Nitric Oxide and a Conditioned Medium Affect the Hematopoietic Development in a Microfluidic Mouse Embryonic Stem Cell/OP9 Co-Cultivation System

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Device Fabrication
2.2. Preparation of mESCs
2.3. Microfluidic Cell Culture and Differentiation
2.4. Cell Viability Assay
2.5. Immunostaining
2.6. Treatment of Cells with Activators and Inhibitors of the NO Pathway
2.7. Treatment of Cells with Conditioned Medium
2.8. Microscopy
2.9. Data Analysis
3. Results
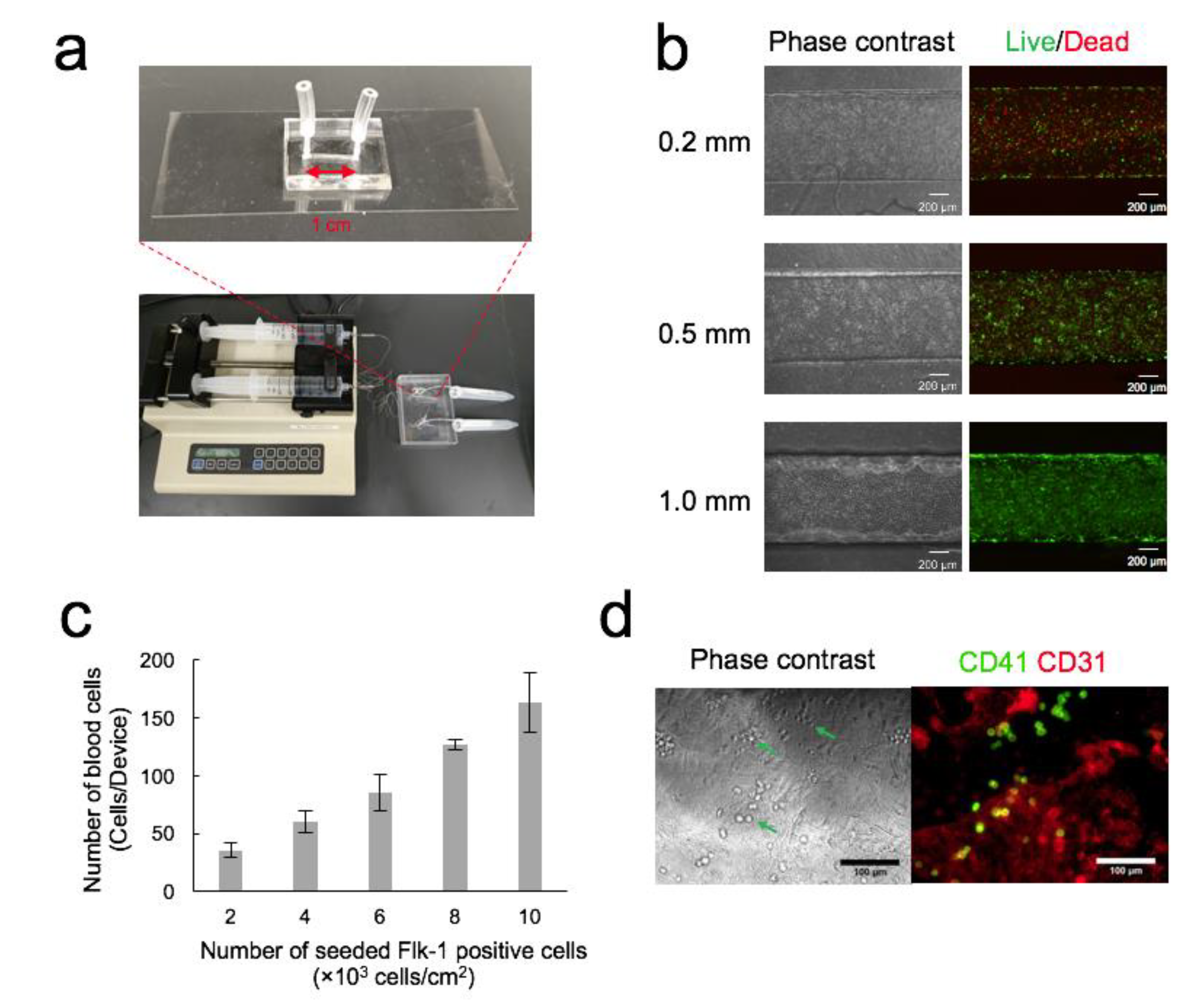
3.1. Optimization of OP9 Cell Seeding Density in Channel
3.2. Optimization of Seeding Density of Flk-1+ Cells
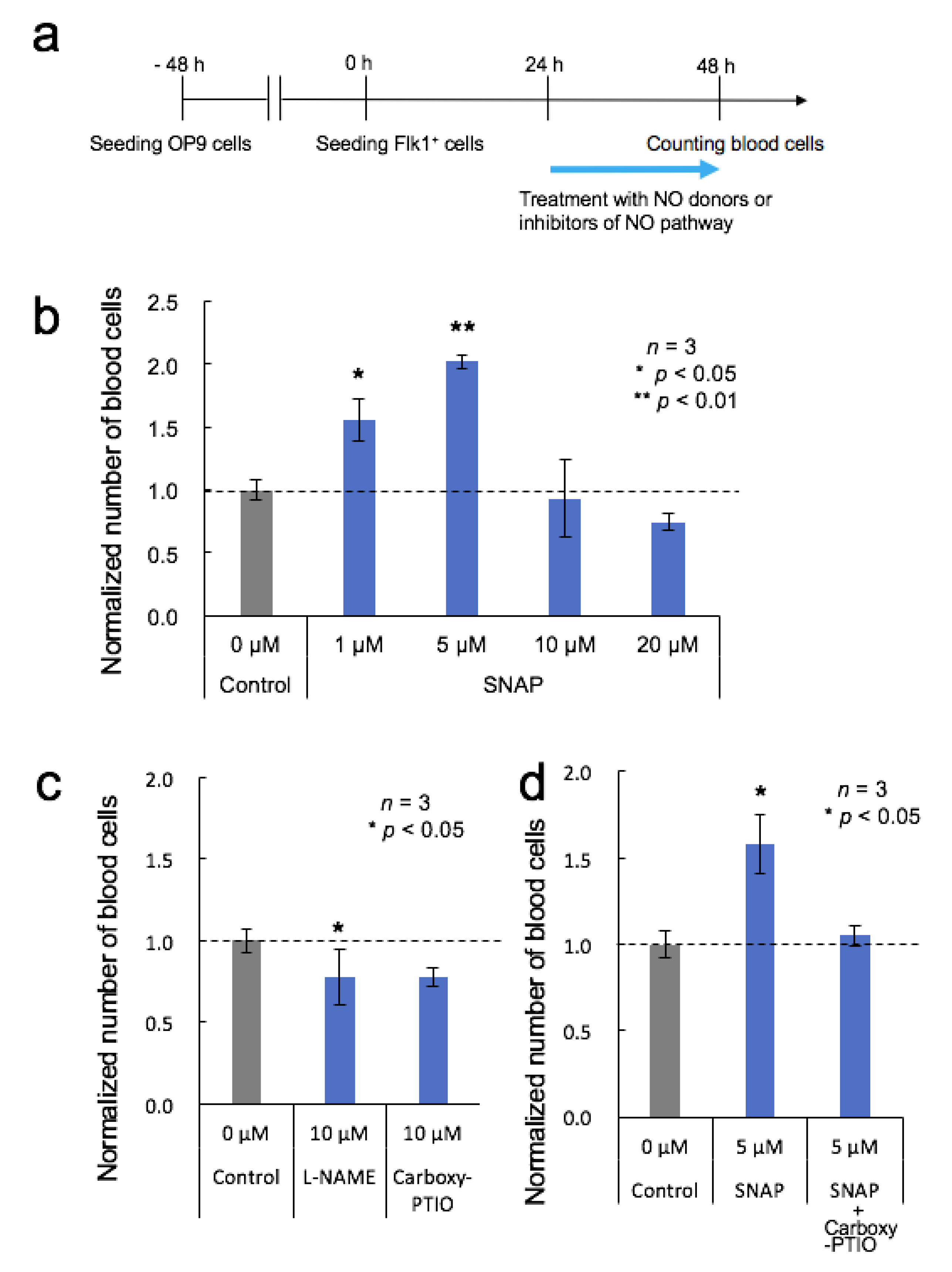
3.3. Effects of NO Donor and NO Pathway Inhibitors
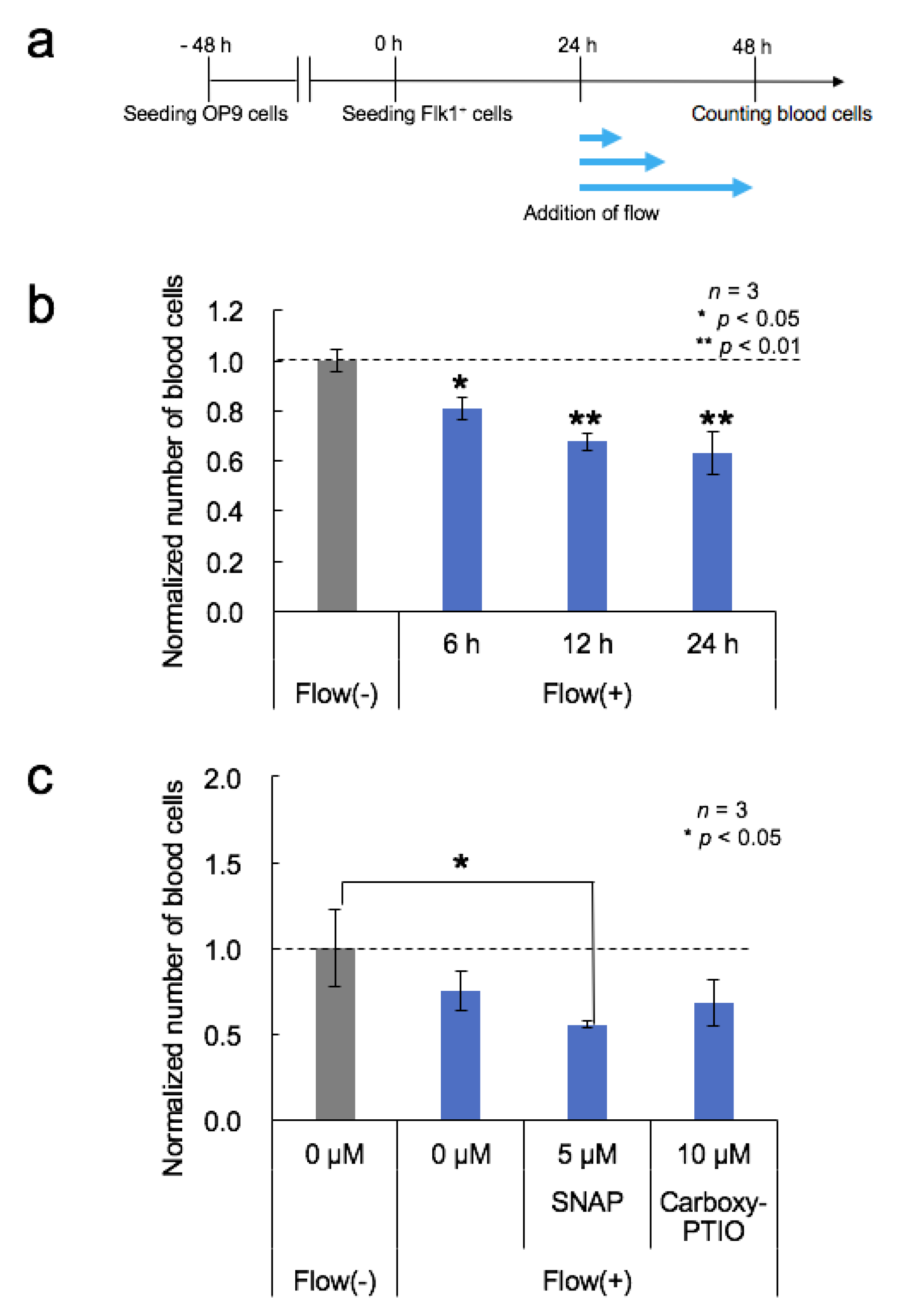
3.4. Effects of Medium Flow
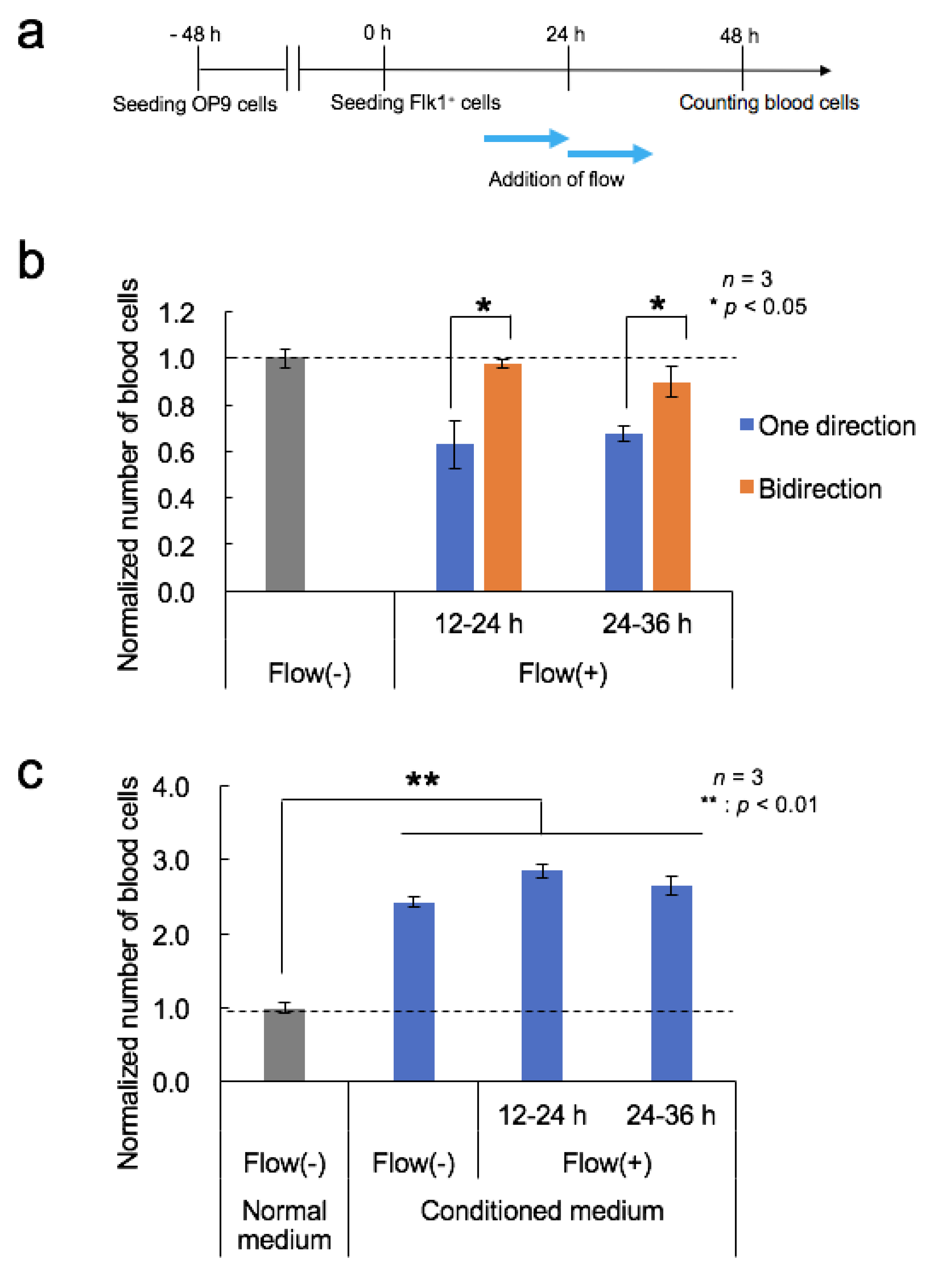
3.5. Effects of Fluid Flow Stress and Conditioned Medium
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Medvinsky, A.; Dzierzak, E. Definitive hematopoiesis is autonomously initiated by the AGM region. Cell 1996, 86, 897–906. [Google Scholar] [CrossRef] [Green Version]
- North, T.E.; Goessling, W.; Peeters, M.; Li, P.; Ceol, C.; Lord, A.M.; Weber, G.J.; Harris, J.; Cutting, C.C.; Huang, P.; et al. Hematopoietic stem cell development is dependent on blood flow. Cell 2009, 137, 736–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, L.; Naveiras, O.; Wenzel, P.L.; McKinney-Freeman, S.; Mack, P.J.; Gracia-Sancho, J.; Suchy-Dicey, A.; Yoshimoto, M.; Lensch, M.W.; Yoder, M.C.; et al. Biomechanical forces promote embryonic haematopoiesis. Nature 2009, 459, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, P.; Wei, Y.; Gao, Y.; Patient, R.; Liu, F. A blood flow-dependent klf2a-NO signaling cascade is required for stabilization of hematopoietic stem cell programming in zebrafish embryos. Blood 2011, 118, 4102–4110. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.P.; Ahsan, T. Shear stress during early embryonic stem cell differentiation promotes hematopoietic and endothelial phenotypes. Biotechnol. Bioeng. 2013, 110, 1231–1242. [Google Scholar] [CrossRef] [Green Version]
- Diaz, M.F.; Li, N.; Lee, H.J.; Adamo, L.; Evans, S.M.; Willey, H.E.; Arora, N.; Torisawa, Y.S.; Vickers, D.A.; Morris, S.A.; et al. Biomechanical forces promote blood development through prostaglandin E2 and the cAMP-PKA signaling axis. J. Exp. Med. 2015, 212, 665–680. [Google Scholar] [CrossRef]
- Kim, P.G.; Nakano, H.; Das, P.P.; Chen, M.J.; Rowe, R.G.; Chou, S.S.; Ross, S.J.; Sakamoto, K.M.; Zon, L.I.; Schlaeger, T.M.; et al. Flow-induced protein kinase A-CREB pathway acts via BMP signaling to promote HSC emergence. J. Exp. Med. 2015, 212, 633–648. [Google Scholar] [CrossRef]
- Lim, W.F.; Inoue-Yokoo, T.; Tan, K.S.; Lai, M.I.; Sugiyama, D. Hematopoietic cell differentiation from embryonic and induced pluripotent stem cells. Stem Cell Res. Ther. 2013, 4, 71. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, K.; Kanokoda, M.; Nakajima, M.; Hara, T. Domain-specific biological functions of the transcription factor Gata2 on hematopoietic differentiation of mouse embryonic stem cells. Genes Cells 2018, 23, 753–766. [Google Scholar] [CrossRef]
- Nakano, T.; Kodama, H.; Honjo, T. Generation of lymphohematopoietic cells from embryonic stem cells in culture. Science 1994, 265, 1098–1101. [Google Scholar] [CrossRef]
- Nakano, T.; Kodama, H.; Honjo, T. In vitro development of primitive and definitive erythrocytes from different precursors. Science 1996, 272, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Nakano, T. Development of hematopoietic cells from embryonic stem cells. Int. J. Hematol. 2001, 73, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kikukawa, Y.; Sato, K.; Sugii, Y.; Kitamori, T. Culture and leukocyte adhesion assay of human arterial endothelial cells in a glass microchip. Anal. Sci. 2007, 23, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, E.W.; Beebe, D.J. Fundamentals of microfluidic cell culture in controlled microenvironments. Chem. Soc. Rev. 2010, 39, 1036–1048. [Google Scholar] [CrossRef]
- Sasaki, N.; Shinjo, M.; Hirakawa, S.; Nishinaka, M.; Tanaka, Y.; Mawatari, K.; Kitamori, T.; Sato, K. A palmtop-sized microfluidic cell culture system driven by a miniaturized infusion pump. Electrophoresis 2012, 33, 1729–1735. [Google Scholar] [CrossRef]
- Sato, K.; Sasaki, N.; Svahn, H.A.; Sato, K. Microfluidics for nano-pathophysiology. Adv. Drug Deliv. Rev. 2014, 74, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wei, X.; Zeng, R.; Xu, F.; Li, X. Stem cell culture and differentiation in microfluidic devices toward organ-on-a-chip. Future Sci. OA 2017, 3, FSO187. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Sakai, Y.; Fujii, T. Organ/body-on-a-chip based on microfluidic technology for drug discovery. Drug Metab. Pharmacokinet. 2018, 33, 43–48. [Google Scholar] [CrossRef]
- Rogal, J.; Zbinden, A.; Schenke-Layland, K.; Loskill, P. Stem-cell based organ-on-a-chip models for diabetes research. Adv. Drug Deliv. Rev. 2018. [Google Scholar] [CrossRef]
- Ergir, E.; Bachmann, B.; Redl, H.; Forte, G.; Ertl, P. Small Force, Big Impact: Next Generation Organ-on-a-Chip Systems Incorporating Biomechanical Cues. Front. Physiol. 2018, 9, 1417. [Google Scholar] [CrossRef]
- Lee, S.; Ko, J.; Park, D.; Lee, S.R.; Chung, M.; Lee, Y.; Jeon, N.L. Microfluidic-based vascularized microphysiological systems. Lab Chip 2018, 18, 2686–2709. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Sato, K. Recent Progress in the Development of Microfluidic Vascular Models. Anal. Sci. 2018, 34, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Sato, M.; Yokoyama, M.; Hirai, M.; Furuta, A. Influence of Culture Conditions on Cell Proliferation in a Microfluidic Channel. Anal. Sci. 2019, 35, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Nitta, M.; Ogawa, A. A Microfluidic Cell Stretch Device to Investigate the Effects of Stretching Stress on Artery Smooth Muscle Cell Proliferation in Pulmonary Arterial Hypertension. Inventions 2019, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Ishigaki, Y.; Sato, K. Effects of Microchannel Shape and Ultrasonic Mixing on Microfluidic Padlock Probe Rolling Circle Amplification (RCA) Reactions. Micromachines 2018, 9, 272. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Nakajima, M.; Tokuda, S.; Ogawa, A. Fluidic Culture and Analysis of Pulmonary Artery Smooth Muscle Cells for the Study of Pulmonary Hypertension. Anal. Sci. 2016, 32, 1217–1221. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, A.; Ishigaki, Y.; Nilsson, M.; Sato, K.; Sato, K. Microfluidics-based in situ padlock/rolling circle amplification system for counting single DNA molecules in a cell. Anal. Sci. 2014, 30, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Sasaki, N.; Ato, M.; Hirakawa, S.; Sato, K.; Sato, K. Microcirculation-on-a-Chip: A Microfluidic Platform for Assaying Blood- and Lymphatic-Vessel Permeability. PLoS ONE 2015, 10, e0137301. [Google Scholar] [CrossRef]
- Imura, Y.; Asano, Y.; Sato, K.; Yoshimura, E. A microfluidic system to evaluate intestinal absorption. Anal. Sci. 2009, 25, 1403–1407. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.A.; Baron, M.H.; Fraser, S.E.; Dickinson, M.E. Measuring hemodynamic changes during mammalian development. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1561–H1569. [Google Scholar] [CrossRef]
- Weninger, W.J.; Maurer, B.; Zendron, B.; Dorfmeister, K.; Geyer, S.H. Measurements of the diameters of the great arteries and semi-lunar valves of chick and mouse embryos. J. Microsc. 2009, 234, 173–190. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, K.; Maeda, M.; Kamata, E.; Ishii, S.; Yanagisawa, K.; Kitajima, K.; Hara, T. Nitric Oxide and a Conditioned Medium Affect the Hematopoietic Development in a Microfluidic Mouse Embryonic Stem Cell/OP9 Co-Cultivation System. Micromachines 2020, 11, 305. https://doi.org/10.3390/mi11030305
Sato K, Maeda M, Kamata E, Ishii S, Yanagisawa K, Kitajima K, Hara T. Nitric Oxide and a Conditioned Medium Affect the Hematopoietic Development in a Microfluidic Mouse Embryonic Stem Cell/OP9 Co-Cultivation System. Micromachines. 2020; 11(3):305. https://doi.org/10.3390/mi11030305
Chicago/Turabian StyleSato, Kae, Momoko Maeda, Eriko Kamata, Sayaka Ishii, Kanako Yanagisawa, Kenji Kitajima, and Takahiko Hara. 2020. "Nitric Oxide and a Conditioned Medium Affect the Hematopoietic Development in a Microfluidic Mouse Embryonic Stem Cell/OP9 Co-Cultivation System" Micromachines 11, no. 3: 305. https://doi.org/10.3390/mi11030305