day. Samples of soil cores were washed in the laboratory with tap water to remove the soil so as to estimate root biomass. All samples were oven-dried at 65 °C to a constant weight.
CO2 equivalent emissions
IPCC (2007) estimated that N2O and CH4 have global warming potentials that are 296 and 23 times higher than CO2, respectively, and these values were used in the conversion of N2O and CH4 to CO2 equivalent warming potential after addition of CO2 emissions. The formula used was as follows:
Statistical analysis
General Linear Model (GLM) Repeated Measures Analysis of Variance (RMANOVA), with exclusion as the main factor (between-subject) and sampling date as the within-subject factor including interactions, was applied to test the effects of the main factor on monthly soil temperature, monthly soil water content, daily GHG fluxes and annual average GHG fluxes. The same analysis was performed using N fertilization as the main factor under the same treatment and with sampling date as the within-subject factor, including interactions. Multi-comparison of least standard difference (LSD) was conducted for all measured variables within each sampling date using one-way ANOVA. In order to assess how N fertilizer affected the magnitude of the effect of exclosure on GHG flues, soil temperature and moisture, aboveground and root biomass, multi-comparison of least standard difference (LSD) was conducted using one-way ANOVA to assess the relative differences in GHG fluxes, soil temperature and moisture, aboveground and root biomass under no-fertilizer and with-fertilizer management practices. All statistical analyses were performed with SPSS (SPSS 16.0, SPSS Inc., Chicago, IL, USA) using the GLM procedure and Type III sum of squares. We further fitted a piecewise structural equation model (SEM) to estimate the direct and indirect effects of grazing exclosure (E) and nitrogen fertilizer (F) on seasonal cumulative CO2 emission, CH4 uptake and N2O emission using the piecewise SEM package in R software (R 3.1.3, R Development Core Team 2014).
Results
Environmental conditions
Air temperature showed a typical pattern that increased from June, peaked in August, and declined in September in each of the 3 years (Fig. S1). Mean air temperature and total rainfall during the growing seasons from 1 June to 30 September in 2011, 2012, and 2013 were 8.9, 9.0, and 9.6 °C, and 326.8, 285.6, 356.0 mm, respectively. The seasonal rainfall distributions and temperatures are shown in Fig. S1.
Soil temperature and soil moisture
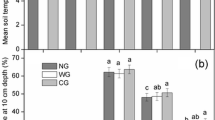
The effect of exclusion on soil temperature (ST) and soil moisture (SM) varied with sampling date and year, and there were significant interactions between exclusion and year and sampling date (p < 0.05). In general, ST under grazing was significantly higher than under exclusion (p < 0.001), but exclusion had no effect on SM (p = 0.21), while fertilization (F) had no significant effect on ST or SM (p > 0.05). Average annual ST (Fig. 1a) was greater for G+-N0 and G+-N+, and the annual SM of G0-N0 and G0-N+ were significantly lower than for G+-N0 and G+-N+ in 2011, but there were no significant differences in 2012 or 2013 (Fig. 1b). In general, N fertilizer did not significantly affect ST or SM for G+-N0 and G0-N0 during the 3 year experiment (Fig. 1a, b).

Seasonal average soil temperature (a) and soil moisture (b) at 5 cm depth under four treatments from 2011 to 2013. G+: grazing; N+: nitrogen fertilizer; G0: no-grazing; N0: without N addition. Bars indicate mean ± 1SE. Different letters indicate significant differences at p = 0.05 level
Aboveground and root biomass
Although aboveground biomass (AB) and root biomass (RB) were not significantly affected by exclosure (E) in 2011–2013 (p > 0.05), nitrogen fertilizer (F) significantly affected AB and RB (p < 0.05). There were no interaction effects between E and F on AB or RB (p > 0.05). Inter-annual variation in AB and RB was not significant (p > 0.05). In general, AB in the G+-N0 treatment was lower than in the G+-N+, G0-N0 and G0-N+ treatments, and RB in the G+-N0 treatment was higher than in the G+-N+ and G0-N+ treatments in 2011–2013 (Fig. 2).

Average aboveground biomass (AB) and root biomass (RB) under four treatments from 2011 to 2013. G+: grazing; N+: nitrogen fertilizer; G0: no-grazing; N0: without N addition. Bars indicate mean ± 1SE. Different letters indicate significant differences at p = 0.05 level
GHG fluxes
CO2 emission followed a clear seasonal pattern with the highest rates in the vigorous growth period and the lowest rates in the early and late growing seasons in each of the 3 years (Fig. 3; Table 1). The highest value appeared in mid-July to mid-August. Regardless of fertilization, compared with the native alpine meadow, grazing exclusion reduced CO2 emissions by an average of 20.1% over the 3-year period, whereas N addition had no significant effect on CO2 emissions (Tables 1, 2). There were no significant interactive effects between N addition and grazing exclusion (F = 0.022, p = 0.887, Table 1). Considering the strong collinearity among affecting factors, we fitted a piecewise structural equation model (SEM) to evaluate the causal relationships among these factors in order to estimate the direct and indirect effects of exclosure and nitrogen fertilizer on cumulative seasonal CO2 emission, CH4 uptake and N2O emission (Fig. 6a–c). The influence of exclosure on seasonal CO2 emission was mainly mediated through soil temperature. There was a strong relationship between exclusion and RB (β = − 0.63, standardized coefficient), and although there were significant relationships between N fertilizer and AB and RB (Fig. 6a), there were weaker relationships between AB, RB and CO2 emissions. Grazing exclusion led to significant change in seasonal average CO2 emission in 2011, 2012 and 2013 (Fig. 6a; Table 1).

Daily CO2 flux under four treatments in 2011 (a), 2012 (b) and 2013 (c). G+: grazing; N+: nitrogen fertilizer; G0: no-grazing; N0: without N addition. Bars indicate mean ± 1SE. Asterisk indicates significant difference between treatments at p < 0.05 level. Mean ± SE (n = 4) are shown in the figure. Arrow indicates the date of N fertilization
The seasonal dynamics of CH4 were very significant (Fig. 4; Table 1). Grazing exclusion (E) had no significant effect on CH4 uptake (Fig. 6b; Table 1), whereas N addition reduced CH4 uptake (Table 2). There were no significant interactive effects on CH4 between N addition and grazing exclusion (F = 0.191, p = 0.673, Table 1). The alpine meadow soil was a CH4 sink in each growing season (Fig. 4; Table 2). There were strong relationships between CH4 uptake and soil moisture, soil moisture and soil temperature, exclusion and soil temperature and weaker relationships between CH4 uptake and soil temperature, CH4 uptake and exclusion (Fig. 6b). However, nitrogen fertilizer had a significant effect on CH4 uptake (Fig. 6b).

Daily CH4 flux under G+-N0, G0-N0 and nitrogen fertilizer in 2011 (a), 2012 (b) and 2013 (c). G+: grazing; N+: nitrogen fertilizer; G0: no-grazing; N0: without N addition. Bars indicate mean ± 1SE. Asterisk indicates significant difference between treatments at p < 0.05 level. Mean ± SE (n = 4) are shown in the figure. Arrow indicates the date of N fertilization
The alpine meadow soil acted as a weak N2O source in the growing seasons under the G+-N+ and G0-N+ treatments, but was a weak N2O sink in the growing seasons for G+-N0 and G0-N0 (Table 2). Exclusion had no effect on N2O exchange capacity (Fig. 6c; Table 1). However, N2O exchange was converted from absorption to emission by N fertilizer application (Table 2). The relationship between N2O and nitrogen fertilizer was very weak (Fig. 6b), but the combination of all direct and indirect effects was significant (Table 1). We detected significant seasonal dynamics in N2O emissions (Fig. 5; Table 1), which was highly variable. N addition significantly increased N2O release (p < 0.001), resulting in net N2O emissions (Fig. 6c). The strongest relationships were between N2O emission and soil moisture, soil moisture and soil temperature, exclosure and soil temperature, and aboveground biomass and nitrogen fertilizer (Fig. 6c), and there were weaker relationships between N2O emission and soil temperature, and exclusion and aboveground biomass. Exclusion affects SM by affecting ST, and indirectly affects emissions of N2O (Fig. 6c).

Daily N2O flux under the G+-N0, G0-N0 and nitrogen fertilizer treatments in 2011 (a), 2012 (b) and 2013 (c). G+: grazing; N+: nitrogen fertilizer; G0: no-grazing; N0: without N addition. Bars indicate mean ± 1SE. Asterisk indicates significant difference between treatments at p < 0.05 level. Mean ± SE (n = 4) are shown in the figure. Arrow indicates the date of N fertilization

A piecewise structural equation model fitted to infer the direct and indirect effects of grazing exclusion (E) and nitrogen fertilizer (F), soil temperature (ST) and moisture (SM), aboveground biomass (AB) and root biomass (RB) on seasonal average CO2, CH4 and N2O emission from 2011 to 2013. Arrows represent the direction of causality. The bold black lines represent highly significant relationships [p < 0.001, three asterisks (***)]; the non-bold black lines represent significant relationships [p < 0.05, two asterisks (**)]; and the dotted lines represent non-significant relationships (p > 0.05). AB and RB are not shown in figure b because the result cannot be run after adding these variables
In general, grazing exclusion had a significant impact on CO2 equivalent emissions, while N fertilizer had no effect on CO2 equivalent emissions (Table 2). The CO2 equivalent emissions of grazing without N addition and grazing with N addition were significantly higher than in the no-grazing without N addition and no-grazing with N addition treatments, with a 3-year average increase of about 20%. CO2 flux is the major contributor to CO2 equivalent emissions. Compared with CO2 flux, CH4 and N2O emissions were very small, so CO2 flux is the decisive factor in determining CO2 equivalent emissions.
Discussion
CO2 emission
Although positive effects of N fertilizer application on CO2 emissions were reported by Juutinen et al. (2010), we found that N addition did not alter CO2 fluxes under either grazing or no-grazing treatments in our 3-year study (Tables 1, 2). The first reason may be because the response of CO2 emissions to N addition is related to the amount of N addition (Carter et al. 2011). The annual N-addition rates in our study were lower than in some studies (Kostyanovsky et al. 2018), but higher than in other studies (Jiang et al. 2010; Wang et al. 2017; Zhu et al. 2015). N-addition rates may affect N-status associated processes because high N addition could remove N constraints on microbial metabolism and improve litter quality (Carter et al. 2012). Other studies have shown that the effects of N fertilizer application on CO2, CH4, and N2O emission are dependent on the form of N (Cai et al. 2007; Peng et al. 2011). Differences in soil structure and climate conditions between study sites may also have an effect, with the range of responses varying from no effect at all to positive or negative effects depending on ecosystem type, age, dominant plant species and soil chemical characteristics (Pregitzer et al. 2008; Rodriguez et al. 2014; Xiao 2017; Zhu et al. 2016a), N loads in the soil in combination with climatic conditions (Alster et al. 2013), and experiment duration (Zhou et al. 2014).
Grazing exclusion significantly decreased annual average CO2 emission (Table 2) in our study. This was largely because exclusion lowered soil temperature (Fig. 6a), which reduced soil respiration (Chen et al. 2016), and our study found that soil temperature was positively correlated with CO2 emissions (Fig. 6a). Studies conducted near our study site showed that grazing exclusion significantly decreased Rs, mainly because exclusion reduces soil temperature and microbial biomass carbon (Chen et al. 2016), but, as in our study, these effects are not correlated with exclusion-induced changes in aboveground and belowground biomass. Our results suggest that the effects of exclusion-induced changes in soil temperature on CO2 emission may have overridden the effects of exclusion-induced changes in plant productivity.
Grazing exclusion can have significant effects on an ecosystem’s biotic and abiotic characteristics, and these in turn can affect soil carbon fluxes (McSherry and Ritchie 2013). In the same site as the present study, Zou et al. (2016) found that accumulated litter was significantly higher under grazing exclusion (386.41 g m−2) than under grazing (58.77 g m−2), and because the low temperature caused by exclusion can slow down the litter decomposition rate (Luo et al. 2010), this further reduces CO2 emissions. Temperature (especially soil temperature) was the dominant environmental variable that controlled seasonal change in CO2 flux in our study, which has been documented in many other studies (Fang and Moncrieff 2001; Jiang et al. 2010; Lloyd and Taylor 1994; Wei et al. 2012; Zhu et al. 2015). Soil moisture is another important factor affecting CO2 emissions. In our study, CO2 fluxes were not positively correlated with soil moisture (Fig. 6a), probably because soil water content influenced microbial activity and litter decomposition rates and then had a combined influence on CO2 diffusion from the soil to the atmosphere.
CH4 uptake
Our results indicated that the natural grassland (exclosed, not exclosed, with or without N fertilizer) functioned as a sink for CH4 during the growing seasons from 2011 to 2013. This finding is consistent with other studies (Jiang et al. 2010; Wei et al. 2012). Low precipitation and soil moisture may have contributed to the stronger CH4 uptake in the alpine meadow (Curry 2007). In our study, CH4 emissions were not affected by N addition under winter grazing, but no grazing with N addition significantly reduced the absorption of CH4 (Table 2). This is likely direct influenced by the reduced activity of methanogens, as indicated by decreased CH4 production potential (Wang et al. 2017) for the no grazing with N addition treatment.
As nitro-bacteria compete for active sites of methane monooxygenase (MMO) when oxidizing NH4+ and CH4 via methane-oxidizing bacteria, the oxidation of CH4 is inhibited (Bowman et al. 1995). A higher concentration of NH4+ can inhibit the growth and activity of methane-oxidizing bacteria by increasing the number of nitrifying bacteria, thereby inhibiting the oxidation of CH4 (Jang et al. 2006). The soil NH4+ and NO3− content in different soil layers was similar between the fenced and grazed plots without N addition in our study site (Zou et al. 2016). This may be the reason for the lack of difference in CH4 emissions between the fenced and grazed plots without N addition (Table 2). Scheutz and Kjeldsen (2004) showed that high concentration of NH4+ (14 mg NH4+/kg) significantly inhibited the oxidation of CH4 (Scheutz and Kjeldsen 2004). Adamsen and King (1993) found in soil culture experiments that nitrate inhibited soil CH4 oxidation (Adamsen and King 1993).
Our results indicated that grazing reduces the absorption of CH4 (Table 2), which is similar to the results of previous studies (Liu et al. 2007; Saggar et al. 2007). This may be because trampling resulting in soil compaction, which can decrease O2 diffusion into the soil, thus limiting CH4 and O2 availability for the oxidation process (Liu et al. 2007; Saggar et al. 2007). However, Lin et al. (2015) found that summer grazing by sheep increased CH4 absorption in the alpine region, which may be due to the difference in grazing season between their experiment and ours. With winter grazing by sheep in our experiment, grass was almost completely consumed, and was also accompanied by a large amount of fecal/urine production. Yak dung application, however, significantly increased CH4 emissions, which was probably attributable to dissolved CH4, large microbial populations, highly degradable organic compounds and anaerobic conditions in the fresh dung patches (Jarvis et al. 1995; Lin et al. 2009; Sherlock et al. 2002), which lowered the absorption of methane. The study of Lin et al. (2015) simulated summer grazing (Wang et al. 2012), with half of canopy height consumed by herbivores and limited trampling or fecal urine, and soil temperature was raised by grazing (Luo et al. 2009). The increase in ST can increase evaporation of SM, and SM directly affects absorption of CH4 (Fig. 6b) and improves the permeability of soils and the activity of CH4 oxidizing bacteria, thus promoting the absorption of CH4 by soil (Hu et al. 2010; Zhuang et al. 2007). Hence, increased ST could offset the effects of grazing.
N2O flux
Our results indicated that exclusion of natural grassland from livestock grazing had no effect on N2O emission (Table 2) during the growing seasons from 2011 to 2013. This finding is different from studies that have shown that grazing may reduce N2O release (Liu et al. 2007; Wolf et al. 2010) or may increase it (Hu et al. 2010; Jiang et al. 2010). Both emission (positive values) and uptake of N2O (negative values) were observed across grazing exclusion and N fertilizer treatments, as has also been reported by other studies (Cantarel et al. 2011; Dijkstra et al. 2013; Jiang et al. 2010; Peng et al. 2017; Teh et al. 2014).
Previous studies showed that increased SM has a positive effect on N2O emission (Dijkstra et al. 2013; Hart 2006), while drought can lead to a reduction in N2O emission (Goldberg and Gebauer 2009; Larsen et al. 2011; Shi et al. 2012). Jiang et al. (2010) also reported for alpine meadow grassland that N2O emissions peaked under higher SM conditions. However, there was a significant correlation between soil moisture and N2O emission (Fig. 6c), suggesting that soil moisture is the main factor limiting N2O emission. None of the treatments significantly affected soil moisture in our study, except in 2011 (Fig. 1b), indicating that soil moisture is not the main factor causing differences in N2O emission between treatments. It is known that low temperature mainly influences microbial metabolic rates (Hulsen et al. 2016), which leads to the deterioration of nitrification (Chen et al. 2018; Delatolla et al. 2012; Hoang et al. 2014). In addition, low temperatures exert an influence on the amount of nitrifying bacteria (Siripong and Rittmann 2007). Low temperature is one of the key factors that shapes the microbial community in biological N removal systems (Urakawa et al. 2008; Zhou et al. 2016). However, there were no significant correlations between soil temperature and N2O emission in our study (Fig. 6c). Thus, there was an interactive effect on N2O emission between soil temperature and N addition because grazing increased soil temperature (Fig. 1a). The absence of increases in N2O emissions could be because denitrification was limited by low temperature (Ambus and Robertson 2006; Curtis et al. 2006; Liu and Greaver 2009) for the grazing exclosure with N addition treatment. In addition, the low temperature may have reduced nitrification by limiting aerobic ammonia oxidation due to the decreased abundance of ammonia-oxidizing archaea and bacteria (Zheng et al. 2014). Our results imply that N2O emission would increase if N addition increases with warming under future climate change. Moreover, we found that N addition significantly increased N2O emission under grazing and exclosure conditions during the experiment period (Tables 1, 2), a similar result to that of Wang et al. (2018) in the region (Wang et al. 2018). Some studies have reported that N fertilizer application can lead to substantial N2O emission through nitrification–denitrification processes (Loecke and Robertson 2009).
N addition reduced CH4 uptake and increased N2O emission. The alpine meadows of the Qinghai-Tibet Plateau are a weak sink of CH4 and a weak N2O source (Cao et al. 2008; Lin et al. 2009; Wang et al. 2009). CH4 and N2O emissions contributed little to CO2 equivalent emissions, because CH4 uptake and N2O emissions are very small (Table 2), and are insufficient to fundamentally affect the ecological system CO2 equivalent emissions (Jiang et al. 2010). CO2 emissions are therefore the main source of global warming potential in the treatments applied in this study.
Conclusions
Our results indicated that N addition had no effect on CO2 emission, but decreased CH4 uptake and increased N2O flux. Grazing exclusion decreased CO2 emission, but had no effect on CH4 uptake or N2O emission. There were no interactive effects of grazing exclusion and N fertilization on CO2, N2O or CH4. These results suggest that with an increase in N deposition and expansion of areas under grazing exclusion in the future, CO2 emission and CH4 uptake would decrease, N2O emission would increase, and total CO2 equivalent emissions would decrease. Therefore, increased N deposition and grazing exclusion could cause negative feedback to global warming in the alpine meadow. Our results suggest that exclusure from livestock grazing might be a promising measure to reduce ecosystem CO2 emissions in the alpine meadow region.
References
Adamsen A, King G (1993) Methane consumption in temperate and subarctic forest soils-rates, vertical zonation, and responses to water and nitrogen. Appl Environ Microbiol 59:485–490. https://doi.org/10.1128/AEM.59.2.485-490.1993
Alster CJ, German DP, Lu Y, Allison SD (2013) Microbial enzymatic responses to drought and to nitrogen addition in a southern California grassland. Soil Biol Biochem 64:68–79. https://doi.org/10.1016/j.soilbio.2013.03.034
Ambus P, Robertson GP (2006) The effect of increased N deposition on nitrous oxide, methane and carbon dioxide fluxes from unmanaged forest and grassland communities in Michigan. Biogeochemistry 79:315–337. https://doi.org/10.1007/s10533-005-5313-x
Bala G, Krishna S, Narayanappa D, Cao L, Caldeira K, Nemani R (2013) An estimate of equilibrium sensitivity of global terrestrial carbon cycle using NCAR CCSM4. Clim Dyn 40:1671–1686. https://doi.org/10.1007/s00382-012-1495-9
Bowman J, Sly L, Stackebrandt E (1995) The phylogenetic position of the family methylococcaceae. Int J Syst Bacteriol 45:182–185. https://doi.org/10.1099/00207713-45-1-182
Cai ZC, Shan YH, Xu H (2007) Effects of nitrogen fertilization on CH4 emissions from rice fields. Soil Sci Plant Nutr 53:353–361. https://doi.org/10.1111/j.1747-0765.2007.00153.x
Cantarel AAM, Bloor JMG, Deltroy N, Soussana JF (2011) Effects of climate change drivers on nitrous oxide fluxes in an upland temperate grassland. Ecosystems 14:223–233. https://doi.org/10.1007/s10021-010-9405-7)
Cao G, Xu X, Long R, Wang Q, Wang C, Du Y, Zhao X (2008) Methane emissions by alpine plant communities in the Qinghai-Tibet Plateau. Biol Lett 4:681–684. https://doi.org/10.1098/rsbl.2008.0373
Carter MS, Ambus P, Albert KR, Larsen KS, Andersson M, Priemé A, van der Linden L, Beier C (2011) Effects of elevated atmospheric CO2, prolonged summer drought and temperature increase on N2O and CH4 fluxes in a temperate heathland. Soil Biol Biochem 43:1660–1670. https://doi.org/10.1016/j.soilbio.2011.04.003
Carter MS, Larsen KS, Emmett B, Estiarte M, Field C, Leith ID, Lund M, Meijide A, Mills RTE, Niinemets Ü, Peñuelas J, Portillo-Estrada M, Schmidt IK, Selsted MB, Sheppard LJ, Sowerby A, Tietema A, Beier C (2012) Synthesizing greenhouse gas fluxes across nine European peatlands and shrublands—responses to climatic and environmental changes. Biogeosciences 9:3739–3755. https://doi.org/10.5194/bg-9-3739-2012
Chen J, Zhou X, Wang J, Hruska T, Shi W, Cao J, Zhang B, Xu G, Chen Y, Luo Y (2016) Grazing exclusion reduced soil respiration but increased its temperature sensitivity in a Meadow Grassland on the Tibetan Plateau. Ecol Evol 6:675–687. https://doi.org/10.1002/ece3.1867
Chen M, Chen Y, Dong S, Lan S, Zhou H, Tan Z, Li X (2018) Mixed nitrifying bacteria culture under different temperature dropping strategies: nitrification performance, activity, and community. Chemosphere 195:800–809. https://doi.org/10.1016/j.chemosphere.2017.12.129
Chinese Soil Taxonomy Research Group (1995) Chinese soil taxonomy. Science Press, Beijing, pp 58–147
Curry CL (2007) Modeling the soil consumption of atmospheric methane at the global scale. Glob Biogeochem Cycles. https://doi.org/10.1029/2006gb002818
Curtis CJ, Emmett BA, Reynolds B, Shilland J (2006) How important is N2O production in removing atmospherically deposited nitrogen from UK moorland catchments? Soil Biol Biochem 38:2081–2091. https://doi.org/10.1016/j.soilbio.2006.01.013
Delatolla R, Tufenkji N, Comeau Y, Gadbois A, Lamarre D, Berk D (2012) Effects of long exposure to low temperatures on nitrifying biofilm and biomass in wastewater treatment. Water Environ Res 84:328–338. https://doi.org/10.2175/106143012x13354606450924
Dijkstra FA, Morgan JA, Follett RF, Lecain DR (2013) Climate change reduces the net sink of CH4 and N2O in a semiarid grassland. Global Change Biol 19:1816–1826. https://doi.org/10.1111/gcb.12182
Fang C, Moncrieff J (2001) The dependence of soil CO2 efflux on temperature. Soil Biol Biochem 33:155–165. https://doi.org/10.1016/S0038-0717(00)00125-5
Galloway J, Cowling E (2002) Reactive nitrogen and the world: 200 years of change. Ambio 31:64–71. https://doi.org/10.1639/0044-7447(2002)031%5b0064:RNATWY%5d2.0.CO;2
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892. https://doi.org/10.1126/science.1136674
Goldberg SD, Gebauer G (2009) Drought turns a Central European Norway spruce forest soil from an N2O source to a transient N2O sink. Global Change Biol 15:850–860. https://doi.org/10.1111/j.1365-2486.2008.01752.x
Hart SC (2006) Potential impacts of climate change on nitrogen transformations and greenhouse gas fluxes in forests: a soil transfer study. Global Change Biol 12:1032–1046. https://doi.org/10.1111/j.1365-2486.2006.01159.x
Hoang V, Delatolla R, Abujamel T, Mottawea W, Gadbois A, Laflamme E, Stintzi A (2014) Nitrifying moving bed biofilm reactor (MBBR) biofilm and biomass response to long term exposure to 1 °C. Water Res 49:215–224. https://doi.org/10.1016/j.watres.2013.11.018
Hu YG, Chang XF, Lin XW, Wang YF, Wang SP, Duan JC, Zhang ZH, Yang XX, Luo CY, Xu GP, Zhao XQ (2010) Effects of warming and grazing on N2O fluxes in an alpine meadow ecosystem on the Tibetan plateau. Soil Biol Biochem 42:944–952. https://doi.org/10.1016/j.soilbio.2010.02.011
Hu YG, Zhang ZH, Wang SP, Zhang ZS, Zhao Y, Wang ZR (2017) The weak effects of fencing on ecosystem respiration, CH4, and N2O fluxes in a Tibetan alpine meadow during the growing season. Sci Cold Arid Regions 9:554–567
Hulsen T, Barry EM, Lu Y, Puyol D, Batstone DJ (2016) Low temperature treatment of domestic wastewater by purple phototrophic bacteria: performance, activity, and community. Water Res 100:537–545. https://doi.org/10.1016/j.watres.2016.05.054
IPCC (Intergovernmental Panel on Climate Change) (2007) Climate change 2007: summary for policymakers. Valencia, Spain
Jang I, Lee S, Hong J-H, Kang H (2006) Methane oxidation rates in forest soils and their controlling variables: a review and a case study in Korea. Ecol Res 21:849–854. https://doi.org/10.1007/s11284-006-0041-9
Jarvis S, Lovell R, Panayides R (1995) Patterns of methane emission from excreta of grazing animals. Soil Biol Biochem 27:1581–1588. https://doi.org/10.1016/0038-0717(95)00092-S
Jiang CM, Yu GR, Fang HJ, Cao GM, Li YN (2010) Short-term effect of increasing nitrogen deposition on CO2, CH4 and N2O fluxes in an alpine meadow on the Qinghai-Tibetan Plateau, China. Atmos Environ 44:2920–2926. https://doi.org/10.1016/j.atmosenv.2010.03.030
Juutinen S, Bubier JL, Moore TR (2010) Responses of vegetation and ecosystem CO2 exchange to 9 years of nutrient addition at Mer Bleue Bog. Ecosystems 13:874–887. https://doi.org/10.1007/s10021-010-9361-2
Kostyanovsky KI, Huggins DR, Stockle CO, Waldo S, Lamb B (2018) Developing a flow through chamber system for automated measurements of soil N2O and CO2 emissions. Measurement 113:172–180. https://doi.org/10.1016/j.measurement.2017.05.040
Larsen KS, Andresen LC, Beier C, Jonasson S, Albert KR, Ambus PER, Arndal MF, Carter MS, Christensen S, Holmstrup M, Ibrom A, Kongstad J, Van Der Linden L, Maraldo K, Michelsen A, Mikkelsen TN, Pilegaard KIM, PriemÉ A, Ro-Poulsen H, Schmidt IK, Selsted MB, Stevnbak K (2011) Reduced N cycling in response to elevated CO2, warming, and drought in a Danish heathland: synthesizing results of the CLIMAITE project after two years of treatments. Global Change Biol 17:1884–1899. https://doi.org/10.1111/j.1365-2486.2010.02351.x
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379. https://doi.org/10.1890/06-2057.1
Lin XW, Wang SP, Ma XZ, Xu GP, Luo CY, Li YN, Jiang GM, Xie ZB (2009) Fluxes of CO2, CH4, and N2O in an alpine meadow affected by yak excreta on the Qinghai-Tibetan plateau during summer grazing periods. Soil Biol Biochem 41:718–725. https://doi.org/10.1016/j.soilbio.2009.01.007
Lin XW, Wang SP, Hu YG, Luo CY, Zhang ZH, Niu HS, Xie ZB (2015) Experimental warming increases seasonal methane uptake in an alpine meadow on the Tibetan Plateau. Ecosystems 18:274–286. https://doi.org/10.1007/s10021-014-9828-7
Liu LL, Greaver TL (2009) A review of nitrogen enrichment effects on three biogenic GHGs: the CO2 sink may be largely offset by stimulated N2O and CH4 emission. Ecol Lett 12:1103–1117. https://doi.org/10.1111/j.1461-0248.2009.01351.x
Liu CY, Holst J, Brüggemann N, Butterbach-Bahl K, Yao ZS, Yue J, Han SH, Han XG, Krümmelbein J, Horn R, Zheng XH (2007) Winter-grazing reduces methane uptake by soils of a typical semi-arid steppe in Inner Mongolia. China Atmos Environ 41:5948–5958. https://doi.org/10.1016/j.atmosenv.2007.03.017
Liu X, Zhang Y, Han W, Tang A, Shen J, Cui Z, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang F (2013a) Enhanced nitrogen deposition over China. Nature 494:459–462. https://doi.org/10.1038/nature11917
Liu YW, Xu R, Xu XL, Wei D, Wang YH, Wang YS (2013b) Plant and soil responses of an alpine steppe on the Tibetan Plateau to multi-level nitrogen addition. Plant Soil 373:515–529. https://doi.org/10.1007/s11104-013-1814-x
Lloyd J, Taylor J (1994) On the temperature dependence of soil respiration. Funct Ecol 8:315–323. https://doi.org/10.2307/2389824
Loecke TD, Robertson GP (2009) Soil resource heterogeneity in terms of litter aggregation promotes nitrous oxide fluxes and slows decomposition. Soil Biol Biochem 41:228–235. https://doi.org/10.1016/j.soilbio.2008.10.017
Lü CQ, Tian HQ (2007) Spatial and temporal patterns of nitrogen deposition in China: synthesis of observational data. J Geophys Res. https://doi.org/10.1029/2006jd007990
Luo C, Xu G, Wang Y, Wang S, Lin X, Hu Y, Zhang Z, Chang X, Duan J, Su A (2009) Effects of grazing and experimental warming on DOC concentrations in the soil solution on the Qinghai-Tibet plateau. Soil Biol Biochem 41:2493–2500. https://doi.org/10.1016/j.soilbio.2009.09.006
Luo C, Xu G, Chao Z, Wang S, Lin X, Hu Y, Zhang Z, Duan J, Chang X, Su A, Li Y, Zhao X, Du M, Tang Y, Kimball B (2010) Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan plateau. Global Change Biol 16:1606–1617. https://doi.org/10.1111/j.1365-2486.2009.02026.x
Luo CY, Zhu XX, Wang SP, Cui SJ, Zhang ZH, Bao XY, Zhao L, Li Y, Zhao XQ (2015) Ecosystem carbon exchange under different land use on the Qinghai-Tibetan plateau. Photosynthetica 53:527–536. https://doi.org/10.1007/s11099-015-0142-1
McSherry ME, Ritchie ME (2013) Effects of grazing on grassland soil carbon: a global review. Global Change Biol 19:1347–1357. https://doi.org/10.1111/gcb.12144
Peng Q, Qi YC, Dong YS, Xiao SS, He YT (2011) Soil nitrous oxide emissions from a typical semiarid temperate steppe in inner Mongolia: effects of mineral nitrogen fertilizer levels and forms. Plant Soil 342:345–357. https://doi.org/10.1007/s11104-010-0699-1
Peng Y, Li F, Zhou G, Fang K, Zhang D, Li C, Yang G, Wang G, Wang J, Yang Y (2017) Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe. Global Change Biol 23:5249–5259. https://doi.org/10.1111/gcb.13789
Pregitzer KS, Burton AJ, Zak DR, Talhelm AF (2008) Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests. Global Change Biol 14:142–153. https://doi.org/10.1111/j.1365-2486.2007.01465.x
Reay DS, Dentener F, Pete S, Grace J, Feely RA (2008) Global nitrogen deposition and carbon sinks. Nat Geosci 1:430–437. https://doi.org/10.1038/ngeo230
Rodriguez A, Lovett GM, Weathers KC, Arthur MA, Templer PH, Goodale CL, Christenson LM (2014) Lability of C in temperate forest soils: assessing the role of nitrogen addition and tree species composition. Soil Biol Biochem 77:129–140. https://doi.org/10.1016/j.soilbio.2014.06.025
Saggar S, Hedley CB, Giltrap DL, Lambie SM (2007) Measured and modelled estimates of nitrous oxide emission and methane consumption from a sheep-grazed pasture. Agric Ecosyst Environ 122:357–365. https://doi.org/10.1016/j.agee.2007.02.006
Scheutz C, Kjeldsen P (2004) Environmental factors influencing attenuation of methane and hydrochlorofluorocarbons in landfill cover soils. J Environ Qual 33:72–79. https://doi.org/10.2134/jeq2004.0072
Sherlock RR, Sommer SG, Khan RZ, Wood CW, Guertal EA, Freney JR, Dawson CO, Cameron KC (2002) Ammonia, methane, and nitrous oxide emission from pig slurry applied to a pasture in New Zealand. J Environ Qual 31:1491–1501. https://doi.org/10.2134/jeq2002.1491
Shi FS, Chen H, Chen HF, Wu Y, Wu N (2012) The combined effects of warming and drying suppress CO2 and N2O emission rates in an alpine meadow of the eastern Tibetan Plateau. Ecol Res 27:725–733. https://doi.org/10.1007/s11284-012-0950-8
Siripong S, Rittmann BE (2007) Diversity study of nitrifying bacteria in full-scale municipal wastewater treatment plants. Water Res 41:1110–1120. https://doi.org/10.1016/j.watres.2006.11.050
Teh YA, Diem T, Jones S, Huaraca Quispe LP, Baggs E, Morley N, Richards M, Smith P, Meir P (2014) Methane and nitrous oxide fluxes across an elevation gradient in the tropical Peruvian Andes. Biogeosciences 11:2325–2339. https://doi.org/10.5194/bg-11-2325-2014
Urakawa H, Tajima Y, Numata Y, Tsuneda S (2008) Low temperature decreases the phylogenetic diversity of ammonia-oxidizing archaea and bacteria in aquarium biofiltration systems. Appl Environ Microbiol 74:894–900. https://doi.org/10.1128/AEM.01529-07
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, Schlesinger WH, David G, Tilman D (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750. https://doi.org/10.2307/2269431
Wang ZP, Song Y, Gulledge J, Yu Q, Liu HS, Han XG (2009) China’s grazed temperate grasslands are a net source of atmospheric methane. Atmos Environ 43:2148–2153. https://doi.org/10.1016/j.atmosenv.2009.01.021
Wang SP, Duan JC, Xu GP, Wang YF, Zhang ZH, Rui YC, Luo CY, Xu BRBY, Zhu XX, Chang XF, Cui XY, Niu HS, Zhao XQ, Wang WY (2012) Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 93:2365–2376. https://doi.org/10.1890/11-1408.1
Wang H, Yu L, Zhang Z, Liu W, Chen L, Cao G, Yue H, Zhou J, Yang Y, Tang Y, He JS (2017) Molecular mechanisms of water table lowering and nitrogen deposition in affecting greenhouse gas emissions from a Tibetan alpine wetland. Global Change Biol 23:815–829. https://doi.org/10.1111/gcb.13467
Wang GS, Liang YP, Ren F, Yang XX, Mi ZR, Gao Y, George TS, Zhang ZH (2018) Greenhouse gas emissions from the Tibetan alpine grassland: effects of nitrogen and phosphorus addition. Sustainability 10:4454. https://doi.org/10.3390/su10124454
Wei D, Xu R, Wang YH, Wang YS, Liu YW, Yao TD (2012) Responses of CO2, CH4 and N2O fluxes to livestock exclosure in an alpine steppe on the Tibetan Plateau, China. Plant Soil 359:45–55. https://doi.org/10.1007/s11104-011-1105-3
Wolf B, Zheng X, Bruggemann N, Chen W, Dannenmann M, Han X, Sutton MA, Wu H, Yao Z, Butterbach-Bahl K (2010) Grazing-induced reduction of natural nitrous oxide release from continental steppe. Nature 464:881–884. https://doi.org/10.1038/nature08931
WRB (2006) World reference base for soil resources. FAO/ISRIC/ISSS, Rome
Xiao Y (2017) Greenhouse gas emissions from paddy soils respond to different crop root residues and N fertilizer types. Water Air Soil Pollut. https://doi.org/10.1007/s11270-017-3594-z
Xiong DP, Shi PL, Zhang XZ, Zou CB (2016) Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China—A meta-analysis. Ecol Eng 94:647–655. https://doi.org/10.1016/j.ecoleng.2016.06.124
Zhang ZH, Duan JC, Wang SP, Luo CY, Chang XF, Zhu XX, Xu BRBY, Wang WY (2012) Effects of land use and management on ecosystem respiration in alpine meadow on the Tibetan plateau. Soil Tillage Res 124:161–169. https://doi.org/10.1016/j.still.2012.05.012
Zhang WN, Ganjurjav H, Liang Y, Gao QZ, Wan YF, Li Y, Baima YZ, Xirao ZM (2015) Effect of a grazing ban on restoring the degraded alpine meadows of Northern Tibet, China. Rangel J 37:89. https://doi.org/10.1071/rj14092
Zheng Y, Yang W, Hu HW, Kim YC, Duan JC, Luo CY, Wang SP, Guo LD (2014) Ammonia oxidizers and denitrifiers in response to reciprocal elevation translocation in an alpine meadow on the Tibetan Plateau. J Soils Sed 14:1189–1199. https://doi.org/10.1007/s11368-014-0867-7
Zhou L, Zhou X, Zhang B, Lu M, Luo Y, Liu L, Li B (2014) Different responses of soil respiration and its components to nitrogen addition among biomes: a meta-analysis. Global Change Biol 20:2332–2343. https://doi.org/10.1111/gcb.12490
Zhou H, Li X, Chu Z, Zhang J (2016) Effect of temperature downshifts on a bench-scale hybrid A/O system: process performance and microbial community dynamics. Chemosphere 153:500–507. https://doi.org/10.1016/j.chemosphere.2016.03.092
Zhu XX, Luo CY, Wang SP, Zhang ZH, Cui SJ, Bao XY, Jiang LL, Li YM, Li XE, Wang Q, Zhou Y (2015) Effects of warming, grazing/cutting and nitrogen fertilization on greenhouse gas fluxes during growing seasons in an alpine meadow on the Tibetan Plateau. Agric For Meteorol 214–215:506–514. https://doi.org/10.1016/j.agrformet.2015.09.008
Zhu C, Ma Y, Wu H, Sun T, La Pierre KJ, Sun Z, Yu Q (2016a) Divergent effects of nitrogen addition on soil respiration in a semiarid grassland. Sci Rep 6:33541. https://doi.org/10.1038/srep33541
Zhu J, Zhang Y, Liu Y (2016b) Effects of short-term grazing exclusion on plant phenology and reproductive succession in a Tibetan alpine meadow. Sci Rep 6:27781. https://doi.org/10.1038/srep27781
Zhuang Q, Melillo J, McGuire A, Kicklighter D, Prinn R, Steudler P, Felzer B, Hu S (2007) Net emissions of CH4 and CO2 in Alaska: implications for the region’s greenhouse gas budget. Ecol. Appl. 17:203–212. https://doi.org/10.1890/1051-0761(2007)017%5b0203:NEOCAC%5d2.0.CO;2
Zou J, Zhao L, Xu S, Xu X, Chen D, Li Q, Zhao N, Luo C, Zhao X (2014) Field 13CO2 pulse labeling reveals differential partitioning patterns of photoassimilated carbon in response to livestock exclosure in a Kobresia meadow. Biogeosciences 11:4381–4391. https://doi.org/10.5194/bg-11-4381-2014
Zou JR, Luo CY, Xu XL, Zhao N, Zhao L, Zhao XQ (2016) Relationship of plant diversity with litter and soil available nitrogen in an alpine meadow under a 9-year grazing exclusion. Ecol Res 31:841–851. https://doi.org/10.1007/s11284-016-1394-3
Acknowledgements
This work was supported by projects from the National Key Research and Development Program of China (2016YFC0501805 and 2016YFC0501905), the Science and Technology Bureau of Qinghai (2017-ZJ-716 and 2018-SF-110-1), the National Science Foundation of China (41731175), and the Qilian Mountain National Park Research Center (Qinghai) (Grant Number: GKQ 2019-01).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10705_2020_10062_MOESM1_ESM.docx
Fig. S1. Daily average temperature and daily precipitation from 1 June to 30 September in 2011, 2012 and 2013 (DOCX 301 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Luo, C., Wang, S., Zhang, L. et al. CO2, CH4 and N2O fluxes in an alpine meadow on the Tibetan Plateau as affected by N-addition and grazing exclusion. Nutr Cycl Agroecosyst 117, 29–42 (2020). https://doi.org/10.1007/s10705-020-10062-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-020-10062-0