The C:N:P Stoichiometry of Planted and Natural Larix principis-rupprechtii Stands along Altitudinal Gradients on the Loess Plateau, China

 , ,
, ,
Abstract
:1. Introduction
- Reveal the elevational and stand origin effects on the C:N:P stoichiometry of leaves, litter, and soil at the community level.
- Determine the plant nutrient resorption and limiting status.
- Explore the nutrient conservation strategies of the larch forest in our study region.
- Planted forests have higher resorption efficiencies and nutrient limitations than natural forests. From the perspective of leaf economics, if the uptake of nutrients from the soil is too energy-demanding for plants, they might employ an internal nutrient cycling strategy that involves enhanced nutrient resorption. Due to the variable disturbance histories between planted and natural forests, they may have different nutrient conservation strategies.
- We further hypothesized that: planted forests converged toward a highly conservative nutrient utilization strategy by enhancing nutrient resorption efficiencies; however, natural forests tended to a strategy that improved soil nutrient-status and utilization efficiencies.
2. Materials and Methods
2.1. Study Site
2.2. Sampling Design
2.3. Sample Treatments and Determination
2.4. Statistical Analysis
3. Results
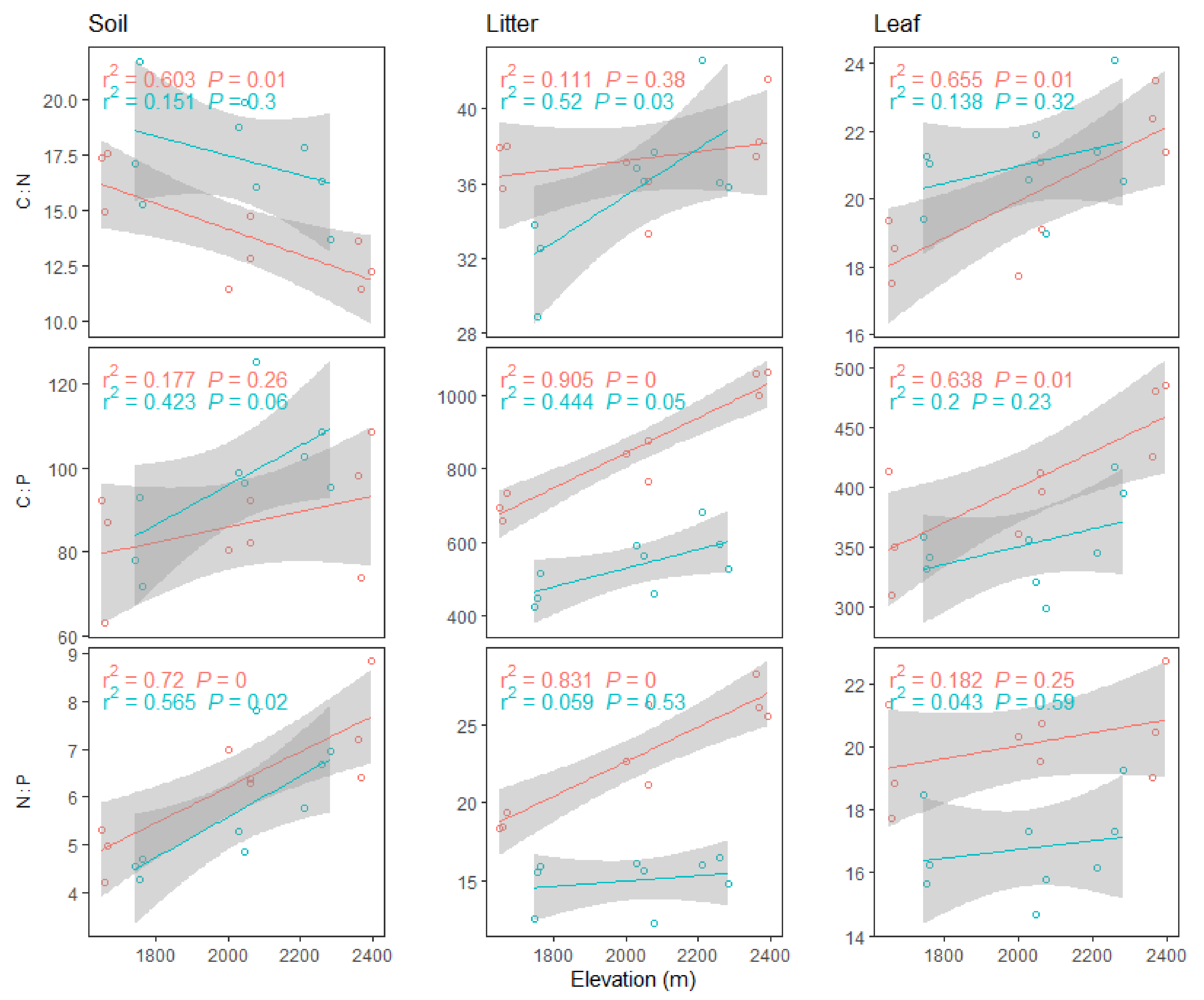
3.1. Altitudinal Patterns of Soil, Litter, and Leaf Carbon (C), Nitrogen (N), and Phosphorus (P) Content
3.2. Altitudinal Patterns of Soil, Litter, and Leaf C:N:P Ratios
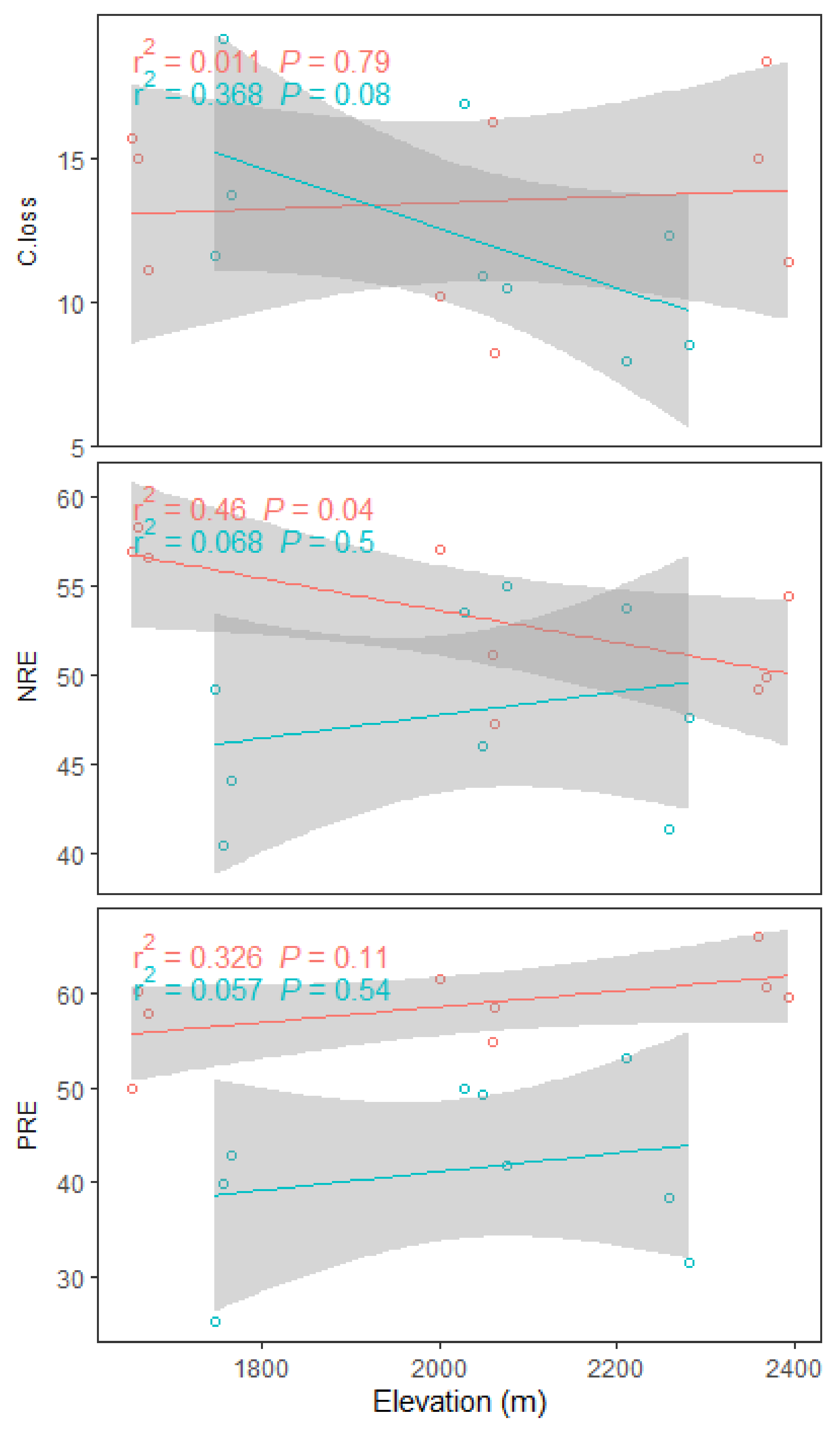
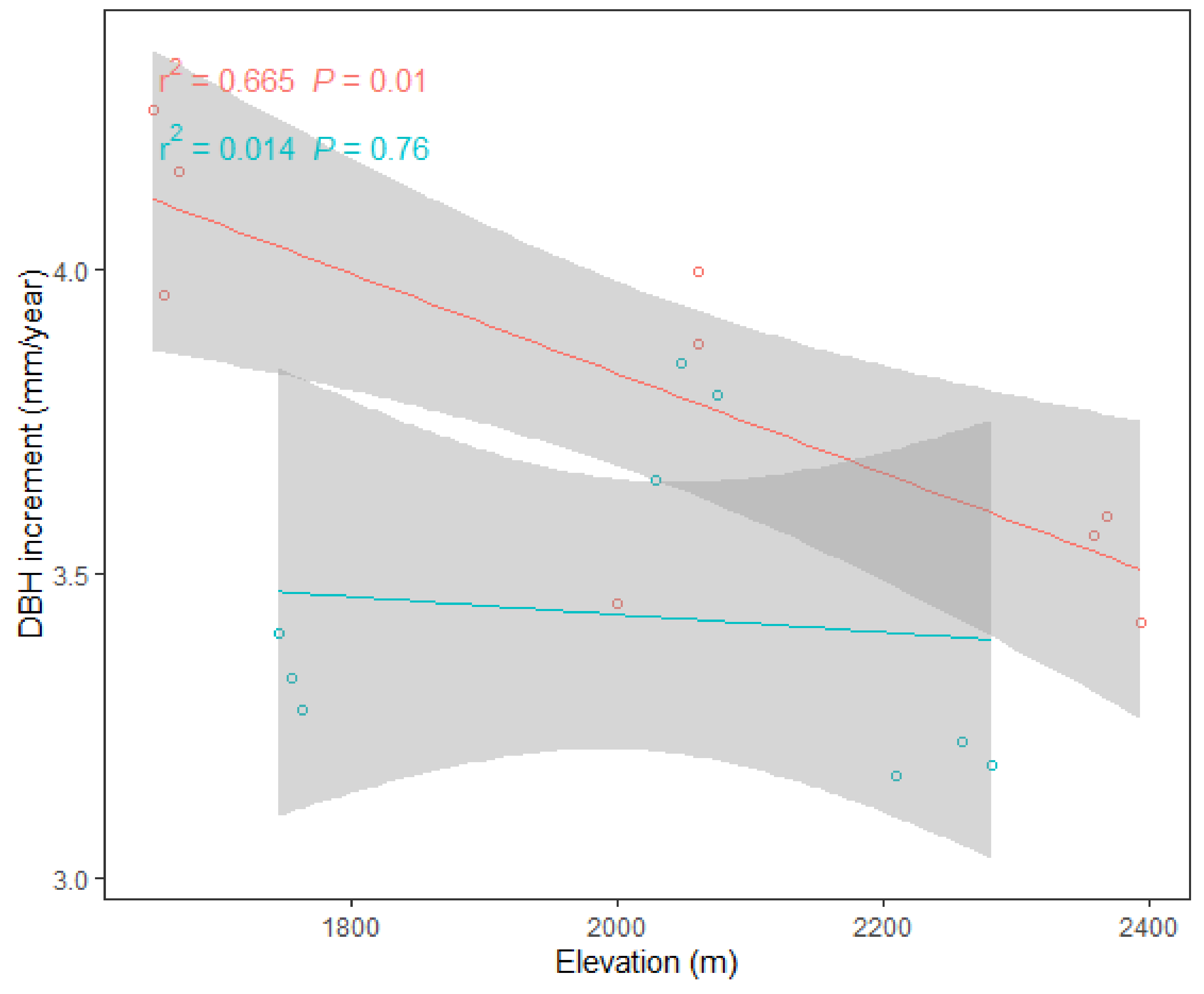
3.3. Effects of Elevation and Forest Management on Leaf Nutrient Resorption Efficiency (NuRE) and Plant Growth
4. Discussion
4.1. Variations of Soil, Litter, and Leaf C, N and P Content, and Ratios and Correlations
4.2. Response of NuRE and Growth to Elevation and Forest Management Indicates a Nutrient Conservation Strategy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elser, J.J.; Dobberfuhl, D.R.; Mackay, N.A.; Schampel, J.H. Organism size, life history, and n:P stoichiometrytoward a unified view of cellular and ecosystem processes. Bioscience 1996, 46, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusewell, S. High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytol. 2005, 166, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, H.; Yu, Q.; Wang, Z.; Wei, C.; Long, M.; Kattge, J.; Smith, M.; Han, X. Sampling date, leaf age and root size: Implications for the study of plant c:N:P stoichiometry. PLoS ONE 2013, 8, e60360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.X.; Fang, L.C.; Deng, L.; Guo, X.B.; Han, F.; Ju, W.L.; Wang, X.; Chen, H.S.; Tan, W.F.; Zhang, X.C. Patterns of soil microbial nutrient limitations and their roles in the variation of soil organic carbon across a precipitation gradient in an arid and semi-arid region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Janssens, I.A.; Alonso, R.; Veresoglou, S.D.; Rillig, M.C.; Sanders, T.G.M.; Carnicer, J.; Filella, I.; Farre-Armengol, G.; Penuelas, J. Foliar elemental composition of european forest tree species associated with evolutionary traits and present environmental and competitive conditions. Glob. Ecol. Biogeogr. 2015, 24, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.H. Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat. Clim. Change 2015, 5, 465–469. [Google Scholar] [CrossRef]
- Tian, D.; Reich, P.B.; Chen, H.Y.H.; Xiang, Y.; Luo, Y.; Shen, Y.; Meng, C.; Han, W.; Niu, S. Global changes alter plant multi-element stoichiometric coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; Van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-microbe-soil interactions in the rhizosphere: An evolutionary perspective. Plant Soil 2009, 321, 83–115. [Google Scholar] [CrossRef]
- Aerts, R. Nutrient resorption from senescing leaves of perennials: Are there general patterns? J. Ecol. 1996, 84, 597–608. [Google Scholar] [CrossRef]
- Brant, A.N.; Chen, H.Y.H. Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 1999, 30, 1–67. [Google Scholar]
- Hayes, P.; Turner, B.L.; Lambers, H.; Laliberte, E. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 2014, 102, 396–410. [Google Scholar] [CrossRef]
- Mayor, J.R.; Sanders, N.J.; Classen, A.T.; Bardgett, R.D.; Clement, J.C.; Fajardo, A.; Lavorel, S.; Sundqvist, M.K.; Bahn, M.; Chisholm, C.; et al. Elevation alters ecosystem properties across temperate treelines globally. Nature 2017, 542, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Piaget, A. Mountain weather and climate. EOS Trans. AGU 1982, 63, 1225. [Google Scholar] [CrossRef]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annu. Rev. Ecol. Evol. S. 2013, 44, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, M.K.; Giesler, R.; Wardle, D.A. Within- and across-species responses of plant traits and litter decomposition to elevation across contrasting vegetation types in subarctic tundra. PLoS ONE 2011, 6, e27056. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Hou, E.; Liu, Y.; Wen, D. Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical china. Sci. Rep. 2016, 6, 24261. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Z.; Zheng, H.; Xiao, Y.; Polasky, S.; Liu, J.; Xu, W.; Wang, Q.; Zhang, L.; Xiao, Y.; Rao, E.; et al. Improvements in ecosystem services from investments in natural capital. Science 2016, 352, 1455–1459. [Google Scholar] [CrossRef]
- Graham, H. Ecological stoichiometry: Biology of elements from molecules to the biosphere. J. Plankton Res. 2002, 25, 1183. [Google Scholar]
- Boczulak, S.A.; Hawkins, B.J.; Maynard, D.G.; Roy, R. Long- and short-term temperature differences affect organic and inorganic nitrogen availability in forest soils. Can. J. Soil Sci. 2015, 95, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Macek, P.; Klimeš, L.; Adamec, L.; Doležal, J.; Chlumská, Z.; De Bello, F.; Dvorský, M.; Řeháková, K. Plant nutrient content does not simply increase with elevation under the extreme environmental conditions of ladakh, nw himalaya. Arct. Antarc. Alpine Res. 2018, 44, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Krner, C. The nutritional status of plants from high altitudes-a worldwide comparison. Oecologia 1989, 81, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Van de Weg, M.J.; Meir, P.; Grace, J.; Atkin, O.K. Altitudinal variation in leaf mass per unit area, leaf tissue density and foliar nitrogen and phosphorus content along an amazon-andes gradient in peru. Plant Ecol. Div. 2009, 2, 243–254. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. Factors affecting nutrient concentration and stoichiometry of forest trees in catalonia (ne spain). For. Ecol. Manag. 2011, 262, 2024–2034. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhao, N.; Liu, C.C.; Yang, H.; Li, M.L.; Yu, G.R.; Wilcox, K.; Yu, Q.; He, N.P. C:N:P stoichiometry in china’s forests: From organs to ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef]
- Murphy, K.L.; Klopatek, J.M.; Klopatek, C.C. The effects of litter quality and climate on decomposition along an elevational gradient. Ecol. Appl. 1998, 8, 1061–1071. [Google Scholar] [CrossRef]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Vitousek, P.M.; Cuevas, E. Experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology 1998, 79, 10. [Google Scholar] [CrossRef]
- Xu, G.P.; Hu, Y.G.; Wang, S.P.; Zhang, Z.H.; Chang, X.F.; Duan, J.C.; Luo, C.Y.; Chao, Z.G.; Su, A.L.; Lin, Q.Y.; et al. Effects of litter quality and climate change along an elevation gradient on litter mass loss in an alpine meadow ecosystem on the tibetan plateau. Plant Ecol. 2010, 209, 257–268. [Google Scholar] [CrossRef]
- Agren, G.I.; Weih, M. Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype. New Phytol. 2012, 194, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, Z.; Xing, W.; Liu, G. Plasticity in latitudinal patterns of leaf n and p of oryza rufipogon in china. Plant Biol. 2014, 16, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Shumilov, O.; Kanatjev, A.; Kasatkina, E. A measuring tool for tree-rings analysis. Geophys. Res. Abstr. 2013, 15, 2347. [Google Scholar]
- Austin, A.T.; Vivanco, L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 2006, 442, 555–558. [Google Scholar] [CrossRef]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S.; Adler, D.; Bates, D.; Baud-Bovy, G.; Ellison, S.; Firth, D.; Friendly, M.; Gorjanc, G.; Graves, S. Package ‘car’; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 3.6.0; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.Rproject.org/ (accessed on 24 March 2020).
- Wardle, D.A.; Hornberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf n and p in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Walbridge, M.R.; Richardson, C.J.; Swank, W.T. Vertical distribution of biological and geochemical phosphorus subcycles in two southern appalachian forest soils. Biogeochemistry 1991, 13, 61–85. [Google Scholar] [CrossRef]
- Tiessen, H.; Stewart, J.W.B.; Cole, C.V. Pathways of phosphorus transformations in soils of differing pedogenesis1. Soil Sci. Soc. Am. J. 1984, 48, 853. [Google Scholar] [CrossRef]
- Leitner, S.; Wanek, W.; Wild, B.; Haemmerle, I.; Kohl, L.; Keiblinger, K.M.; Zechmeister-Boltenstern, S.; Richter, A. Influence of litter chemistry and stoichiometry on glucan depolymerization during decomposition of beech (fagus sylvatica l.) litter. Soil Biol. Biochem. 2012, 50, 174–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Mcgroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of c:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, Y. Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Hume, A.; Chen, H.Y.H.; Taylor, A.R.; Kayahara, G.J.; Man, R.Z. Soil c:N:P dynamics during secondary succession following fire in the boreal forest of central canada. For. Ecol. Manag. 2016, 369, 1–9. [Google Scholar] [CrossRef]
- Hume, A.M.; Chen, H.Y.H.; Taylor, A.R. Intensive forest harvesting increases susceptibility of northern forest soils to carbon, nitrogen and phosphorus loss. J. Appl. Ecol. 2018, 55, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Smethurst, P.J.; Sadanandan Nambiar, E.K. Changes in soil carbon and nitrogen during the establishment of a second crop of pinus radiata. For. Ecol. Manag. 1995, 73, 145–155. [Google Scholar] [CrossRef]
- Stark, S.; Ylanne, H.; Tolvanen, A.T. Long-term warming alters soil and enzymatic n:P stoichiometry in subarctic tundra. Soil Biol. Biochem. 2018, 124, 184–188. [Google Scholar] [CrossRef]
- Xu, Z.W.; Yu, G.R.; Zhang, X.Y.; He, N.P.; Wang, Q.F.; Wang, S.Z.; Wang, R.L.; Zhao, N.; Jia, Y.L.; Wang, C.Y. Soil enzyme activity and stoichiometry in forest ecosystems along the north-south transect in eastern china (nstec). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Zhao, N.; He, N.; Wang, Q.; Zhang, X.; Wang, R.; Xu, Z.; Yu, G. The altitudinal patterns of leaf crationratiop stoichiometry are regulated by plant growth form, climate and soil on changbai mountain, china. PLoS ONE 2014, 9, e95196. [Google Scholar]
- Hoch, G.; Korner, C. Global patterns of mobile carbon stores in trees at the high-elevation tree line. Glob. Ecol. Biogeogr. 2012, 21, 861–871. [Google Scholar] [CrossRef]
- Hoch, G.; Popp, M.; Körner, C. Altitudinal increase of mobile carbon pools in pinus cembra suggests sink limitation of growth at the swiss treeline. Oikos 2010, 98, 361–374. [Google Scholar] [CrossRef]
- Santiago, L.S.; Kitajima, K.; Wright, S.J.; Mulkey, S.S. Coordinated changes in photosynthesis, water relations and leaf nutritional traits of canopy trees along a precipitation gradient in lowland tropical forest. Oecologia 2004, 139, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Sandel, B.; Goldstein, L.J.; Kraft, N.J.; Okie, J.G.; Shuldman, M.I.; Ackerly, D.D.; Cleland, E.E.; Suding, K.N. Contrasting trait responses in plant communities to experimental and geographic variation in precipitation. New Phytol. 2010, 188, 565–575. [Google Scholar] [CrossRef]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in china. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Müller, M.; Oelmann, Y.; Schickhoff, U.; Böhner, J.; Scholten, T. Himalayan treeline soil and foliar c:N:P stoichiometry indicate nutrient shortage with elevation. Geoderma 2017, 291, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Matzek, V.; Vitousek, P.M. N:P stoichiometry and protein:Rna ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecol. Lett. 2009, 12, 765–771. [Google Scholar] [CrossRef]
- Agren, G.I. The c:N:P stoichiometry of autotrophs–theory and observations. Ecol. Lett. 2010, 7, 185–191. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dj, T.J.R. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar]
- Reich, P.B. Global biogeography of plant chemistry: Filling in the blanks. New Phytol. 2010, 168, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Crews, T.E.; Kitayama, K.; Fownes, J.H.; Riley, R.H.; Herbert, D.A.; Muellerdombois, D.; Vitousek, P.M. Changes in soil-phosphorus fractions and ecosystem dynamics across a long chronosequence in hawaii. Ecology 1995, 76, 1407–1424. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; St Clair, S.B. Mineral stress: The missing link in understanding how global climate change will affect plants in real world soils. Field Crop Res. 2004, 90, 101–115. [Google Scholar] [CrossRef]
- Liao, Y.C.; McCormack, M.L.; Fan, H.B.; Wang, H.M.; Wu, J.P.; Tu, J.; Liu, W.F.; Guo, D.L. Relation of fine root distribution to soil c in a cunninghamia lanceolata plantation in subtropical china. Plant Soil 2014, 381, 225–234. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M.M. Controls over foliar n:P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef]
- Bowman, W.D.; Theodose, T.A.; Schardt, J.C.; Conant, R.T. Constraints of nutrient availability on primary production in two alpine tundra communities. Ecology 1993, 74, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Fife, D.N.; Nambiar, E.K.; Saur, E. Retranslocation of foliar nutrients in evergreen tree species planted in a mediterranean environment. Tree Physiol. 2008, 28, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleksyn, J.; Reich, P.B.; Zytkowiak, R.; Karolewski, P.; Tjoelker, M.G. Nutrient conservation increases with latitude of origin in european pinus sylvestris populations. Oecologia 2003, 136, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G.; Barnard, H.R.; Fownes, J. Age-related decline in forest ecosystem growth: An individual-tree, stand-structure hypothesis. Ecosystems 2002, 5, 58–67. [Google Scholar] [CrossRef]
- Searle, E.B.; Chen, H.Y.H. Complementarity effects are strengthened by competition intensity and global environmental change in the central boreal forests of canada. Ecol. Lett. 2020, 23, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Chen, H.Y.H. Changes in nitrogen resorption of trembling aspen (populus tremuloides) with stand development. Plant Soil 2010, 327, 121–129. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Cui, J.; Zhao, X.; Zhang, T. Nutrient resorption in Caragana microphylla along a chronosequence of plantations: Implications for desertified land restoration in North China. Ecol. Eng. 2013, 53, 299–305. [Google Scholar] [CrossRef]
- Ye, G.F.; Zhang, S.J.; Zhang, L.H.; Lin, Y.M.; Wei, S.D.; Liao, M.M.; Lin, G.H. Age-related changes in nutrient resorption patterns and tannin concentration of casuarina equisetifolia plantations. J. Trop. For. Sci. 2012, 24, 546–556. [Google Scholar]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Altitude Gradient | Altitude (m) | Slope Aspect | Slope Degree | Mean Tree Age (Years) | Mean Tree DBH (cm) | Mean Tree Height (m) | Stand Density (Trees per Hectare) | Canopy Cover |
|---|---|---|---|---|---|---|---|---|---|
| Plantation | P-H1 | 1694.16 | Shady slope | 17.00 | 41 | 24.55 | 18.28 | 925 | 0.57 |
| P-H2 | 2031.52 | semi-shady slope | 18.00 | 39 | 23.36 | 17.97 | 1158 | 0.61 | |
| P-H3 | 2330.19 | semi-shady slope | 24.00 | 41 | 22.48 | 15.71 | 1190 | 0.69 | |
| Natural Forest | N-H1 | 1732.07 | Shady slope | 19.00 | 66 | 33.10 | 23.61 | 875 | 0.52 |
| N-H2 | 2030.74 | Shady slope | 22.00 | 63 | 34.63 | 23.00 | 1050 | 0.53 | |
| N-H3 | 2293.56 | semi-shady slope | 21.00 | 65 | 31.64 | 25.12 | 1090 | 0.67 |
| Attribute | E | M | E × M | |||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| Soil C | 21.8 | <0.001 | <0.1 | 0.874 | 3.2 | 0.096 |
| Soil N | 49.2 | <0.001 | 14.8 | 0.002 | 0.1 | 0.734 |
| Soil P | 0.5 | 0.502 | 3.4 | 0.085 | <0.1 | 0.959 |
| Soil C:N | 8.1 | 0.013 | 12.1 | 0.004 | 0.1 | 0.735 |
| Soil C:P | 5.2 | 0.039 | 3.0 | 0.108 | 1.3 | 0.278 |
| Soil N:P | 26.1 | <0.001 | 2.5 | 0.138 | 0.1 | 0.746 |
| Litter C | 4.5 | 0.053 | 0.1 | 0.715 | 2.7 | 0.120 |
| Litter N | 1.6 | 0.231 | 2.4 | 0.141 | 1.6 | 0.232 |
| Litter P | 13.9 | 0.002 | 68.6 | <0.001 | <0.1 | 0.952 |
| Litter C:N | 5.9 | 0.029 | 1.9 | 0.187 | 4.0 | 0.065 |
| Litter C:P | 55.6 | <0.001 | 131.4 | <0.001 | 3.6 | 0.079 |
| Litter N:P | 28.8 | <0.001 | 103.5 | <0.001 | 8.3 | 0.012 |
| Leaf C | 6.7 | 0.021 | 0.6 | 0.453 | 0.1 | 0.740 |
| Leaf N | 9.1 | 0.009 | 4.0 | 0.066 | 1.5 | 0.248 |
| Leaf P | 9.4 | 0.008 | 8.0 | 0.014 | 0.6 | 0.434 |
| Leaf C:N | 11.8 | 0.004 | 2.2 | 0.164 | 1.1 | 0.309 |
| Leaf C:P | 14.1 | 0.002 | 9.1 | 0.009 | 1.1 | 0.312 |
| Leaf N:P | 1.9 | 0.189 | 22.3 | <0.001 | 0.1 | 0.825 |
| DBHi | 6.8 | 0.020 | 11.4 | 0.005 | 2.0 | 0.182 |
| C loss | 0.7 | 0.416 | 0.5 | 0.493 | 3.0 | 0.108 |
| NRE | 0.8 | 0.393 | 6.7 | 0.021 | 2.9 | 0.109 |
| PRE | 1.9 | 0.191 | 26.5 | <0.001 | <0.1 | 0.912 |
| Components | C | N | P | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| Soil vs. litter | 0.51 * | −0.24 | −0.46 | −0.35 | −0.06 | 0.44 |
| Soil vs. leaf | 0.49 * | −0.18 | −0.43 | 0.01 | 0.03 | 0.49 * |
| Litter vs. leaf | 0.3 | 0.05 | 0.65 ** | 0.08 | 0.73 *** | 0.69 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bo, F.; Zhang, Y.; Chen, H.Y.H.; Wang, P.; Ren, X.; Guo, J. The C:N:P Stoichiometry of Planted and Natural Larix principis-rupprechtii Stands along Altitudinal Gradients on the Loess Plateau, China. Forests 2020, 11, 363. https://doi.org/10.3390/f11040363
Bo F, Zhang Y, Chen HYH, Wang P, Ren X, Guo J. The C:N:P Stoichiometry of Planted and Natural Larix principis-rupprechtii Stands along Altitudinal Gradients on the Loess Plateau, China. Forests. 2020; 11(4):363. https://doi.org/10.3390/f11040363
Chicago/Turabian StyleBo, Fujing, Yunxiang Zhang, Han Y. H. Chen, Pingan Wang, Xuming Ren, and Jinping Guo. 2020. "The C:N:P Stoichiometry of Planted and Natural Larix principis-rupprechtii Stands along Altitudinal Gradients on the Loess Plateau, China" Forests 11, no. 4: 363. https://doi.org/10.3390/f11040363