Equivocal Evidence for Colony Level Stress Effects on Bumble Bee Pollination Services

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set up
2.1.1. Insecticide Stress
2.1.2. Field Cages
2.1.3. Heat Stress
2.1.4. Crop Pollination and Foraging Behaviour
2.1.5. Foraging Behaviour
2.2. Assessing Pollination Effects on Seed Set
2.3. Statistical Analysis
2.3.1. Behaviour
2.3.2. Plant Yields
3. Results
3.1. Behaviour Responses to Heat Stress and Pesticide
3.2. Yield parameters
4. Discussion
4.1. Heat Stress
4.2. Clothianidin Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A meta-analysis of crop yield under climate change and adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Bishop, J.; Jones, H.E.; Lukac, M.; Potts, S.G. Insect pollination reduces yield loss following heat stress in faba bean (Vicia faba L.). Agric. Ecosyst. Environ. 2016, 220, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Bishop, J.; Jones, H.E.; O’Sullivan, D.M.; Potts, S.G. Elevated temperature drives a shift from selfing to outcrossing in the insect-pollinated legume, faba bean (Vicia faba). J. Exp. Bot. 2017, 68, 2055–2063. [Google Scholar] [CrossRef] [Green Version]
- Martinet, B.; Lecocq, T.; Smet, J.; Rasmont, P. A protocol to assess insect resistance to heat waves, applied to bumblebees (Bombus Latreille, 1802). PLoS ONE 2015, 10, e0118591. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.L.; DeGabriel, J.L.; Riegler, M.; Cook, J.M. A temperate pollinator with high thermal tolerance is still susceptible to heat events predicted under future climate change. Ecol. Entomol. 2018, 43, 506–512. [Google Scholar] [CrossRef]
- Medina, R.G.; Paxton, R.J.; De Luna, E.; Fleites-Ayil, F.A.; Medina Medina, L.A.; Quezada-Euán, J.J.G. Developmental stability, age at onset of foraging and longevity of Africanized honey bees (Apis mellifera L.) under heat stress (Hymenoptera: Apidae). J. Therm. Biol. 2018, 74, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Bordier, C.; Dechatre, H.; Suchail, S.; Peruzzi, M.; Soubeyrand, S.; Pioz, M.; Pélissier, M.; Crauser, D.; Conte, Y.L.; Alaux, C. Colony adaptive response to simulated heat waves and consequences at the individual level in honeybees (Apis mellifera). Sci. Rep. 2017, 7, 3760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidenmüller, A.; Kleineidam, C.; Tautz, J. Collective control of nest climate parameters in bumblebee colonies. Anim. Behav. 2002, 63, 1065–1071. [Google Scholar] [CrossRef] [Green Version]
- Weidenmuller, A. The control of nest climate in bumblebee (Bombus terrestris) colonies: Interindividual variability and self reinforcement in fanning response. Behav. Ecol. 2004, 15, 120–128. [Google Scholar] [CrossRef]
- Arce, A.N.; David, T.I.; Randall, E.L.; Ramos Rodrigues, A.; Colgan, T.J.; Wurm, Y.; Gill, R.J. Impact of controlled neonicotinoid exposure on bumblebees in a realistic field setting. J. Appl. Ecol. 2017, 54, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Isaac, N.J.B.; Bullock, J.M.; Roy, D.B.; Garthwaite, D.G.; Crowe, A.; Pywell, R.F. Impacts of neonicotinoid use on long-term population changes in wild bees in England. Nat. Commun. 2016, 7, 12459. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Firbank, L.G.; Petit, S.; Smart, S.; Blain, A.; Fuller, R.J. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 777–787. [Google Scholar] [CrossRef]
- Krauss, J.; Bommarco, R.; Guardiola, M.; Heikkinen, R.K.; Helm, A.; Kuussaari, M.; Lindborg, R.; Ockinger, E.; Pärtel, M.; Pino, J.; et al. Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecol. Lett. 2010, 13, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D. REVIEW: An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.A.; Garratt, M.P.D.; Wickens, J.B.; Wickens, V.J.; Potts, S.G.; Raine, N.E. Neonicotinoid pesticide exposure impairs crop pollination services provided by bumblebees. Nature 2015, 528, 548–550. [Google Scholar] [CrossRef] [Green Version]
- Tosi, S.; Démares, F.J.; Nicolson, S.W.; Medrzycki, P.; Pirk, C.W.W.; Human, H. Effects of a neonicotinoid pesticide on thermoregulation of African honey bees (Apis mellifera scutellata). J. Insect Physiol. 2016, 93, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Crall, J.D.; Switzer, C.M.; Oppenheimer, R.L.; Ford Versypt, A.N.; Dey, B.; Brown, A.; Eyster, M.; Guérin, C.; Pierce, N.E.; Combes, S.A.; et al. Neonicotinoid exposure disrupts bumblebee nest behavior, social networks, and thermoregulation. Science 2018, 362, 683–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterman, J.; Wintermantel, D.; Locke, B.; Jonsson, O.; Semberg, E.; Onorati, P.; Forsgren, E.; Rosenkranz, P.; Rahbek-Pedersen, T.; Bommarco, R.; et al. Clothianidin seed-treatment has no detectable negative impact on honeybee colonies and their pathogens. Nat. Commun. 2019, 10, 692. [Google Scholar] [CrossRef] [Green Version]
- Godfray, H.C.J.; Blacquière, T.; Field, L.M.; Hails, R.S.; Petrokofsky, G.; Potts, S.G.; Raine, N.E.; Vanbergen, A.J.; McLean, A.R. A restatement of the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garratt, M.P.D.; Coston, D.J.; Truslove, C.L.; Lappage, M.G.; Polce, C.; Dean, R.; Biesmeijer, J.C.; Potts, S.G. The identity of crop pollinators helps target conservation for improved ecosystem services. Biol. Conserv. 2014, 169, 128–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, S.; Tiedeken, E.J.; Simcock, K.L.; Derveau, S.; Mitchell, J.; Softley, S.; Stout, J.C.; Wright, G.A. Bees prefer foods containing neonicotinoid pesticides. Nature 2015, 521, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.M.; Willis, S.J.; Wright, G.A. Exposure to neonicotinoids influences the motor function of adult worker honeybees. Ecotoxicology 2014, 23, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Qian, W.; Suppanz, I.; Wei, L.; Mao, B.; Long, Y.; Meng, J.; Müller, A.E.; Jung, C. Flowering time variation in oilseed rape (Brassica napus L.) is associated with allelic variation in the FRIGIDA homologue BnaA.FRI.a. J. Exp. Bot. 2011, 62, 5641–5658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, D.A.; Raine, N.E. Chronic exposure to a neonicotinoid pesticide alters the interactions between bumblebees and wild plants. Funct. Ecol. 2016, 30, 1132–1139. [Google Scholar] [CrossRef]
- Herrera, C.M. Daily Patterns of Pollinator Activity, Differential Pollinating Effectiveness, and Floral Resource Availability, in a Summer-Flowering Mediterranean Shrub. Oikos 1990, 58, 277. [Google Scholar] [CrossRef] [Green Version]
- Vogt, F.D. Thermoregulation in Bumblebee Colonies. I. Thermoregulatory versus Brood-Maintenance Behaviors during Acute Changes in Ambient Temperature. Physiol. Zool. 1986, 59, 55–59. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M. Chapter 3: Impacts of 1.5 °C Global Warming on Natural and Human Systems; Global Warming of 1.5 °C; IPCC: Geneva, Switzerland, 2018; pp. 3–246. [Google Scholar]
- Kendall, D.A.; Smith, B.D. The Pollinating Efficiency of Honeybee and Bumblebee Visits to Field Bean Flowers (Vicia faba L.). J. Appl. Ecol. 1975, 12, 709. [Google Scholar] [CrossRef]
- Fijen, T.P.M.; Scheper, J.A.; Boom, T.M.; Janssen, N.; Raemakers, I.; Kleijn, D. Insect pollination is at least as important for marketable crop yield as plant quality in a seed crop. Ecol. Lett. 2018, 21, 1704–1713. [Google Scholar] [CrossRef]
- Stout, J.C.; Kells, A.R.; Goulson, D. Pollination of the invasive exotic Lupinus arboreus (Fabaceae) by introduced bees in Tasmania. Biol. Conserv. 2002, 106, 425–434. [Google Scholar] [CrossRef]
- Stout, J.C.; Allen, J.A.; Goulson, D. Nectar robbing, forager efficiency and seed set: Bumblebees foraging on the self incompatible plant Linaria vulgaris (Scrophulariaceae). Acta Oecologica 2000, 21, 277–283. [Google Scholar] [CrossRef]
- Knott, C.M. A key for stages of development of the faba bean (Vicia faba). Ann. Appl. Biol. 1990, 116, 391–404. [Google Scholar] [CrossRef]
- Carpenter, B.; Gelman, A.; Hoffman, M.D.; Lee, D.; Goodrich, B.; Betancourt, M.; Brubaker, M.; Guo, J.; Li, P.; Riddell, A. Stan: A probabilistic programming language. J. Stat. Softw. 2017, 76, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Bürkner, P.C. Brms: An R package for bayesian multilevel models using stan. J. Stat. Softw. 2017, 80. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. Available online: https://www.R-project.org/ (accessed on 1 August 2019).
- Northrup, J.M.; Gerber, B.D. A comment on priors for Bayesian occupancy models. PLoS ONE 2018, 13, e0192819. [Google Scholar] [CrossRef] [PubMed]
- Peat, J.; Goulson, D. Effects of experience and weather on foraging rate and pollen versus nectar collection in the bumblebee, Bombus terrestris. Behav. Ecol. Sociobiol. 2005, 58, 152–156. [Google Scholar] [CrossRef]
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef] [Green Version]
- Bengio, Y.; Grandvalet, Y. No unbiased estimator of the variance of k-fold cross-validation. J. Mach. Learn. Res. 2004, 5, 1089–1105. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from Iterative Simulation Using Multiple Sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Zuur, A.F.; Leno, E.N. Beginner’s Guide to Zero-Inflated Models with R.; Highland Statistics Ltd.: Newburgh, NY, USA, 2016. [Google Scholar]
- Lesaffre, E.; Albert, A. Partial separation in logistic discrimination. J. R. Stat. Soc. Ser. B 1989, 51, 109–116. [Google Scholar] [CrossRef]
- López-Bellido, F.J.; López-Bellido, L.; López-Bellido, R.J. Competition, growth and yield of faba bean (Vicia faba L.). Eur. J. Agron. 2005, 23, 359–378. [Google Scholar] [CrossRef]
- Kambal, A.E. Flower drop and fruit set in field beans, Vicia faba L. J. Agric. Sci. 1969, 72, 131. [Google Scholar] [CrossRef]
- Link, W. Autofertility and rate of cross-fertilization: Crucial characters for breeding synthetic varieties in faba beans (Vicia faba L.). Theor. Appl. Genet. 1990, 79, 713–717. [Google Scholar] [CrossRef]
- Kambal, A.E.; Bond, D.A.; Toynbee-Clarke, G. A study on the pollination mechanism in field beans (Vicia faba L.). J. Agric. Sci. 1976, 87, 519. [Google Scholar] [CrossRef]
- Cartar, R.V. Adjustment of foraging effort and task switching in energy-manipulated wild bumblebee colonies. Anim. Behav. 1992, 44, 75–87. [Google Scholar] [CrossRef]
- Cartar, R.V.; Dill, L.M. Costs of energy shortfall for bumble bee colonies: Predation, social parasitism, and brood development. Can. Entomol. 1991, 123, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Garratt, M.P.D.; Powney, G.D.; Shaw, R.F.; Osborne, J.L.; Soroka, J.; Lindström, S.A.M.; Stanley, D.; Ouvrard, P.; Edwards, M.E.; et al. Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nat. Commun. 2019, 10, 1481. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Irwin, R.E.; Bronstein, J.L.; Manson, J.S.; Richardson, L. Nectar Robbing: Ecological and Evolutionary Perspectives. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 271–292. [Google Scholar] [CrossRef]
- Leadbeater, E.; Chittka, L. Social transmission of nectar-robbing behaviour in bumble-bees. Proc. Biol. Sci. 2008, 275, 1669–1674. [Google Scholar] [CrossRef] [Green Version]
- Sáez, A.; Morales, C.L.; Ramos, L.Y.; Aizen, M.A. Extremely frequent bee visits increase pollen deposition but reduce drupelet set in raspberry. J. Appl. Ecol. 2014, 51, 1603–1612. [Google Scholar] [CrossRef]
- King, C.; Ballantyne, G.; Willmer, P.G. Why flower visitation is a poor proxy for pollination: Measuring single-visit pollen deposition, with implications for pollination networks and conservation. Methods Ecol. Evol. 2013, 4, 811–818. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Heard, M.S.; Jitlal, M.S.; Rundlöf, M.; Bullock, J.M.; Shore, R.F.; Pywell, R.F. Replication, effect sizes and identifying the biological impacts of pesticides on bees under field conditions. J. Appl. Ecol. 2016, 53, 1358–1362. [Google Scholar] [CrossRef]
- Cresswell, J.E. A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology 2011, 20, 149–157. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Sato, S.; Peet, M.M.; Thomas, J.F. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 2000, 23, 719–726. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic exposure to neonicotinoids reduces honey bee health near corn crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, M.; Cerrutti, N.; Aupinel, P.; Decourtye, A.; Gayrard, M.; Odoux, J.-F.; Pissard, A.; Rüger, C.; Bretagnolle, V. Reconciling laboratory and field assessments of neonicotinoid toxicity to honeybees. Proc. Biol. Sci. 2015, 282, 20152110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreck, N.L.; Ratnieks, F.L.W. The dose makes the poison: Have “field realistic” rates of exposure of bees to neonicotinoid insecticides been overestimated in laboratory studies? J. Apic. Res. 2014, 53, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 2012, 491, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, R.J.; Raine, N.E. Chronic impairment of bumblebee natural foraging behaviour induced by sublethal pesticide exposure. Funct. Ecol. 2014, 28, 1459–1471. [Google Scholar] [CrossRef]
- Montoya, J.M.; Raffaelli, D. Climate change, biotic interactions and ecosystem services. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2013–2018. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Giannini, T.C.; Costa, W.F.; Cordeiro, G.D.; Imperatriz-Fonseca, V.L.; Saraiva, A.M.; Biesmeijer, J.; Garibaldi, L.A. Projected climate change threatens pollinators and crop production in Brazil. PLoS ONE 2017, 12, e0182274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogilvie, J.E.; Griffin, S.R.; Gezon, Z.J.; Inouye, B.D.; Underwood, N.; Inouye, D.W.; Irwin, R.E. Interannual bumble bee abundance is driven by indirect climate effects on floral resource phenology. Ecol. Lett. 2017, 20, 1507–1515. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2018, 21, 243–252. [Google Scholar] [CrossRef]
- Oliver, T.H.; Isaac, N.J.B.; August, T.A.; Woodcock, B.A.; Roy, D.B.; Bullock, J.M. Declining resilience of ecosystem functions under biodiversity loss. Nat. Commun. 2015, 6, 10122. [Google Scholar] [CrossRef] [Green Version]
- Kohler, M.; Devaux, C.; Grigulis, K.; Leitinger, G.; Lavorel, S.; Tappeiner, U. Plant functional assemblages as indicators of the resilience of grassland ecosystem service provision. Ecol. Indic. 2017, 73, 118–127. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Response Variable | Description | Reason for Inclusion |
|---|---|---|
| Legitimate pollination visitation rate (number of visits per 5 min interval) | Legitimate visits were classed when a bee entered the front of the flower to forage; this behaviour is most likely to lead to pollination [36]. | Often used measure of bee pollination services [23,37,38]. |
| Non-forage visitation rate (number of visits per 5 min interval) | This is the total number of visits to a plant where a bee did not actively forage. | Provides a measure of the level of activity carried out that does not provide any nutritional benefit to an individual or the colony. |
| Nectar rob (0 = no robbing, 1 = robbed) | Number of nectar robbing visits to a flower cluster. This was modelled as a binary response variable as the nectar robbing occurred in only 2.75% of visits and show little variation across the response. | Nectar robbing is unlikely to have a beneficial effect on pollination [36,39]. |
| Control vs. Bumble bee exposed plants | |
| Response variable | Model description |
| Probability of a plant producing pods (binary 0 = no pods produced and 1 = >0 pods produced) | Priors: Weakly informative Normal (0, 2.5) on intercept and fixed effect coefficients, this prior still allows extreme values but places a lower mass on them on the probability scale [44]. Default prior in brms for the random effects. Distribution: Bernoulli with logit link function As 70.73% of control plants produced no pods this was a binary response variable. |
| Colony stress treatment models | |
| Response variable | Model description |
| Proportional pod set per plant (n= 41) | Priors: Normal (0, 2.5) on intercept and fixed effect coefficients [44]. Default prior in brms for the random effects. Distribution: Binomial with logit link function. The proportion of flowers that turned into pods. |
| Total bean weight per plant (g) (n = 41) | Priors: Normal (0,100) on intercept and fixed effects. Default prior in brms for the random effects. Distribution: Normal. Log+1 transformed. The total weight of all beans per plant. |
| Number of beans per pods (binary 0 = 2 beans or less or 1 = > 2 beans per pod) (n = 124) | Priors: Normal (0, 2.5) on intercept and fixed effect coefficients [44]. Default prior in brms for the random effects. Distribution: Bernoulli with logit link. This variable was the number of beans produced per pod and followed a uniform distribution across the values 1–4 and was poorly modelled by a poisson or negative binomial response distribution. Included Plant ID random effect. |
| Behavioural Variable | Model | Difference in Validation Error | Standard Error of the Difference |
|---|---|---|---|
| Legitimate visitation rate | Interaction effects model Main effects model | 0 −3.52 | 0 5.1 |
| Non-foraging visitation rate | Main effects model Interaction effects model | 0 −1.38 | 0 2.17 |
| Probability of nectar robbing | Interaction effects model Main effects model | 0 −4.31 | 0 2.94 |
| Behavioural Variables | Parameter | Estimate | Lower 95%CI | Upper 95%CI |
|---|---|---|---|---|
| Legitimate visitation rate (visits/5 min period) | Intercept Heat H+ Insecticide I+ | 0.31 0.37 0.03 | −1.56 −0.95 −1.21 | 2.09 1.64 1.24 |
| Non-foraging visitation rate (visits/5 min period) | Intercept Heat H+ Insecticide I+ | −0.85 0.64 0.30 | −2.17 −0.15 −0.34 | 0.36 1.46 0.94 |
| Probability of nectar robbing (0 = not robbed and 1 = robbed) | Intercept Heat H+ Insecticide I+ | −3.36 −1.79 1.44 | −6.15 −4.36 −0.82 | −0.86 0.53 3.98 |
| Yield Variable | Model | Difference in Validation Error | Standard Error of the Difference |
|---|---|---|---|
| Proportional pod set | Main effects model | 0 | 0 |
| Interaction effects model | −5.28 | 1.66 | |
| Total bean weight | Main effects model | 0 | 0 |
| Interaction effects model | −3.69 | 1.78 | |
| Probability of a pod producing >2 beans | Interaction effects model Main effects model | 0 −0.32 | 0 2.74 |
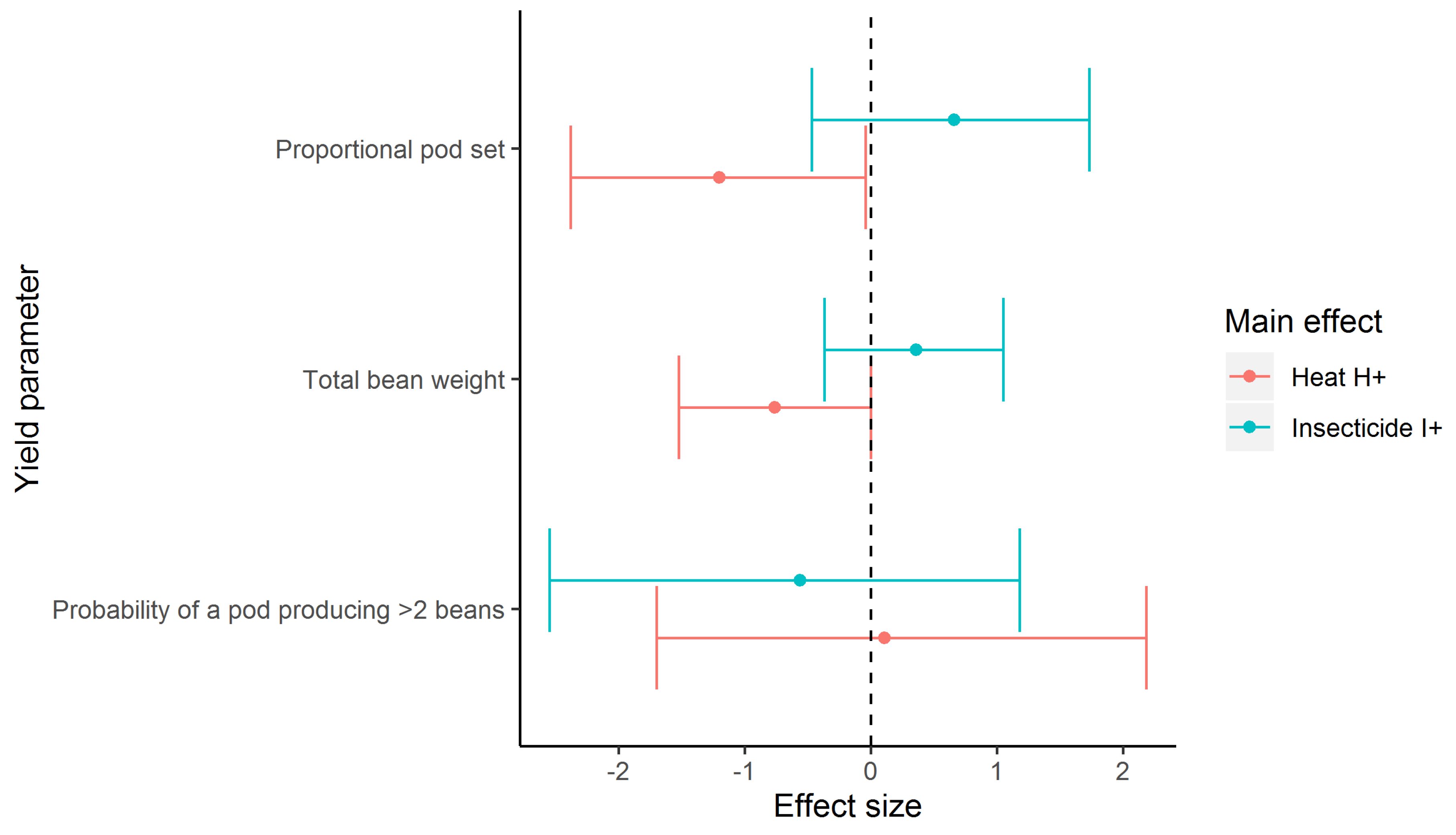
| Yield Variable | Parameter | Estimate | Lower 95%CI | Upper 95%CI |
|---|---|---|---|---|
| Proportional pod set | Control | −1.69 | −2.88 | −0.50 |
| Heat H+ | −1.20 | −2.38 | −0.04 | |
| Insecticide I+ | 0.66 | −0.47 | 1.73 | |
| Total bean weight | Control | 1.41 | 0.66 | 2.18 |
| Heat H+ | −0.76 | −1.52 | 0.00 | |
| Insecticide I+ | 0.36 | −0.37 | 1.05 | |
| Probability of a pod producing > 2 beans | Control Heat H+ Insecticide I+ | −0.19 0.11 −0.56 | −2.04 −1.70 −2.55 | 1.54 2.18 1.18 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greenop, A.; Mica-Hawkyard, N.; Walkington, S.; Wilby, A.; Cook, S.M.; Pywell, R.F.; Woodcock, B.A. Equivocal Evidence for Colony Level Stress Effects on Bumble Bee Pollination Services. Insects 2020, 11, 191. https://doi.org/10.3390/insects11030191
Greenop A, Mica-Hawkyard N, Walkington S, Wilby A, Cook SM, Pywell RF, Woodcock BA. Equivocal Evidence for Colony Level Stress Effects on Bumble Bee Pollination Services. Insects. 2020; 11(3):191. https://doi.org/10.3390/insects11030191
Chicago/Turabian StyleGreenop, Arran, Nevine Mica-Hawkyard, Sarah Walkington, Andrew Wilby, Samantha M Cook, Richard F Pywell, and Ben A Woodcock. 2020. "Equivocal Evidence for Colony Level Stress Effects on Bumble Bee Pollination Services" Insects 11, no. 3: 191. https://doi.org/10.3390/insects11030191