Evaluation of the Field Efficacy of Heterorhabditis Bacteriophora Poinar (Rhabditida: Heterorhabditidae) and Synthetic Insecticides for the Control of Western Corn Rootworm Larvae




Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites and Experimental Set-Up
2.2. Treatments
2.3. Evaluation of Treatment Efficacy
2.4. Evaluation of Maize Root Damage, Plant Lodging, Fresh Plant Weight, and Fresh Grain Yield Per Plant
2.5. Assessment of Nematode Persistence in Soil
2.6. Data Analysis
3. Results
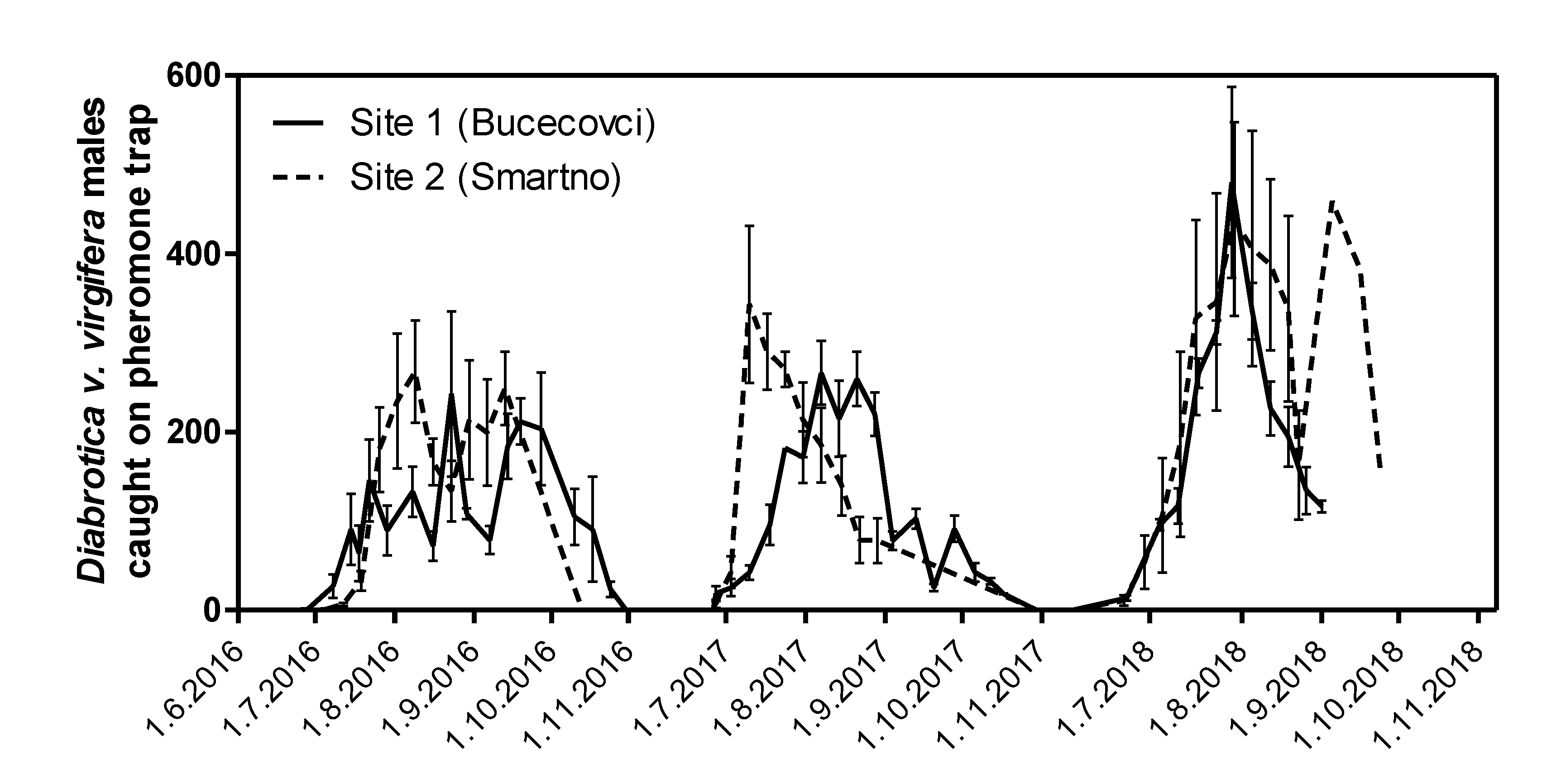
3.1. Insect Pest Density at Two Experimental Sites
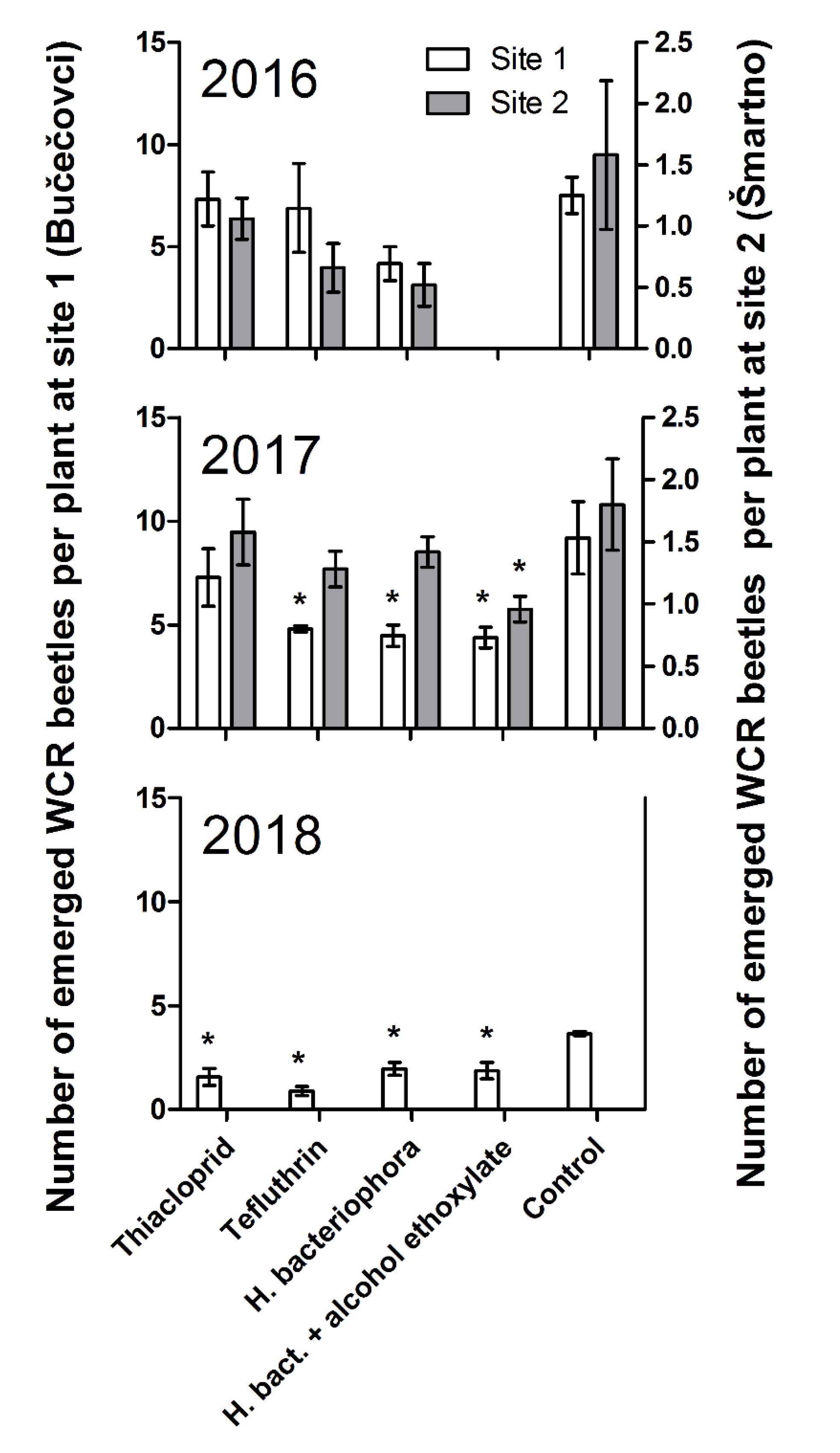
3.2. Treatment Efficacy
3.3. Evaluation of Maize Root Damage, Plant Lodging, Fresh Plant Weight, and Fresh Grain Yield Per Plant
3.4. Assessment of Persistence of Nematodes (EPN) in the Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gillette, C.P. Diabrotica virgifera LeConte as a corn rootworm. J. Econ. Entomol. 1912, 54, 364–366. [Google Scholar] [CrossRef]
- Ball, H.J.; Weekman, G.T. Insecticide Resistance in the Adult Western Corn Rootworm in Nebraska1. J. Econ. Entomol. 1962, 55, 439–441. [Google Scholar] [CrossRef]
- Mayo, Z.B. Field Evaluation of Insecticides for Control of Larvae of Corn Rootworms. In Methods for the Study of Pest Diabrotica; Springer: New York, NY, USA, 1986; pp. 183–203. [Google Scholar]
- Zhu, K.Y.; Wilde, G.E.; Sloderbeck, P.E.; Buschman, L.L.; Higgins, R.A.; Whitworth, R.J.; Bowling, R.A.; Starkey, S.R.; He, F. Comparative susceptibility of western corn rootworm (Coleoptera: Chrysomelidae) adults to selected insecticides in Kansas. J. Econ. Entomol. 2005, 98, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.E.; Souza, D.; Zukoff, S.N.; Meinke, L.J.; Siegfried, B.D. Cross-resistance and synergism bioassays suggest multiple mechanisms of pyrethroid resistance in western corn rootworm populations. PLoS ONE 2017, 12, e0179311. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, T.; Cavato, T.; Brar, G.; Coombe, T.; DeGooyer, T.; Ford, S.; Groth, M.; Howe, A.; Johnson, S.; Kolacz, K.; et al. A method of controlling corn rootworm feeding using a Bacillus thuringiensis protein expressed in transgenic maize. Crop Sci. 2005, 45, 931–938. [Google Scholar] [CrossRef]
- Meissle, M.; Pilz, C.; Romeis, J. Susceptibility of Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) to the entomopathogenic fungus Metarhizium anisopliae when feeding on Bacillus thuringiensis Cry3Bb1-expressing maize. Appl. Environ. Microbiol. 2009, 75, 3937–3943. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Keweshan, R.S.; Dunbar, M.W. Field-evolved resistance to Bt maize by Western corn rootworm. PLoS ONE 2011, 6, e22629. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Dunbar, M.W. Evidence of Resistance to Cry34 / 35Ab1 Corn by Western Corn Rootworm (Coleoptera : Chrysomelidae): Root Injury in the Field and Larval Survival in Plant-Based Bioassays. J. Econ. Entomol. 2016, 109, 1872–1880. [Google Scholar] [CrossRef]
- Kiss, J.; Edwards, C.R.; Berger, H.K.; Cate, P.; Cean, M.; Cheek, S.; Derron, J.; Festic, H.; Furlan, L.; Igrc-Barcic, J.; et al. Monitoring of western corn rootworm (Diabrotica virgifera virgifera LeConte) in Europe 1992–2003. In Western Corn Rootworm: Ecology and Management; CABI Publishing: Wallingford, UK, 2009. [Google Scholar]
- Sivcev, L.; Stankovic, S.; Kljajic, P.; Kostic, M.; Sivcev, I. Management of western corn rootworm (Diabrotica virgifera virgifera). Pestic. phytomedicina 2012, 27, 189–201. [Google Scholar] [CrossRef]
- Toepfer, S.; Knuth, P.; Glas, M.; Kuhlmann, U. Successful application of entomopathogenic nematodes for the biological control of western corn rootworm larvae in Europe—A mini review. In Julius-Kühn-Archiv; 2014; p. 59. [Google Scholar] [CrossRef]
- Prasifka, J.R.; Spencer, J.L.; Tinsley, N.A.; Estes, R.E.; Gray, M.E. Adult activity and oviposition of corn rootworms, Diabrotica spp. (Coleoptera: Chrysomelidae), in Miscanthus, corn and switchgrass. J. Appl. Entomol. 2013, 137, 481–487. [Google Scholar] [CrossRef]
- Gray, M.E.; Sappington, T.W.; Miller, N.J.; Moeser, J.; Bohn, M.O. Adaptation and Invasiveness of Western Corn Rootworm: Intensifying Research on a Worsening Pest. Annu. Rev. Entomol. 2009, 54, 303–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, L.; Canzi, S.; Di Bernardo, A.; Edwards, C.R. The ineffectiveness of insecticide seed coatings and planting-time soil insecticides as Diabrotica virgifera virgifera LeConte population suppressors. J. Appl. Entomol. 2006, 130, 485–490. [Google Scholar] [CrossRef]
- Van Rozen, K.; Ester, A. Chemical control of Diabrotica virgifera virgifera LeConte. J. Appl. Entomol. 2010, 134, 376–384. [Google Scholar] [CrossRef]
- Frank, D.L.; Kurtz, R.; Tinsley, N.A.; Gassmann, A.J.; Meinke, L.J.; Moellenbeck, D.; Gray, M.E.; Bledsoe, L.W.; Krupke, C.H.; Estes, R.E.; et al. Effect of Seed Blends and Soil-Insecticide on Western and Northern Corn Rootworm Emergence from mCry3A + eCry3.1Ab Bt Maize. J. Econ. Entomol. 2015, 108, 1260–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVries, T.A.; Wright, R.J. Evaluation of Liquid and Granular Insecticide Formulations at Planting for Larval Corn Rootworm Control, 2018B. Arthropod Manag. Tests 2019, 44, 1–2. [Google Scholar] [CrossRef]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- Cizej, M.R.; Žveplan, S.; Škerbot, I.; Persolja, J. Reducing population of the Western Corn Rootworm (Diabrotica virgifera virgifera) (Coleoptera: Chrysomelidae) with mating disruption. In Proceedings of the 13th Slovenian Conference on Plant Protection with International Participation, Rimske Toplice, Slovenija, 7–8 March 2017. [Google Scholar]
- Ivezić, M. Root compensation of seven maize hybrids due to western corn rootworm (Diabrotica virgifera virgifera LeConte) larval injury. Bulg. J. Agric. Sci. 2011, 17, 371–378. [Google Scholar]
- Schumann, M.; Toepfer, S.; Vemmer, M.; Patel, A.; Kuhlmann, U.; Vidal, S. Field evaluation of an attract and kill strategy against western corn rootworm larvae. J. Pest Sci. 2014, 87, 259–271. [Google Scholar] [CrossRef]
- Kuhlmann, U.; van der Burgt, W.A.C.M. Possibilities for biological control of the western corn rootworm, Diabrotica virgifera virgifera LeConte, in Central Europe. Biocontrol 1998, 19, 59N–68N. [Google Scholar]
- Toepfer, S.; Gueldenzoph, C.; Ehlers, R.-U.; Kuhlmann, U. Screening of entomopathogenic nematodes for virulence against the invasive western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) in Europe. Bull. Entomol. Res. 2005, 95, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Toepfer, S.; Peters, A.; Ehlers, R.-U.; Kuhlmann, U. Comparative assessment of the efficacy of entomopathogenic nematode species at reducing western corn rootworm larvae and root damage in maize. J. Appl. Entomol. 2008, 132, 337–348. [Google Scholar] [CrossRef]
- Toepfer, S.; Burger, R.; Ehlers, R.-U.; Peters, A.; Kuhlmann, U. Controlling western corn rootworm larvae with entomopathogenic nematodes: Effect of application techniques on plant-scale efficacy. J. Appl. Entomol. 2009, 134, 467–480. [Google Scholar] [CrossRef]
- Kahrer, A.; Pilz, C.; Heimbach, U.; Wechselberger, K.; Grabenweger, G. Five years of experience in biocontrol of the western corn rootworm, Diabrotica virgifera virgifera, by entomoparasitic nematodes. IOBC/WPRS Bull. 2016, 113, 81–85. [Google Scholar]
- Pilz, C.; Keller, S.; Kuhlmann, U.; Toepfer, S. Comparative efficacy assessment of fungi, nematodes and insecticides to Control western corn rootworm larvae in maize. Biocontrol 2009, 54, 671–684. [Google Scholar] [CrossRef]
- Laznik, Ž.; Tóth, T.; Lakatos, T.; Trdan, S. Heterorhabditis bacteriophora (Poinar)—The first member from Heterorhabditidae family in Slovenia. Acta Agric. Slov. 2009, 93, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Laznik, Z.; Trdan, S. Entomopathogenic Nematodes (Nematoda: Rhabditida) in Slovenia: From Tabula Rasa to Implementation into Crop Production Systems. In Insecticides—Advances in Integrated Pest Management; InTech: Rijeka, Croatia, 2011; pp. 627–656. [Google Scholar]
- Pilz, C.; Wegensteiner, R.; Keller, S. Selection of entomopathogenic fungi for the control of the western corn rootworm Diabrotica virgifera virgifera. J. Appl. Entomol. 2007, 131, 426–431. [Google Scholar] [CrossRef]
- Babendreier, D.; Jeanneret, P.; Pilz, C.; Toepfer, S. Non-target effects of insecticides, entomopathogenic fungi and nematodes applied against western corn rootworm larvae in maize. J. Appl. Entomol. 2015, 139, 457–467. [Google Scholar] [CrossRef]
- Rauch, H.; Steinwender, B.M.; Mayerhofer, J.; Sigsgaard, L.; Eilenberg, J.; Enkerli, J.; Zelger, R.; Strasser, H. Field efficacy of Heterorhabditis bacteriophora (Nematoda: Heterorhabditidae), Metarhizium brunneum (Hypocreales: Clavicipitaceae), and chemical insecticide combinations for Diabrotica virgifera virgifera larval management. Biol. Control 2017, 107, 1–10. [Google Scholar] [CrossRef]
- Toepfer, S.; Kurtz, B.; Kuhlmann, U. Influence of soil on the efficacy of entomopathogenic nematodes in reducing Diabrotica virgifera virgifera in maize. J. Pest Sci. 2010, 83, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, B.; Hiltpold, I.; Turlings, T.C.J.; Kuhlmann, U.; Toepfer, S. Comparative susceptibility of larval instars and pupae of the western corn rootworm to infection by three entomopathogenic nematodes. Biocontrol 2009, 54, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.K. Soil ecology. In Entomopathogenic Nematodes in Biological Control; CRC press: Boca Raton, FL, USA, 2018; ISBN 9781351080194. [Google Scholar]
- Klein, M.G. Efficacy against soil-inhabiting insect pests. In Entomopathogenic Nematodes in Biological Control; CRC press: Boca Raton, FL, USA, 2018; ISBN 9781351080194. [Google Scholar]
- Gaugler, R.; Han, R. Production technology. In Entomopathogenic Nematology; CABI: Wallingford, UK, 2009. [Google Scholar]
- Glazer, I. Survival biology. In Entomopathogenic Nematology; CABI: Wallingford, UK, 2009. [Google Scholar]
- Hoctor, T.L.; Gibb, T.J.; Bigelow, C.A.; Richmond, D.S. Survival and infectivity of the insect-parasitic nematode Heterorhabditis bacteriophora poinar in solutions containing four different turfgrass soil surfactants. Insects 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, H.; Zelger, R.; Strasser, H. Highly Efficient Field Emergence Trap for Quantitative Adult Western Corn Rootworm Monitoring. J. Kansas Entomol. Soc. 2016, 89, 256–266. [Google Scholar] [CrossRef]
- Oleson, J.D.; Park, Y.L.; Nowatzki, T.M.; Tollefson, J.J. Node-injury scale to evaluate root injury by corn rootworms (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2005, 98, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. The ‘Galleria bait method’ for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Woodring, J.L.; Kaya, H.K. Steinernematid and Heterorhabditid Nematodes: A Handbook of Techniques; Arkansas Agricultural Experiment Station: Fayetteville, AR, USA, 1988. [Google Scholar]
- Modic, Š.; Knapič, M.; Razinger, J.; Urek, G. Applicability of the prognostic model to predict the western corn rootworm (Diabrotica virgifera) and influence of crop rotation to increase of WCR population in Slovenia. In Proceedings of the Zbornik predavanj in referatov 12. Slovenskega posvetovanja o varstvu rastlin z mednarodno udeležbo, Ptuj, Slovenija, 3–4 March 2015. [Google Scholar]
- Toepfer, S.; Zellner, M. Entomopathogenic nematodes can replace soil insecticides in western corn rootworm control. IOBC-WPRS Bull. 2017, 129, 144–156. [Google Scholar]
- Souza, D.; Peterson, J.A.; Wright, R.J.; Meinke, L.J. Field efficacy of soil insecticides on pyrethroid-resistant western corn rootworms (Diabrotica virgifera virgifera LeConte). Pest Manag. Sci. 2019, 76, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Furlan, L.; Vasileiadis, V.P.; Chiarini, F.; Huiting, H.; Leskovšek, R.; Razinger, J.; Holb, I.J.; Sartori, E.; Urek, G.; Verschwele, A.; et al. Risk assessment of soil-pest damage to grain maize in Europe within the framework of Integrated Pest Management. Crop Prot. 2016, 97, 52–59. [Google Scholar] [CrossRef]
- Calles-Torrez, V.; Knodel, J.J.; Boetel, M.A.; Doetkott, C.D.; Podliska, K.K.; Ransom, J.K.; Beauzay, P.; French, B.W.; Fuller, B.W. Transgenic Bt Corn, Soil Insecticide, and Insecticidal Seed Treatment Effects on Corn Rootworm (Coleoptera: Chrysomelidae) Beetle Emergence, Larval Feeding Injury, and Corn Yield in North Dakota. J. Econ. Entomol. 2018, 111, 348–360. [Google Scholar] [CrossRef]
- Boetel, M.A.; Fuller, B.W.; Evenson, P.D. Emergence of Adult Northern and Western Corn Rootworms (Coleoptera: Chrysomelidae) Following Reduced Soil Insecticide Applications. J. Econ. Entomol. 2003, 96, 714–729. [Google Scholar] [CrossRef]
- Gray, M.E.; Felsot, A.S.; Steffey, K.L.; Levine, E. Planting Time Application of Soil Insecticides and Western Corn Rootworm (Coleoptera: Chrysomelidae) Emergence: Implications for Long-Term Management Programs. J. Econ. Entomol. 1992, 85, 544–553. [Google Scholar] [CrossRef]
- Harris, C.R. Factors Influencing the Effectiveness of Soil Insecticides. Annu. Rev. Entomol. 1972, 17, 177–198. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E.M. Soil moisture effects on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, S. glaseri, Heterorhabditis zealandica, and H. bacteriophora. Appl. Soil Ecol. 2007, 35, 128–139. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E.M. Effect of soil type on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, Steinernema glaseri, Heterorhabditis zealandica, and Heterorhabditis bacteriophora. J. Invertebr. Pathol. 2006, 92, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Susurluk, A. Establishment and Persistence of the Entomopathogenic Nematodes, Steinernema Feltiae and Heterorhabditis Bacteriophora. Ph.D. Thesis, Christian-Albrechts Universität Kiel, Kiel, Germany, 2005. [Google Scholar]
- Kurtz, B.; Toepfer, S.; Ehlers, R.U.; Kuhlmann, U. Assessment of establishment and persistence of entomopathogenic nematodes for biological control of western corn rootworm. J. Appl. Entomol. 2007, 131, 420–425. [Google Scholar] [CrossRef]
- Brust, G.E. Augmentation of an endemic entomogenous nematode by agroecosystem manipulation for the control of a soil pest. Agric. Ecosyst. Environ. 1991, 36, 175–184. [Google Scholar] [CrossRef]
- Riga, E.; Whistlecraft, J.; Potter, J. Potential of controlling insect pests of corn using entomopathogenic nematodes. Can. J. Plant Sci. 2001, 81, 783–787. [Google Scholar] [CrossRef]
- Blandino, M.; Ferracini, C.; Rigamonti, I.; Testa, G.; Saladini, M.A.; Jucker, C.; Agosti, M.; Alma, A.; Reyneri, A. Control of western corn rootworm damage by application of soil insecticides at different maize planting times. Crop Prot. 2017, 93, 19–27. [Google Scholar] [CrossRef]
- Tinsley, N.A.; Estes, R.E.; Schrader, P.M.; Gray, M.E. Evaluating multiple approaches for managing western corn rootworm larvae with seed blends. J. Appl. Entomol. 2015, 139, 76–86. [Google Scholar] [CrossRef]
- Johnson, K.D.; Campbell, L.A.; Lepping, M.D.; Rule, D.M. Field Trial Performance of Herculex XTRA (Cry34Ab1/Cry35Ab1) and SmartStax (Cry34Ab1/Cry35Ab1 + Cry3Bb1) Hybrids and Soil Insecticides Against Western and Northern Corn Rootworms (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2017, 110, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, C.S.; Schroeder, P.C.; Shields, E.J. Vertical distribution, persistence, and activity of entomopathogenic nematodes (Nematoda: Heterorhabditidae and Steinernematidae) in alfalfa snout beetle- (Coleoptera: Curculionidae) infested fields. Environ. Entomol. 1995, 24, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Smits, P.H. Post-application persistence of entomopathogenic nematodes. Biocontrol Sci. Technol. 1996, 6, 379–388. [Google Scholar] [CrossRef]
- Schaub, L.; Furlan, L.; Tóth, M.; Steinger, T.; Carrasco, L.R.; Toepfer, S. Efficiency of pheromone traps for monitoring Diabrotica virgifera virgifera LeConte. EPPO Bull. 2011, 41, 189–194. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Field | Site 1: Eastern Slovenia, Bučečovci | Site 2: Northern Slovenia, Šmartno Pri Cerkljah |
|---|---|---|
| Experiment location | 46°35’07"N 16°06’37"E | 46°15’08.8"N 14°29’54.7"E |
| Date of sowing/nematode application | 22 April 2016 | 11 May 2016 |
| 26 April 2017 | 17 May 2017 | |
| 9 May 2018 | ||
| Maize seed | Chapalu (Agrosaat) | LG 34.90 (Agrosaat) |
| Field size | 0.14 ha | 0.14 ha |
| Soil types | silty loam texture | sandy loam texture |
| Field Site | Date of Sowing and Application | Treatment Active Ingredient (Tradename) | Dose |
|---|---|---|---|
| 1 | 22 April 2016 | 1. Untreated maize seed (Control) | 85,000 plants/ha |
| 2. H. bacteriophora (Dianem) | 2 × 109 nematodes/ha with 200 L water | ||
| 3. Tefluthrin (Force 1,5 G) | 12 kg/ha (1.5% a.i.) | ||
| 4. Thiacloprid (Sonido FS 400) | 0.125 L/50,000 seeds (40% a.i.) | ||
| 26 April 2017 | 1. Untreated maize seed (Control) | 85,000 plants/ha | |
| 2. H. bacteriophora (Dianem) | 2 × 109 nematodes/ha with 200 L water | ||
| 3. Tefluthrin (Force 1,5 G) | 12 kg/ha (1.5% a.i.) | ||
| 4. Thiacloprid (Sonido FS 400) | 0.125 L/50,000 seeds (40% a.i.) | ||
| 5. H. bacteriophora + alcohol ethoxylate (Dianem + Transformer) | 5 L/ha, 20% w/w | ||
| 9 May 2018 | 1. Untreated maize seed (Control) | 85,000 plants/ha | |
| 2. H. bacteriophora (Dianem) | 2 × 109 nematodes/ha with 200 L water | ||
| 3. Tefluthrin (Force 1,5 G) | 12 kg/ha (1.5% a.i.) | ||
| 4. Thiacloprid (Sonido FS 400) | 0.125 L/50,000 seeds (40% a.i.) | ||
| 5. H. bacteriophora + alcohol ethoxylate (Dianem + Transformer) | 5 L/ha, 20% w/w | ||
| 2 | 11 May 2016 | 1. Untreated maize seed (Control) | 85,000 plants/ha |
| 2. H. bacteriophora (Dianem) | 2 × 109 nematodes /ha with 200 L water | ||
| 3. Tefluthrin (Force 1,5 G) | 12 kg/ha (1.5% a.i.) | ||
| 4. Thiacloprid (Sonido FS 400) | 0.125 L/50,000 seeds (40% a.i.) | ||
| 17 May 2017 | 1. Untreated maize seed (Control) | 85,000 plants/ha | |
| 2. H. bacteriophora (Dianem) | 2 × 109 nematodes /ha with 200 L water | ||
| 3. Tefluthrin (Force 1,5 G) | 12 kg/ha (1.5% a.i.) | ||
| 4. Thiacloprid (Sonido FS 400) | 0.125 L/50,000 seeds (40% a.i.) | ||
| 5. H. bacteriophora + alcohol ethoxylate (Dianem + Transformer) | 5 L/ha, 20% w/w |
| Year | Site/Treatment | Thiacloprid | Tefluthrin | H. bacteriophora | H. bacteriophora + Alcohol Ethoxylate | Control |
|---|---|---|---|---|---|---|
| 2016 | 1 | 2.7% | 8.4% | 44.7% | nd | 0.0% |
| 2 | 33.0% | 58.5% | 67.0% | nd | 0.0% | |
| 2017 | 1 | 20.9% | 47.7% | 51.5% | 52.5% | 0.0% |
| 2 | 12.0% | 29.6% | 21.3% | 46.3% | 0.0% | |
| 2018 | 1 | 57.2% | 76.0% | 46.6% | 48.8% | 0.0% |
| Average efficacy ± SE | 25.2 ± 9.4% | 44.1 ± 11.7% | 46.2 ± 7.4% | 49.2 ± 1.8% | 0.0 ± 0.0% | |
| HSD Multiple Range test | AB | A | A | A | B | |
| Treatment | Weight of 1 Maize Plant [g] | Fresh Grain Yield Per Plant [g] | |||
|---|---|---|---|---|---|
| Site 1 | Site 2 | HSD test | Site 1 | Site 2 | |
| Thiacloprid | 557 ± 36 | 1033 ± 36 | A | 246 ± 13 | 395 ± 13 |
| Tefluthrin | 490 ± 36 | 956 ± 36 | AB | 222 ± 13 | 365 ± 13 |
| H. bacteriophora | 523 ± 36 | 1021 ± 36 | A | 240 ± 13 | 382 ± 13 |
| H. bacteriophora + alcohol ethoxylate | 561 ± 36 | 1011 ± 36 | A | 259 ± 13 | 385 ± 19 |
| Control | 440 ± 36 | 1021 ± 36 | B | 216 ± 13 | 369 ± 13 |
| Site/Year | Days Post Treatment | Treatment/Depth of Sampling [cm] | |||
|---|---|---|---|---|---|
| H. bacteriophora | H. bacteriophora + Alcohol Ethoxylate | ||||
| 5–10 | 10–15 | 5–10 | 10–15 | ||
| Site 1 | |||||
| 2016 | 26 | 78 | 89 | nd | nd |
| 61 | 11 | 22 | nd | nd | |
| 83 | 11 | 33 | nd | nd | |
| 2017 | 30 | 73 | 37 | 50 | 43 |
| 57 | 0 | 0 | 10 | 37 | |
| 83 | 50 | 63 | 90 | 83 | |
| 113 | 8 | 8 | 25 | 13 | |
| 141 | 4 | 8 | 4 | 0 | |
| 2018 | 20 | 13 | 27 | 40 | 27 |
| 41 | 13 | 7 | 7 | 13 | |
| 57 | 0 | 0 | 0 | 22 | |
| 69 | 0 | 7 | 0 | 0 | |
| 83 | 13 | 13 | 3 | 3 | |
| 100 | 13 | 0 | 0 | 17 | |
| 128 | 0 | 0 | 13 | 0 | |
| Site 2 | |||||
| 2016 | 28 | 0 | 33 | nd | nd |
| 49 | 0 | 0 | nd | nd | |
| 84 | 0 | 0 | nd | nd | |
| 2017 | 22 | 4 | 0 | 7 | 11 |
| 57 | 33 | 10 | 60 | 50 | |
| 114 | 8 | 4 | 13 | 8 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modic, Š.; Žigon, P.; Kolmanič, A.; Trdan, S.; Razinger, J. Evaluation of the Field Efficacy of Heterorhabditis Bacteriophora Poinar (Rhabditida: Heterorhabditidae) and Synthetic Insecticides for the Control of Western Corn Rootworm Larvae. Insects 2020, 11, 202. https://doi.org/10.3390/insects11030202
Modic Š, Žigon P, Kolmanič A, Trdan S, Razinger J. Evaluation of the Field Efficacy of Heterorhabditis Bacteriophora Poinar (Rhabditida: Heterorhabditidae) and Synthetic Insecticides for the Control of Western Corn Rootworm Larvae. Insects. 2020; 11(3):202. https://doi.org/10.3390/insects11030202
Chicago/Turabian StyleModic, Špela, Primož Žigon, Aleš Kolmanič, Stanislav Trdan, and Jaka Razinger. 2020. "Evaluation of the Field Efficacy of Heterorhabditis Bacteriophora Poinar (Rhabditida: Heterorhabditidae) and Synthetic Insecticides for the Control of Western Corn Rootworm Larvae" Insects 11, no. 3: 202. https://doi.org/10.3390/insects11030202