Genetic Basis Dissection for Eating and Cooking Qualities of Japonica Rice in Northeast China

Abstract
:1. Introduction
2. Methods
2.1. Plant Materials
2.2. AC, GC, and GT Evaluation in Rice Grain
2.3. Genotyping, Kinship, and LD Decay Analysis
2.4. Genome-Wide Associated Mapping
2.5. Candidate Gene Analysis
3. Results
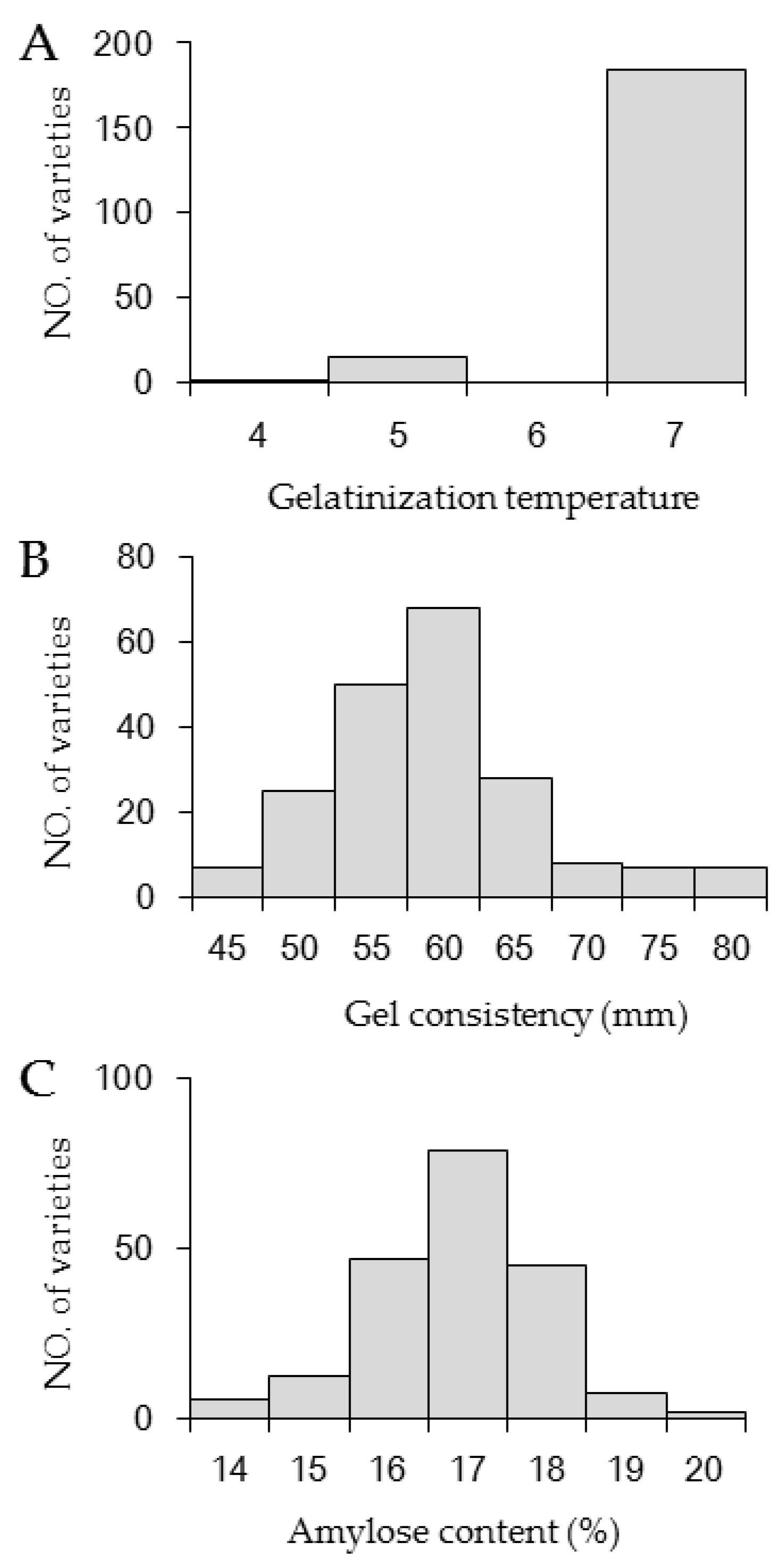
3.1. Distributions of AC, GC, and GT in the Japonica Population
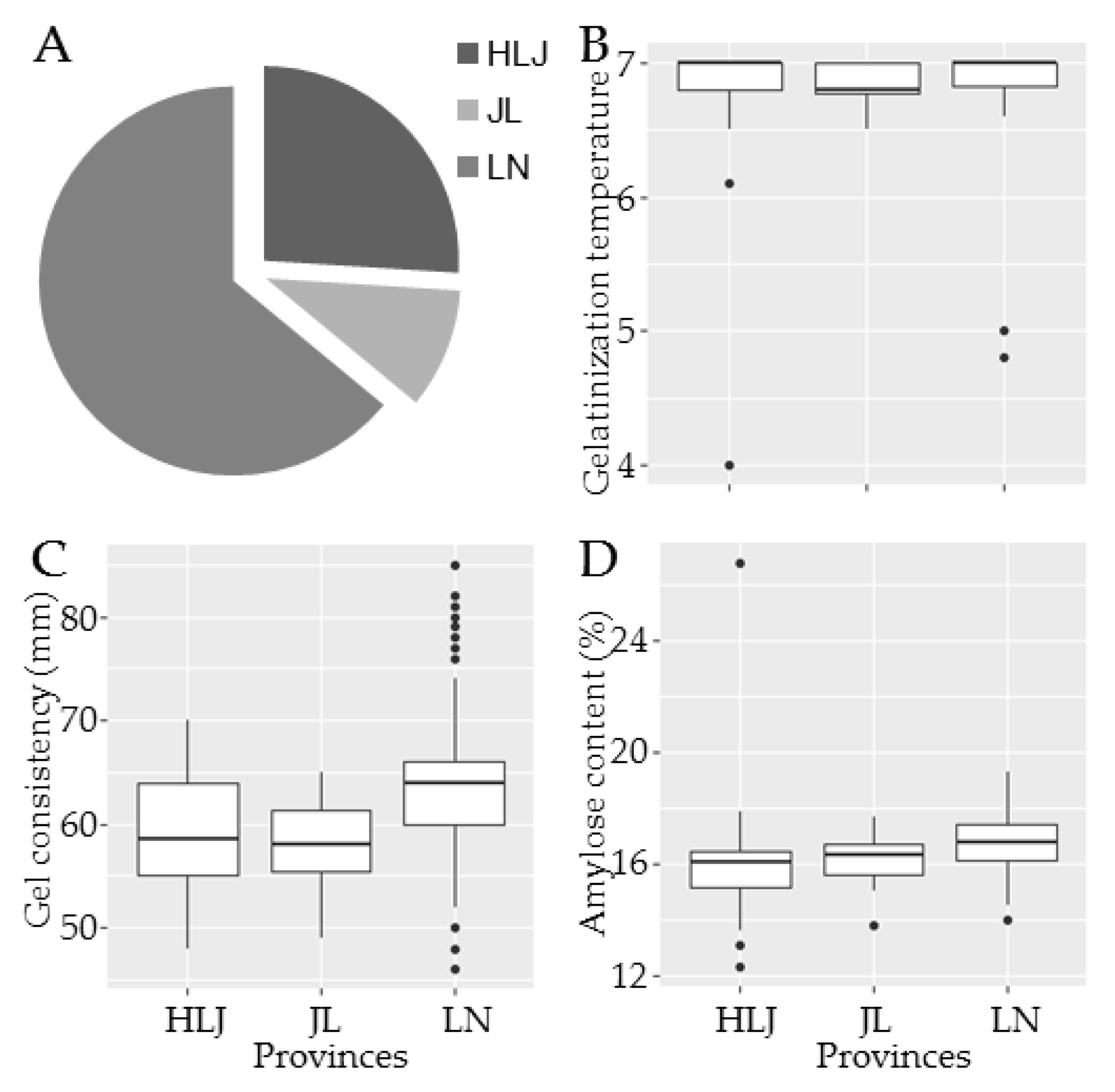
3.2. Geographical Distributions of Rice Varieties
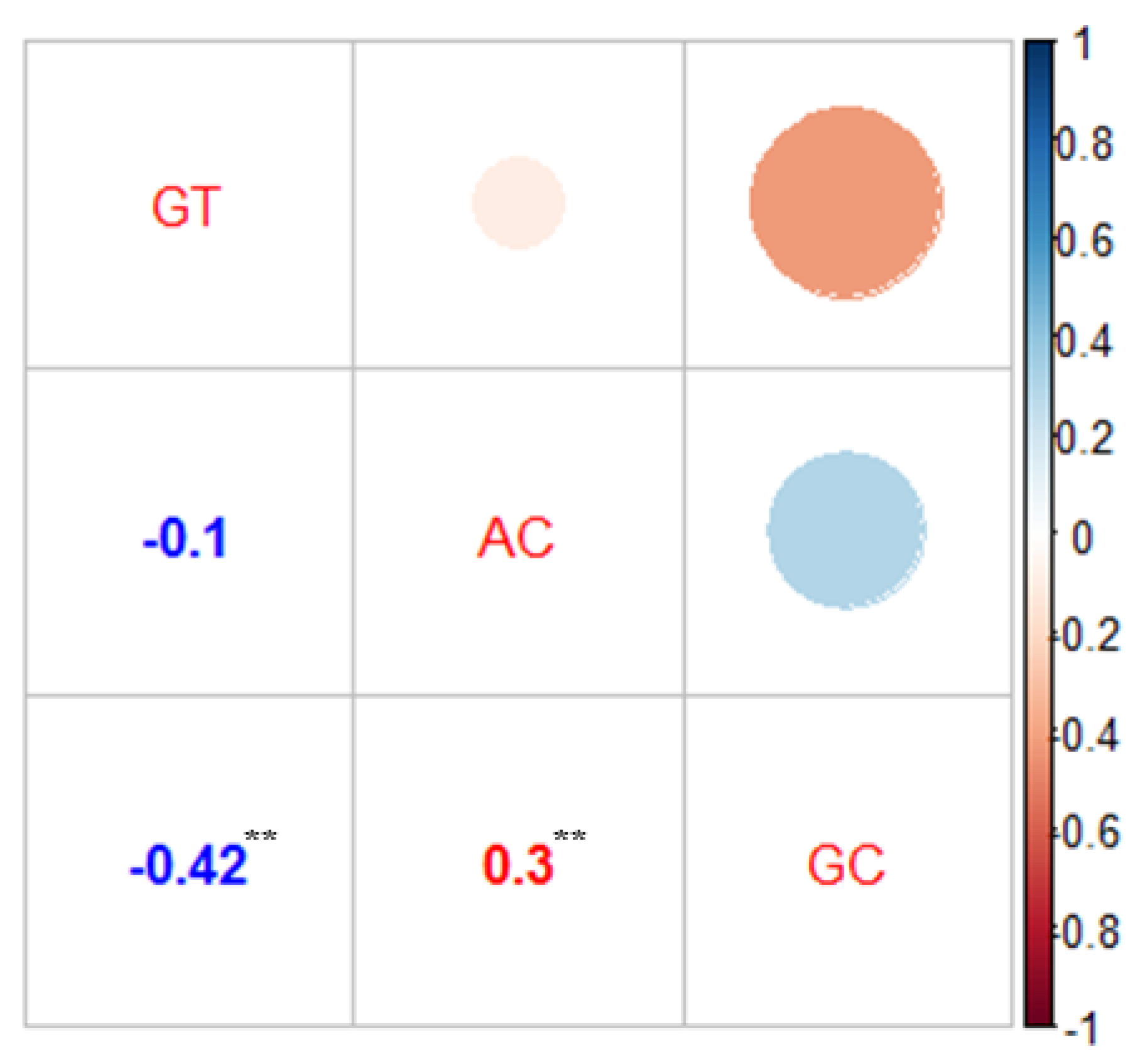
3.3. Correlation Analysis of AC, GC, and GT
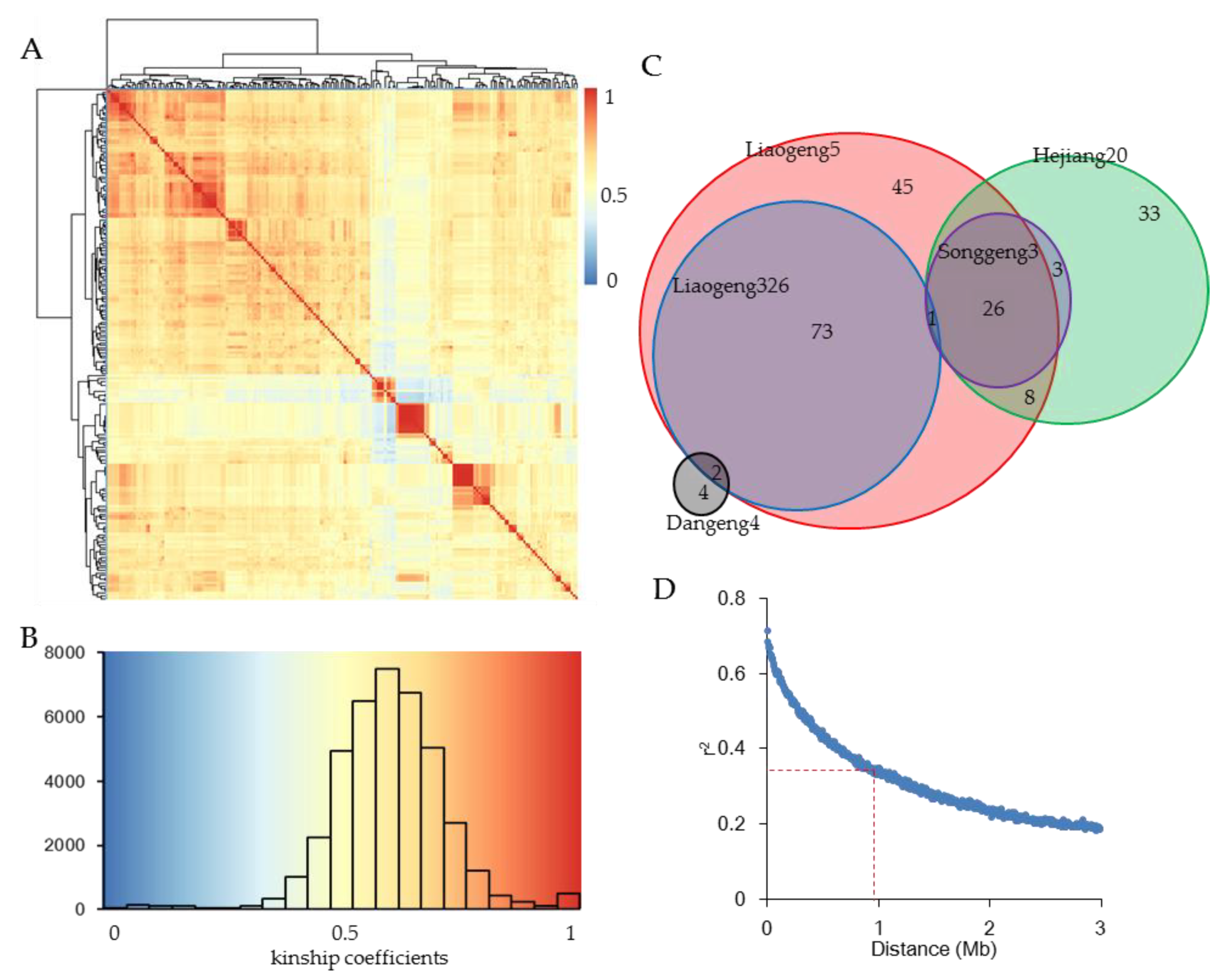
3.4. The Japonica Varieties in Northeast China Displayed Narrow Genetic Basis
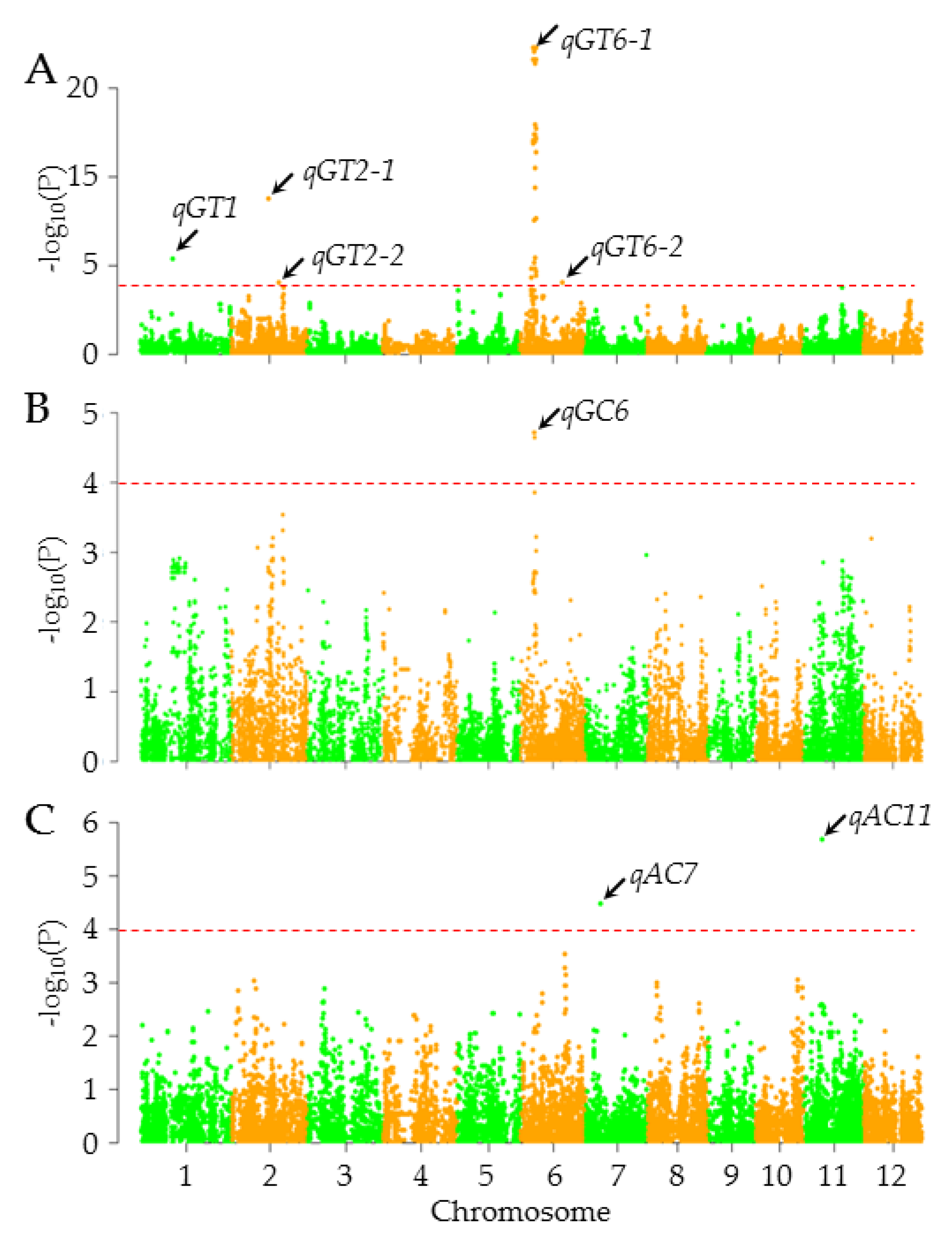
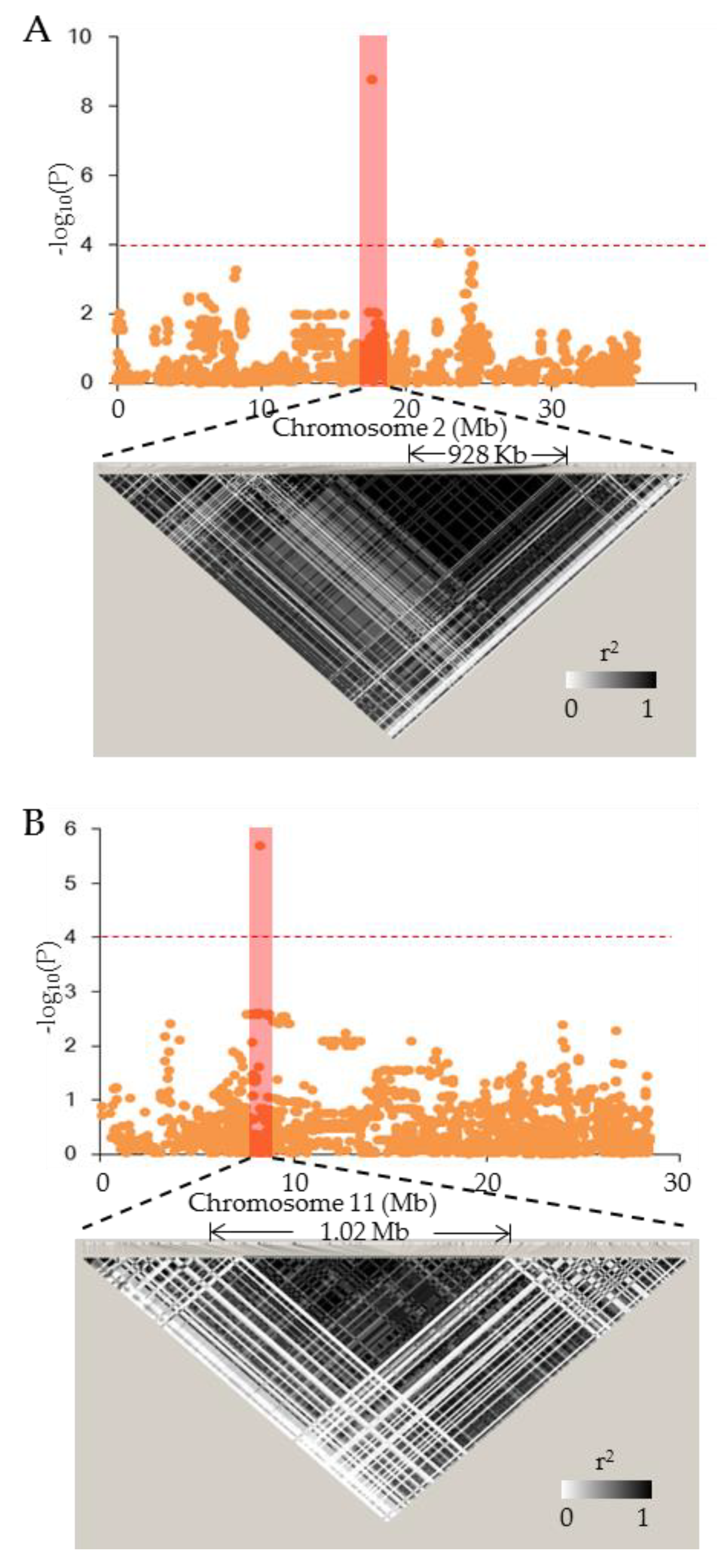
3.5. GWAS for AC, GC, and GT
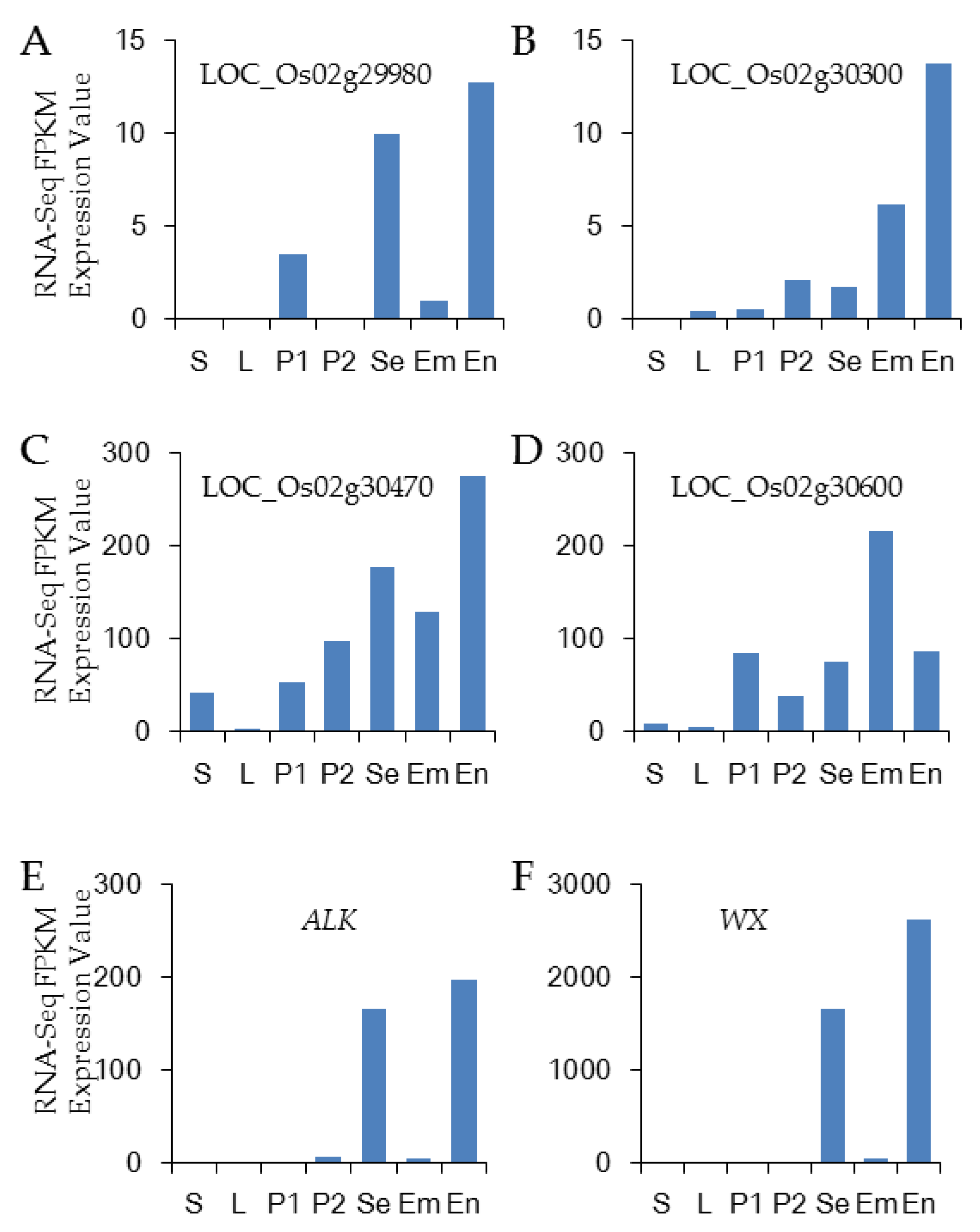
3.6. Candidate Gene Analysis
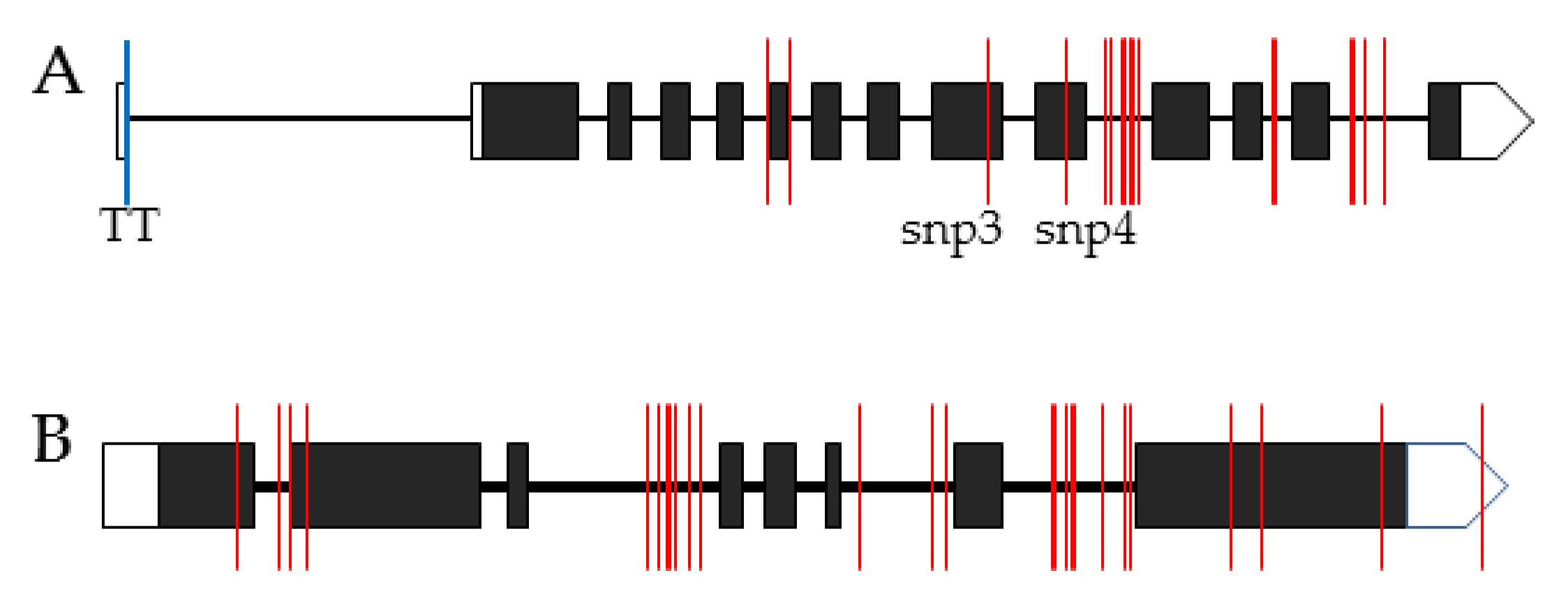
3.7. Analysis of ALK and Wx in the Japonica Varieties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Delseny, M.; Salses, J.; Cooke, R.; Sallaud, C.; Regad, F.; Lagoda, P.; Guiderdoni, E.; Ventelon, M.; Brugidou, C.; Ghesquiere, A. Rice genomics: Present and future. Plant Physiol. Bioch. 2001, 39, 323–334. [Google Scholar] [CrossRef]
- Preiss, J. Biology and molecular biology of starch synthesis and its regulation. Plant Mol.Cell. biol. 1991, 7, 5880–5883. [Google Scholar]
- Bao, J.S.; Shen, S.Q.; Sun, M.; Corke, H. Analysis of genotypic diversity in the starch physicochemical properties of nonwaxy rice: Apparent amylose content, pasting viscosity and gel texture. Starch-Starke 2006, 58, 259–267. [Google Scholar] [CrossRef]
- Hu, P.S.; Zhao, H.J.; Duan, Z.Y.; Zhang, L.L.; Wu, D.X. Starch digestibility and the estimated glycemic score of different types of rice differing in amylose contents. J. Cereal Sci. 2004, 40, 231–237. [Google Scholar] [CrossRef]
- Tang, S.X.; Khush, G.S.; Juliano, B.O. Genetics of gel consistency in rice ( Oryza sativa L.). J. Genet. 1991, 70, 69–78. [Google Scholar] [CrossRef]
- Khush, G.S.; Paule, C.M.; De, L.; Cruz, N.M. Rice grain quality evaluation and improvement at IRRI. In the Workshop on Chemical Aspects of Rice Grain Quality; IRRI (ed. Brady NC), the Philippines: Laguna, Los Banos, 1979; pp. 20–31. [Google Scholar]
- Wang, Z.Y.; Zheng, F.Q.; Shen, G.Z.; Gao, J.P.; Snustad, D.P.; Li, M.G.; Zhang, J.L.; Hong, M.M. The Amylose Content In Rice Endosperm Is Related To the Posttranscriptional Regulation Of the Waxy Gene. Plant J. 1995, 7, 613–622. [Google Scholar] [CrossRef]
- Fan, C.C.; Yu, X.Q.; Xing, Y.Z.; Xu, C.G.; Luo, L.J.; Zhang, Q.F. The main effects, epistatic effects and environmental interactions of QTLs on the cooking and eating quality of rice in a doubled-Haploid line population. Theor. Appl. Genet. 2005, 110, 1445–1452. [Google Scholar] [CrossRef]
- Su, Y.; Rao, Y.C.; Hu, S.K.; Yang, Y.L.; Gao, Z.Y.; Zhang, G.H.; Liu, J.; Hu, J.; Yan, M.X.; Dong, G.J.; et al. Map-Based cloning proves qGC-6, a major QTL for gel consistency of japonica/indica cross, responds by Waxy in rice (Oryza sativa L.). Theor. Appl. Genet. 2011, 123, 859–867. [Google Scholar] [CrossRef]
- Tan, Y.F.; Li, J.X.; Yu, S.B.; Xing, Y.Z.; Xu, C.G.; Zhang, Q. The three important traits for cooking and eating quality of rice grains are controlled by a single locus in an elite rice hybrid, Shanyou 63. Theor. Appl. Genet. 1999, 99, 642–648. [Google Scholar] [CrossRef]
- Isshiki, M.; Morino, K.; Nakajima, M.; Okagaki, R.J.; Wessler, S.R.; Izawa, T.; Shimamoto, K. A naturally occurring functional allele of the rice waxy locus has a GT to TT mutation at the 5′ splice site of the first intron. Plant J. 1998, 15, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.Y.; Zeng, D.L.; Cui, X.; Zhou, Y.H.; Yan, M.X.; Huang, D.N.; Li, J.Y.; Qian, Q. Map-Based cloning of the ALK gene, which controls the gelatinization temperature of rice. Sci. China Ser. C 2003, 46, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.X.; Qian, Q.; Liu, Q.Q.; Yan, M.X.; Liu, X.F.; Yan, C.J.; Liu, G.F.; Gao, Z.Y.; Tang, S.Z.; Zeng, D.L.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Han, Y.P.; Jiang, L.; Xu, C.W.; Lu, J.F.; Xu, M.L. Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities. Mol. Breed. 2006, 18, 277–290. [Google Scholar] [CrossRef]
- Sun, S.Y.; Wei, H.; Lin, H.X. Identification of QTLs for Cooking and Eating Quality of Rice Grain. Rice Sci. 2006, 13, 161–169. [Google Scholar]
- Hsu, Y.C.; Tseng, M.C.; Wu, Y.P.; Lin, M.Y.; Wei, F.J.; Hwu, K.K.; Hsing, Y.I.; Lin, Y.R. Genetic factors responsible for eating and cooking qualities of rice grains in a recombinant inbred population of an inter-subspecific cross. Mol. Breed. 2014, 34, 655–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.M.; Mu, P.; Liu, J.F.; Lu, Y.X.; Li, Z.C. QTL mapping and QxE interactions of grain cooking and nutrient qualities in rice under upland and lowland environments. J. Genet. Genom. 2007, 34, 420–428. [Google Scholar] [CrossRef]
- Leng, Y.J.; Xue, D.W.; Yang, Y.L.; Hu, S.K.; Su, Y.; Huang, L.C.; Wang, L.; Zheng, T.T.; Zhang, G.H.; Hu, J.; et al. Mapping of QTLs for eating and cooking quality-Related traits in rice (Oryza sativa L.). Euphytica 2014, 197, 99–108. [Google Scholar] [CrossRef]
- Huang, X.H.; Wei, X.H.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.Y.; Zhu, C.R.; Lu, T.T.; Zhang, Z.W.; et al. Genome-Wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef]
- Wang, C.H.; Yang, Y.L.; Yuan, X.P.; Xu, Q.; Feng, Y.; Yu, H.Y.; Wang, Y.P.; Wei, X.H. Genome-Wide association study of blast resistance in indica rice. BMC Plant Biol. 2014, 14, 311. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.F.; Bao, J.S.; He, Q.; Park, Y.J. Genome-Wide association study of eating and cooking qualities in different subpopulations of rice (Oryza sativa L.). BMC Genom. 2016, 17, 663. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Pang, Y.L.; Zhang, J.; Wu, Z.C.; Chen, K.; Ali, J.; Ye, G.Y.; Xu, J.L.; Li, Z.K. Genome-Wide and gene-Based association mapping for rice eating and cooking characteristics and protein content. Sci. Rep. 2017, 7, 17203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.M.; Zhong, L.J. Variation of rice quality traits under different climate conditions and its main affected factors. Chin. J. Rice Sci. 2001, 15, 187–191. [Google Scholar]
- Ye, J.; Niu, X.J.; Yang, Y.L.; Wang, S.; Xu, Q.; Yuan, X.P.; Yu, H.Y.; Wang, Y.P.; Wang, S.; Feng, Y.; et al. Divergent Hd1, Ghd7, and DTH7 Alleles Control Heading Date and Yield Potential of Japonica Rice in Northeast China. Front. Plant Sci. 2018, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C. Haploview: Visualization and analysis of SNP genotype data. Cold Spring Harb. Protoc. 2009, 2009. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Arora, R.; Agarwal, P.; Ray, S.; Sharma, P.; Kapoor, S.; Tyagi, A.K.; Khurana, J.P. F-Box Proteins in Rice. Genome-Wide Analysis, Classification, Temporal and Spatial Gene Expression during Panicle and Seed Development, and Regulation by Light and Abiotic Stress. Plant Physiol. 2007, 143, 1467–1483. [Google Scholar] [CrossRef] [Green Version]
- Hirano, H.Y.; Eiguchi, M.; Sano, Y. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol. 1998, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhou, G.S.; Liu, D.L.; Wang, B.; Xiao, D.P.; He, L. Climate-Associated rice yield change in the Northeast China Plain: A simulation analysis based on CMIP5 multi-Model ensemble projection. Sci. Total Environ. 2019, 666, 126–138. [Google Scholar] [CrossRef]
- Biselli, C.; Cavalluzzo, D.; Perrini, R.; Gianinetti, A.; Bagnaresi, P.; Urso, S.; Orasen, G.; Desiderio, F.; Lupotto, E.; Cattivelli, L.; et al. Improvement of marker-Based predictability of Apparent Amylose Content in japonica rice through GBSSI allele mining. Rice 2014, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.H.; Yang, S.H.; Gong, J.Y.; Zhao, Q.; Feng, Q.; Zhan, Q.L.; Zhao, Y.; Li, W.J.; Cheng, B.Y.; Xia, J.H.; et al. Genomic architecture of heterosis for yield traits in rice. Nature 2016, 537, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhang, M.C.; Niu, X.J.; Wang, S.; Xu, Q.; Feng, Y.; Wang, C.H.; Deng, H.Z.; Yuan, X.P.; Yu, H.Y.; et al. Genetic variation and association mapping for 12 agronomic traits in indica rice. BMC Genom. 2015, 16, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, P.G.; Xu, J.S.; Zeng, D.L.; Zhang, B.L.; Geng, M.F.; Zhang, G.Z.; Huang, K.; Huang, L.J.; Xu, R.; Ge, S.; et al. Natural Variation in the Promoter of GSE5 Contributes to Grain Size Diversity in Rice. Mol. Plant 2017, 10, 685–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, L.Z.; Chen, J.Y.; Huang, X.H.; Gong, H.; Luo, J.H.; Hou, Q.Q.; Zhou, T.Y.; Lu, T.T.; Zhu, J.J.; Shangguan, Y.Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.C.; Hu, L.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K.; et al. Genome-Wide association study using whole-Genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Name | Peak SNPs | Chr | Position | P-Value | −log10P | R2 | Reported gene/QTL |
|---|---|---|---|---|---|---|---|---|
| GT | qGT1 | seq1_15261369 | 1 | 15261369 | 4.13E-06 | 5.38 | 0.14 | |
| qGT2-1 | seq2_17671386 | 2 | 17671386 | 1.68E-09 | 8.78 | −0.19 | ||
| qGT2-2 | seq2_22314159 | 2 | 22314159 | 9.33E-05 | 4.03 | 0.10 | ||
| qGT6-1 | seq6_6726252 | 6 | 6726252 | 5.25E-18 | 17.28 | 0.30 | ALK | |
| qGT6-2 | seq6_20151786 | 6 | 20151786 | 8.94E-05 | 4.05 | 0.09 | ||
| GC | qGC6 | seq6_6720486 | 6 | 6720486 | 1.90E-05 | 4.72 | −0.21 | ALK |
| AC | qAC7 | seq7_6847358 | 7 | 6847358 | 3.36E-05 | 4.47 | −0.20 | |
| qAC11 | seq11_8300677 | 11 | 8300677 | 2.04E-06 | 5.69 | 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xu, X.; Zhang, M.; Xu, Q.; Feng, Y.; Yuan, X.; Yu, H.; Wang, Y.; Wei, X. Genetic Basis Dissection for Eating and Cooking Qualities of Japonica Rice in Northeast China. Agronomy 2020, 10, 423. https://doi.org/10.3390/agronomy10030423
Yang Y, Xu X, Zhang M, Xu Q, Feng Y, Yuan X, Yu H, Wang Y, Wei X. Genetic Basis Dissection for Eating and Cooking Qualities of Japonica Rice in Northeast China. Agronomy. 2020; 10(3):423. https://doi.org/10.3390/agronomy10030423
Chicago/Turabian StyleYang, Yaolong, Xin Xu, Mengchen Zhang, Qun Xu, Yue Feng, Xiaoping Yuan, Hanyong Yu, Yiping Wang, and Xinghua Wei. 2020. "Genetic Basis Dissection for Eating and Cooking Qualities of Japonica Rice in Northeast China" Agronomy 10, no. 3: 423. https://doi.org/10.3390/agronomy10030423