Biomorphological Characterization of Brazilian Capsicum Chinense Jacq. Germplasm

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotyping
2.2. Genotyping
2.3. Data Analysis
3. Results
3.1. Morpho-Agronomic Characterization Data
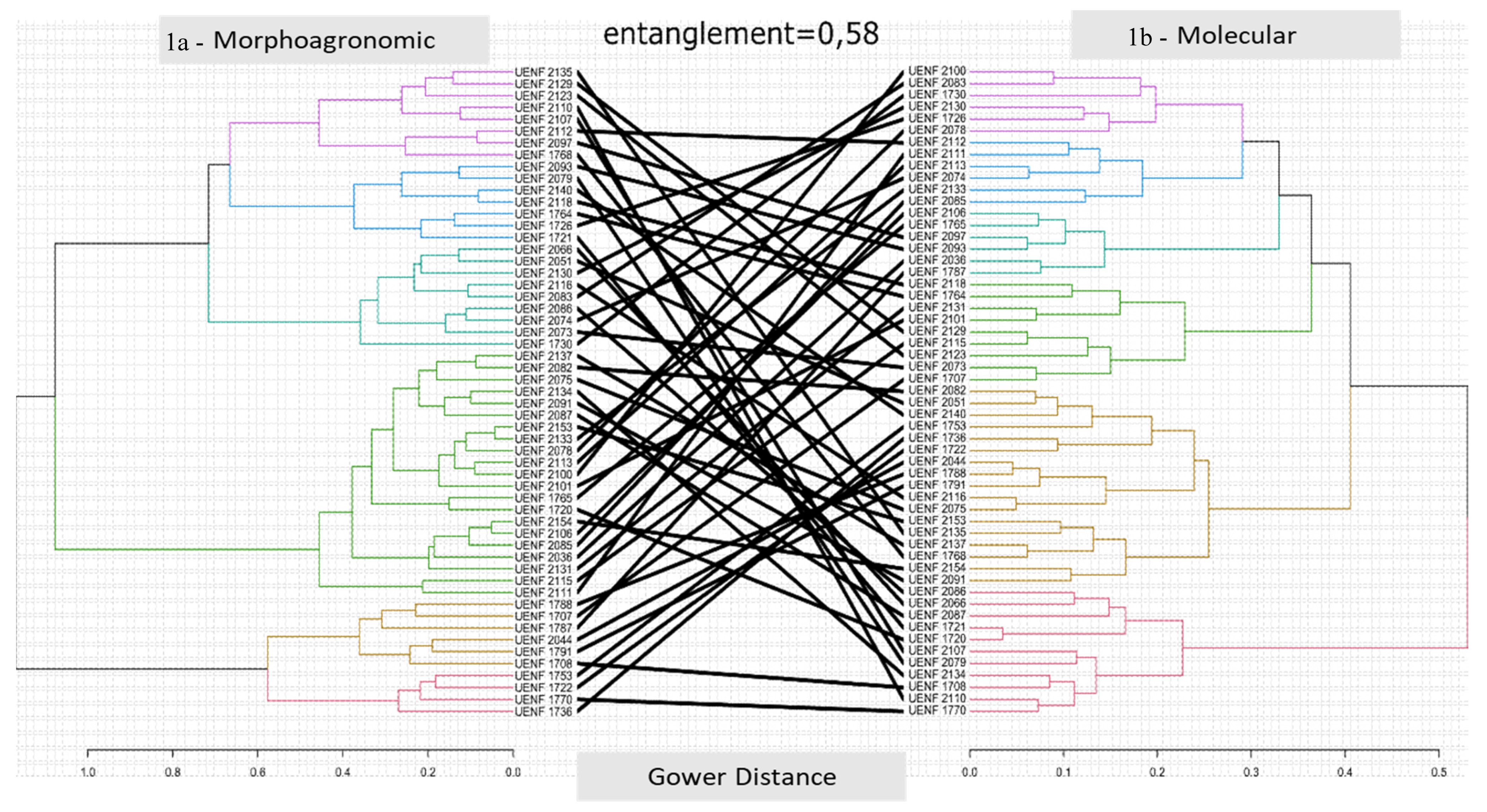
3.2. Morpho-Agronomic Diversity
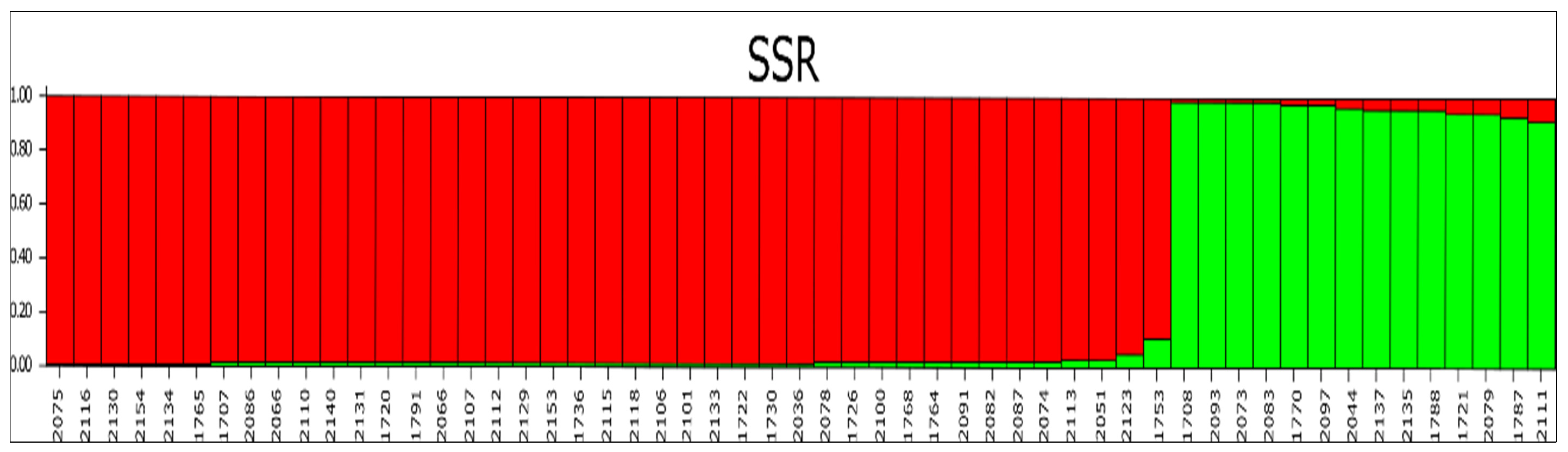
3.3. Molecular Characterization Data
3.4. Molecular Diversity
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hulse-Kemp, A.M.; Ashrafi, H.; Plieske, J.; Lemm, J.; Stoffel, K.; Hill, T.; Luerssen, H.; Pethiyagoda, C.L.; Lawley, C.T.; Ganal, M.W. A HapMap leads to a Capsicum annuum SNP infinium array: A new tool for pepper breeding. Hortic. Res. 2016, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosland, P.W. Capsicum: Innovative uses of an ancient crop. In Progress in New Crops; Janick, J., Ed.; ASHS Press: Arlington, VA, USA, 1996; pp. 479–487. [Google Scholar]
- Neitzke, R.S. Genetic Resources of Capsicum Peppers—Exploring the Multiplicity of Uses. Tese (Doutorado em Agronomia); Universidade Federal de Pelotas: Pelotas, Brazil, 2012. [Google Scholar]
- Neitzke, R.S.; Fischer, S.Z.; Vasconcelos, C.S.; Barbieri, R.L.; Treptow, R.O. Pimentas ornamentais: Aceitação e preferências do público consumidor. Hortic. Bras. 2016, 34, 102–109. [Google Scholar] [CrossRef]
- Barboza, G.E.; Garcia, C.C.; Gonzalez, S.L.; Scaldaferro, M.; Reyes, X. Four new species of Capsicum (Solanaceae) from the tropical Andes and an update on the phylogeny of the genus. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Plant List. Available online: http://www.theplantlist.org (accessed on 10 December 2019).
- DeWitt, D.; Bosland, P.W. The Complete Chilli Pepper Book: A Gardener’s Guide to Choosing, Growing, Preserving and Cooking; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Barboza, G.E.; Bianchetti, L.D.B. Three New Species of Capsicum (Solanaceae) and a Key to the Wild Species from Brazil. Am. Soc. Plant Taxon. 2005, 30, 863–871. [Google Scholar] [CrossRef]
- Moscone, E.A.; Scaldaferro, M.A.; Gabriele, M.; Cecchini, N.M.; García, Y.S.; Jarret, R.; Daviña, J.R.; Ducasse, D.A.; Barboza, G.E.; Ehrendorfer, F. The Evolution of Chili Peppers (Capsicum—Solanaceae): A Cytogenetic Perspective. In Proceedings of the 6th International Solanaceae Conference, Madison, WI, USA, 23–27 July 2006; Spooner, D.M., Ed.; Volume 745, pp. 137–170. [Google Scholar]
- Pickersgill, B. Cytogenetics and evolution of Capsicum L. In Chromosome Engineering in Plants: Genetics, Breeding, Evolution, Part B; Tsuchiya, T., Gupta, P.K., Eds.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 139–160. [Google Scholar]
- Moses, M.; Umaharan, P. Genetic structure and phylogenetic relationships of Capsicum Chinese. J. Am. Soc. Hortic. Sci. 2012, 137, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Moses, M.; Umaharan, P.; Dayanandan, S. Microsatellite based analysis of the genetic structure and diversity of Capsicum chinense in the Neotropics. Genet. Resour. Crop. Evolut. 2014, 61, 741–755. [Google Scholar] [CrossRef]
- Carvalho, S.I.C.; Ragassi, C.F.; Bianchetti, L.B.; Reifschneider, F.J.B.; Buso, G.S.C.; Faleiro, F.G. Morphological and genetic relationships between wild and domesticated forms of peppers (Capsicum frutescens L. and C. chinense Jacquin). Genet. Mol. Res. 2014, 13, 7447–7464. [Google Scholar] [CrossRef] [Green Version]
- Baba, V.Y.; Rocha, K.R.; Gomes, G.P.; Ruas, C.F.; Ruas, P.M.; Rodrigues, R.; Gonçalves, L.S.A. Genetic diversity of Capsicum chinense accessions based on fruit morphological characterization and AFLP markers. Genet. Resour. Crop Evolut. 2016, 63, 1371–1381. [Google Scholar] [CrossRef]
- Moreira, A.F.P.; Ruas, P.M.; Ruas, C.F.; Baba, V.Y.; Giordani, W.; Arruda, I.M.; Rodrigues, R.; Gonçalves, L.S.A. Genetic diversity, population structure and genetic parameters of fruit traits in Capsicum Chinese. Sci. Hortic. 2018, 236, 1–9. [Google Scholar] [CrossRef]
- Nass, L.L.; Sigrist, M.S.; Ribeiro, C.S.C.; Reifschneider, F.J.B. Genetic resources: The basis for sustainable and competitive plant breeding. Crop Breed. Appl. Biotechnol. 2012, 12, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Ballina-Gomez, H.; Ruiz-Sanchez, E.; Chan-Cupul, W.; Latournerie-Moreno, L.; Hernandez-Alvarado, L.; Islas-Flores, I.; Zuñiga-Aguilar, J.J. Response of Bemisia tabaci Genn. (Hemiptera: Aleyrodidae) Biotype B to Genotypes of Pepper Capsicum annuum (Solanales: Solanaceae). Neotrop. Entomol. 2013, 42, 205. [Google Scholar] [CrossRef]
- Nicolai, M.; Cantet, M.; Lefebvre, V.; Sage-Palloix, A.M.; Palloix, A. Genotyping a large collection of pepper (Capsicum spp.) with SSR loci brings new evidence for the wild origin of cultivated C. annuum and the structuring of genetic diversity by human selection of cultivar types. Genet. Resour. Crop Evolut. 2013, 60, 2375–2390. [Google Scholar] [CrossRef]
- Quang, D.V.; Schreinemachers, P.; Berger, T. Ex-ante assessment of soil conservation methods in the uplands of Vietnam: An agent-based modeling approach. Agric. Syst. 2015, 123, 108–119. [Google Scholar] [CrossRef]
- Sudré, C.P.; Gonçalves, L.S.A.; Rodrigues, R.; Amaral Júnior, A.T.; Riva-Souza, E.M.; Bento, C.S. Genetic variability in domesticated Capsicum spp. as assessed by morphological and agronomic data in mixed statistical analysis. Genet. Mol. Res. 2010, 9, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Santos, M. Metamorfoses do Espaço Habitado; EDUSP: São Paulo, Brazil, 2014. [Google Scholar]
- Newman, P. The environmental impact of cities. Environ. Urban. 2006, 18, 275–295. [Google Scholar] [CrossRef]
- Ribeiro, L.S. A Relevância Dos Patrimônios Naturais Da Humanidade E Das Reservas Da Biosfera Para a Conservação Da Biodiversidade—O Caso Do Parque Nacional Da Chapada Dos Veadeiros (PNCV) Monografia De Especialização—Universidade De Brasília; Centro UnB Cerrado: Alto Paraíso, Brazil, 2018. [Google Scholar]
- Organização das Nações Unidas. Available online: https://nacoesunidas.org/pos2015/agenda2030/ (accessed on 14 December 2019).
- Araújo, R.F.; Abud, H.F.; Silva, L.J.; Araújo, E.F.; Pinto, C.M.F.; Silva, F.W.S. Alterações fisiológicas e atividade de enzimas antioxidantes em sementes de pimentas Capsicum chinense Jacq e Capsicum frutescens L. durante o processo de maturação. Rev. Ceres 2018, 65, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Zonneveld, M.; Ramirez, M.; Williams, D.E.; Petz, M.; Meckelmann, S.; Avila, T.; Bejarano, C.; Ros, L.; Pea, K.; Jger, M. Screening Genetic Resources of Capsicum Peppers in Their Primary Center of Diversity in Bolivia and Peru. PLoS ONE 2015, 10, e0134663. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, P.A.; Dutra, I.P.; Moulin, M.M.; Santos, J.O.; Júnior, A.C.S. Morphological characterization and analysis of genetic variability among pepper accessions. Ciência Rural 2016, 46, 1151–1157. [Google Scholar] [CrossRef]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef]
- Grover, A.; Sharma, P. Development and use of molecular markers: Past and present. Crit. Rev. Biotechnol. 2014, 36, 1–13. [Google Scholar] [CrossRef]
- Finger, F.L.; Lannes, S.D.; Schuelter, A.R.; Doege, J.; Comerlato, A.P.; Gonçalves, L.S.A.; Ferreira, F.R.A.; Clovis, L.R.; Scapim, C.A. Genetic diversity of Capsicum chinense (Solanaceae) accessions based on molecular markers and morphological and agronomic traits. Genet. Mol. Res. 2010, 9, 1852–1864. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.R.; Tikunov, Y.; de Vos, R.C.H.; Pelgrom, K.T.B.; Maharijaya, A.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolomics and molecular marker analysis to explore pepper (Capsicum sp.) biodiversity. Metabolomics 2013, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Thul, S.; Darokar, M.; Shasany, A.; Khanuja, S. Molecular Profiling for Genetic Variability in Capsicum Species Based on ISSR and RAPD Markers. Mol. Biotechnol. 2011, 51, 137–147. [Google Scholar] [CrossRef]
- Pacheco-Olvera, A.; Hernandez-Verdugo, S.; Rocha-Ramirez, V.; Gonzalez-Rodriguez, A.; Oyama, K. Genetic Diversity and Structure of Pepper (Capsicum Annuum L.) from Northwestern Mexico Analyzed by Microsatellite Markers. Crop Sci. 2012, 52, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.P.; Kumar, R.; Kumar, S.; Rai, A.; Kumar, S.; Singh, M.; Singh, S.P.; Rai, A.B.; Paliwal, R. Genetic diversity in Capsicum germplasm based on microsatellite and random amplified microsatellite polymorphism markers. Physiol. Mol. Biol. Plants 2013, 19, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.I.C.; Ragassi, C.F.; Oliveira, I.B.; Amaral, Z.P.S.; Reifschneider, F.J.B.; Faleiro, F.G.; Buso, G.S.C. Transferability of microsatellite markers of Capsicum annuum L. to C. frutescens L. and C. chinense Jacq. Genet. Mol. Res. 2015, 14, 7937–7946. [Google Scholar] [CrossRef]
- Cardoso, R.; Ruas, C.F.; Giacomin, R.M.; Ruas, P.M.; Ruas, E.A.; Barbieri, R.L.; Rodrigues, R.; Gonçalves, L.S.A. Genetic variability in Brazilian Capsicum baccatum germplasm collection assessed by morphological fruit traits and AFLP markers. PLoS ONE 2018, 13, e0196468. [Google Scholar] [CrossRef] [Green Version]
- Jarret, R.L.; Berke, T. Variation for Fruit Morphological Characteristics in a Capsicum chinense Jacq. Germplasm Collection. HortScience 2008, 43, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Bharath, S.M.; Cilas, C.; Umaharan, P. Fruit Trait Variation in a Caribbean Germplasm Collection of Aromatic Hot Peppers (Capsicum chinense Jacq.). HortScience 2013, 48, 531–538. [Google Scholar] [CrossRef] [Green Version]
- IPGRI. Descriptors for Capsicum (Capsicum spp.); International Plant Genetic Resources Institute: Rome, Italy, 1995; 49p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1987, 12, 13. [Google Scholar]
- Daher, R.F.; Pereira, M.G.; Tupinamba, E.A.; Amaral Júnior, A.T.; Aragão, W.N.; Ribeiro, F.E.; Oliveira, L.O.; Sakiyama, N.S. Assessment of coconut tree genetic divergence by compound sample RAPD marker analysis. Crop Breed. Appl. Biotechnol. 2002, 3, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Minamiyama, Y.; Tsuro, M.; Hirai, M. An SSR-basead linkage map of Capsicum annuum. Mol. Breed. 2006, 18, 157–169. [Google Scholar] [CrossRef]
- Singh, D. The relative importance of characters affecting genetic divergence. Indian J. Genet. Plant Breed. 1981, 41, 237–245. [Google Scholar]
- Cruz, C.D. GENES—A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. 2013, 35, 271–276. [Google Scholar]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–874. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and ENVIRONMENT for statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 9 May 2019).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Cheema, D.S.; Dhaliwa, M.S.; Garg, N. Heterosis and 51 combining ability for earliness, plant growth, yield and fruit attributes in hot pepper (Capsicum annuum L.) Involving genetic and cytoplasmic-genetic male sterile lines. Sci. Hortic. 2014, 168, 175–188. [Google Scholar] [CrossRef]
- Castro, S.P.; Dávila, A.M.A.G. Caracterización morfológica de 93 accesiones de Capsicum spp del banco de germoplasma de la Universidad Nacional de Colombia—Sede Palmira. Acta Agron. 2008, 57, 247–252. [Google Scholar]
- Lannes, S.D.; Finger, F.L.; SchuelteR, A.R.; Casali, V.W.D. Growth and quality of Brazilian accessions of Capsicum chinense fruits. Sci. Hortic. 2007, 112, 266–270. [Google Scholar] [CrossRef]
- Domenico, C.I.; Coutinho, J.P.; Godoy, H.T.; Melo, A.M.T. Caracterização agronômica e pungência em pimenta de cheiro. Hortic. Bras. 2012, 30, 466–472. [Google Scholar] [CrossRef] [Green Version]
- Rêgo, E.R.; Rêgo, M.M.; Matos, I.W.F.; Barbosa, L.A. Morphological and chemical characterization of fruits of Capsicum spp. accessions. Hortic. Bras. 2011, 29, 3. [Google Scholar] [CrossRef] [Green Version]
- Ferrão, L.F.V.; Cecon, P.R.; Finger, F.L.; Silva, F.F.; Puiatti, M. Divergência genética entre genótipos de pimenta com base em caracteres morfo-agrônomicos. Hortic. Bras. 2011, 29, 354–358. [Google Scholar] [CrossRef]
- Pessoa, A.M.S.; Rêgo, E.R.; Santos, C.A.P.; Carvalho, M.G.; Rêgo, M.M. Inheritance of seedling and plant traits in ornamental pepper (Capsicum annuum). Genet. Mol. Res. 2019, 18, 18120. [Google Scholar] [CrossRef]
- Agyare, R.Y.; Akromah, R.; Abdulai, M.S. Assesment of genetic diversity in pepper (Capsicum spp.) landraces from Ghana using agro-morphological characters. Am. J. Exp. Agric. 2016, 12, 1–16. [Google Scholar]
- Ghosh, K.P.; Islam, A.K.M.A.; Mian, M.A.K.; Hossain, M.M. Variability and character association in F2 segregating population of different commercial hybrids of tomato (Solanum lycopersicum L.). J. Appl. Sci. Environ. Manag. 2010, 14, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Andrade Júnior, V.C.; Pedrosa, C.E.; Miranda, T.G.; Valadares, N.R.; Pereira, S.L.; Azevedo, A.M. Biometric evaluation of morpho-agronomic traits in pepper lines and hybrids. Hortic. Bras. 2018, 36, 357–361. [Google Scholar] [CrossRef]
- Peña-Yam, L.P.; Muñoz-Ramírez, L.S.; Avilés-Viñas, S.A.; Canto-Flick, A.; Pérez-Pastrana, J.; Guzmán-Antonio, A.; Santana-Buzzy, N.; Aguilera-Cauich, E.A.; Mijangos-Cortes, J.O. Analysis of Genetic Parameters of Habanero Pepper (Capsicum chinense Jacq.) in the Yucatan, Mexico. HortScience 2019, 54, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Naegele, R.P.; Mitcheil, J.; Hausbeck, M.K. Genetic diversity, population structure, and heritability of fruits traits in Capsicum annuum. PLoS ONE 2016, 11, e0156969. [Google Scholar] [CrossRef]
- Moulin, M.M.; Rodrigues, R.; Ramos, H.C.C.; Bento, C.S.; Sudré, C.P.; Gonçalves, L.S.A.; Viana, A.P. Construction of an integrated genetic map for Capsicum baccatum L. Genet. Mol. Res. 2015, 14, 6683–6694. [Google Scholar] [CrossRef]
- Campos, A.L. Coleta e Caracterização de Acessos de Capsicum spp. Da Agrobiodiversidade Da Região Sudoeste Do Mato Grosso. Master’s Thesis, Universidade do Estado do Mato Grosso, Mato Grosso, Brazil, 2014. [Google Scholar]
- Mimura, Y.; Inoue, T.; Minamiyama, Y.; Kubo, N. An SSR-based genetic map of pepper (Capsicum annuum L.) serves as an anchor for the alignment of major pepper maps. Breed. Sci. 2012, 62, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, M.S.; Yadav, A.; Jindal, S.K. Molecular Characterization and Diversity Analysis in Chilli Pepper Using Simple Sequence Repeats (SSR) Markers. Afr. J. Biotechnol. 2014, 13, 3137–3143. [Google Scholar]
- Meng, C.Y.; Wei, X.C.; Zhao, Y.Y.; Yuan, Y.X.; Yang, S.J.; Wang, Z.Y.; Zhang, X.W.; Sun, J.W.; Yang, L.; Yao, Q.J.; et al. Genetic diversity analysis of Capsicum genus by SSR markers. Mol. Plant Breed. 2017, 8, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.R.; Pereira, T.N.S.; Vitória, A.P.; Campos, K.P.; Rodrigues, R.; Silva, D.H.; Pereira, M.G. Genetic diversity among Capsicum accessions using RAPD markers. Crop Breed. Appl. Biotechnol. 2006, 6, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Leite, P.S.S.; Rodrigues, R.; Silva, R.N.O.; Pimenta, S.; Medeiros, A.M.; Bento, C.S.; Gonçalves, L.S.A. Molecular and agronomic analysis of intraspecific variability in Capsicum baccatum var. pendulum accessions. Genet. Mol. Res. 2016, 15, 4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Quantitative Traits | Relative Contribution (%) | Minimum Value | Maximum Value | Average Value |
|---|---|---|---|---|
| Fruit diameter (mm) | 9.83% | 8.99 | 34.44 | 21.22 |
| Days for germination | 9.1% | 5 | 11 | 8 |
| Fruit weight (g) | 8.84% | 1.04 | 18.61 | 6.93 |
| Pericarp thickness (mm) | 7.76% | 1.38 | 3.08 | 2.33 |
| Cotyledon leaf diameter (mm) | 7.38% | 6.04 | 8.05 | 6.97 |
| Fruit length (mm) | 6.77% | 7.85 | 84.93 | 34.32 |
| Leaf length (cm) | 6.67% | 6.5 | 9.5 | 7.57 |
| Plant height (cm) | 6.59% | 49 | 156 | 84.85 |
| Stem diameter (cm) | 5.77% | 0.96 | 2.08 | 1.35 |
| Leaf width (cm) | 5.52% | 3.8 | 5.9 | 4.18 |
| Days for maturation | 5.46% | 135 | 217 | 158 |
| Canopy diameter (cm) | 5.12% | 44.18 | 123.25 | 70.46 |
| Days for flowering | 4.27% | 62 | 92 | 75 |
| Peduncle length (mm) | 4.22% | 19 | 43 | 24.37 |
| Days for fruiting | 3.79% | 62 | 117 | 92 |
| Cotyledon leaf length (mm) | 2.85% | 12.81 | 25.96 | 16.82 |
| Genotype | MAF | Na | I | Ho | F |
|---|---|---|---|---|---|
| UENF 1707 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 1708 | 0.50 | 2 | 0.50 | 0.33 | 0.38 |
| UENF 1720 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 1721 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 1722 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 1726 | 0.78 | 2 | 0.35 | 0.22 | 0.41 |
| UENF 1730 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 1736 | 0.78 | 2 | 0.35 | 0 | 1.00 |
| UENF 1753 | 0.56 | 2 | 0.49 | 0.44 | 0.16 |
| UENF 1764 | 0.78 | 2 | 0.35 | 0 | 1.00 |
| UENF 1765 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 1768 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 1770 | 0.50 | 2 | 0.50 | 0.11 | 0.80 |
| UENF 1787 | 0.61 | 2 | 0.48 | 0.11 | 0.79 |
| UENF 1788 | 0.56 | 2 | 0.49 | 0 | 1.00 |
| UENF 1791 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2036 | 0.94 | 2 | 0.10 | 0.11 | 0 |
| UENF 2044 | 0.78 | 2 | 0.35 | 0 | 1.00 |
| UENF 2051 | 0.89 | 2 | 0.20 | 0 | 1.00 |
| UENF 2066 | 0.81 | 2 | 0.30 | 0.13 | 0.63 |
| UENF 2073 | 0.61 | 2 | 0.48 | 0.11 | 0.79 |
| UENF 2074 | 0.78 | 2 | 0.35 | 0 | 1.00 |
| UENF 2075 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2078 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2079 | 0.67 | 2 | 0.44 | 0.22 | 0.54 |
| UENF 2082 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 2083 | 0.56 | 2 | 0.49 | 0 | 1.00 |
| UENF 2085 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2086 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 2087 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 2091 | 0.89 | 2 | 0.20 | 0 | 1.00 |
| UENF 2093 | 0.56 | 2 | 0.49 | 0.13 | 0.77 |
| UENF 2097 | 0.71 | 2 | 0.41 | 0 | 1.00 |
| UENF 2100 | 0.78 | 2 | 0.35 | 0.22 | 0.41 |
| UENF 2101 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 2106 | 0.83 | 2 | 0.28 | 0.33 | −0.14 |
| UENF 2107 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 2110 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2111 | 0.56 | 2 | 0.49 | 0 | 1.00 |
| UENF 2112 | 0.81 | 2 | 0.30 | 0.13 | 0.63 |
| UENF 2113 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 2115 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2116 | 0.61 | 2 | 0.48 | 0.11 | 0.79 |
| UENF 2118 | 0.94 | 2 | 0.10 | 0.11 | 0 |
| UENF 2123 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 2129 | 0.83 | 2 | 0.28 | 0.11 | 0.64 |
| UENF 2130 | 0.61 | 2 | 0.48 | 0.11 | 0.79 |
| UENF 2131 | 0.72 | 2 | 0.40 | 0.11 | 0.75 |
| UENF 2133 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 2134 | 0.56 | 2 | 0.49 | 0 | 1.00 |
| UENF 2135 | 0.67 | 2 | 0.44 | 0.22 | 0.54 |
| UENF 2137 | 0.67 | 2 | 0.44 | 0 | 1.00 |
| UENF 2140 | 0.78 | 2 | 0.35 | 0.22 | 0.41 |
| UENF 2153 | 0.63 | 2 | 0.47 | 0 | 1.00 |
| UENF 2154 | 0.61 | 2 | 0.48 | 0.11 | 0.79 |
| Mean | 0.72 | 2 | 0.38 | 0.10 | 0.77 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvares Bianchi, P.; Renata Almeida da Silva, L.; André da Silva Alencar, A.; Henrique Araújo Diniz Santos, P.; Pimenta, S.; Pombo Sudré, C.; Erpen-Dalla Corte, L.; Simões Azeredo Gonçalves, L.; Rodrigues, R. Biomorphological Characterization of Brazilian Capsicum Chinense Jacq. Germplasm. Agronomy 2020, 10, 447. https://doi.org/10.3390/agronomy10030447
Alvares Bianchi P, Renata Almeida da Silva L, André da Silva Alencar A, Henrique Araújo Diniz Santos P, Pimenta S, Pombo Sudré C, Erpen-Dalla Corte L, Simões Azeredo Gonçalves L, Rodrigues R. Biomorphological Characterization of Brazilian Capsicum Chinense Jacq. Germplasm. Agronomy. 2020; 10(3):447. https://doi.org/10.3390/agronomy10030447
Chicago/Turabian StyleAlvares Bianchi, Paola, Lígia Renata Almeida da Silva, Antônio André da Silva Alencar, Pedro Henrique Araújo Diniz Santos, Samy Pimenta, Cláudia Pombo Sudré, Lígia Erpen-Dalla Corte, Leandro Simões Azeredo Gonçalves, and Rosana Rodrigues. 2020. "Biomorphological Characterization of Brazilian Capsicum Chinense Jacq. Germplasm" Agronomy 10, no. 3: 447. https://doi.org/10.3390/agronomy10030447