Protura in Arctic Regions, with Description of Mastodonentomon n. gen. (Acerentomidae, Nipponentominae) and a Key to Known Arctic Taxa


1
Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, Poland
2
State Museum of Natural History, Ukrainian National Academy of Sciences, Teatral’na St. 18, UA 79008 L’viv, Ukraine
3
Institute of Soil Biology, Biology Centre Czech Academy of Sciences, Na Sádkách 7, 370 05 České Budějovice, Czechia
4
Institute of Nature Conservation, Polish Academy of Sciences, Mickiewicza 33, 31-120 Kraków, Poland
5
University of Tennessee, Entomology and Plant Pathology, 2505 E.J. Chapman Drive, 370 Plant Biotechnology, Knoxville, TN 37996, USA
*
Author to whom correspondence should be addressed.
Insects 2020, 11(3), 173; https://doi.org/10.3390/insects11030173
Submission received: 31 January 2020
/
Revised: 4 March 2020
/
Accepted: 4 March 2020
/
Published: 9 March 2020
(This article belongs to the Special Issue Polar Entomology)
Abstract
:Protura are widespread, but their presence in the Arctic was first noted only ca. 70 years ago and is still little acknowledged. This work compiles taxonomic information on proturans in the Arctic regions and adds unpublished data from Northern Siberia. Currently, this fauna is represented by 23 species in two orders and 14 genera. The large cosmopolitan genus Eosentomon is represented by only four species, whereas Acerentomidae is much more diverse, with 19 species in 13 genera (eight Nipponentominae, five Acerentominae). Most of the Arctic species possess a larger number of setae than species living in temperate regions. Based on several unique characters, a new genus, Mastodonentomon, is erected for Nipponentomon macleani, and the species is re-described with the original description supplemented with new characters, including head chaetotaxy, seta length, and porotaxy. Proturan occurrence in the Arctic is limited to Beringia, but the majority of species have restricted distributions and none have been found in both the American Arctic and Siberia. This implies relict origins and high levels of proturan endemism in the Arctic. This emerging view on biogeographical history is, however, hampered by the limited extent of available data, which highlights the need for considerably greater survey efforts. A key to Arctic proturans is provided to facilitate further studies.
Keywords:
proturans; arthropods; chaetotaxy; porotaxy; distribution; biogeography; Alaska; Siberia; Northern Canada; Beringia1. Introduction
Protura, known as coneheads, are among the smallest of soil-dwelling microarthropods. These hexapods are wingless, eyeless, and have slender elongate bodies ranging between 0.7–2 mm. Due to their minuteness and cryptic edaphic lifestyle, they are easily overlooked and were discovered relatively late in the history of entomology [1]. Originally regarded as an order belonging to the class Insecta [2], Protura was later placed among the basal hexapod taxa as the sister taxon to either Collembola or Diplura. Their phylogenetic position, however, is still a point of controversy [1,3]. Since their first description in 1907 [2], over 800 valid species belonging to three orders, seven families, and 76 genera have been described worldwide [1,4,5]. However, this number likely represents only ca. 10% of the world’s species [6].
Proturans usually live alongside other microarthropods, such as Acari, Collembola, Pauropoda and Symphyla, with which they form an essential component of edaphic communities in all terrestrial regions and climatic zones, and elevations ranging from sea level to the treeless zone on high mountains [1,7]. Their distribution seems to be limited only by the presence of sufficient moisture levels and the availability of decaying organic matter [7,8]. Although proturans are sometimes common, they are infrequently collected, usually only as a by-catch of other arthropods. Thus, their diversity and geographical and ecological distributions are still little known [1,7].
Some authors have also pointed to the general lack of proturans in polar regions and nival zones of high mountains [1,4,9]. While proturans have never been found in the Antarctic and Sub-Antarctic regions, they are recorded above the Arctic Circle in tundra and taiga biomes, reaching past 72° N. However, only a few papers concern the study of proturan fauna in the Arctic regions. For instance, until now, only Tuxen [10], Nosek [11,12,13], and Bernard [14] have documented the occurrence of Protura in Alaska and the Canadian Arctic. Nosek [13] also summarized the data on proturan fauna and presented a key for the identification of all proturan species known at that time from Alaska. Data about Arctic Protura from the Palearctic region also are very scarce [15,16,17,18]. Recent advances in proturan taxonomy highlight the need for taxonomic revisions, and several species from Alaska and Northern Canada have been re-described and transferred to other genera [19,20,21,22].
In this work, all data on proturans from the Arctic regions available from the literature are reviewed and summarized in order to get a clearer idea of the true diversity and distribution of these arthropods. Proturans from soil and moss samples collected recently in Northern Yakutia (Siberia) were also extracted and analysed, which significantly extends the known northernmost limits of their distribution. Furthermore, an integrative re-description of Nipponentomon macleani Nosek, 1977 is provided and a new genus, Mastodonentomon, is established for this species. A key for determination of all proturans known from the Arctic regions is also provided to facilitate further studies.
2. History of Proturan Research in the Arctic
Protura were first collected inside the Arctic Circle by M. Hammer, who, in 1948, collected well over 100 specimens from two different locations above 68° N in Northern Canada [10]. The majority of these proturans were described as Acerentulus canadensis Tuxen, 1955, and six specimens from one sample as Nosekiella condei Tuxen, 1955 (Table 1). Currently, these species are in the genera Verrucoentomon and Vesiculentomon, respectively [19,22]. Hammer’s collection contained one specimen belonging to the genus Eosentomon. It was, however, a “larva 2” and, therefore, indeterminable to species [10]. Weber [23] mentioned a record of “Acerentulus sp. (Family Acerentomidae)” collected in 1949 from two locations in Alaska, extending the northernmost proturan record past 69° N. Then it was not until summer 1976 when a notable collection of proturans from a number of localities across Alaska was made by S.F. MacLean and A. Fjellberg (Institute of Arctic Biology, Fairbanks AK) [11] with the northernmost localities above 70° N. Based on that collection, Nosek [11,12,13] described 14 species, including three Eosentomidae and 11 Acerentomidae, and wrote a key [13] for determining all proturan species known from Alaska. Lastly, Bernard (1985) [14] described Eosentomon adakense from samples collected in 1978 on Adak Island (Aleutian Islands, Alaska). Some of the Acerentomidae species have been transferred to other genera: Orinentomon, Sugaentulus, and Imadateiella [19,20,24]. The genus Nosekientomon was erected for the species Vesiculentomon ruseki Nosek, 1977 [19]. Two Alaskaentomon species and Verrucoentomon imadatei Nosek, 1977 are valid and were re-described recently [21,22]. Revision of the type material of Nipponentomon macleani Nosek, 1977 shows that this species has some unique characters within Acerentomidae and a new genus has to be established for it.
The first known Arctic proturan in the Palearctic was collected from Snow Valley in the Magadan district in Siberia: Acerella sharovi Martynova, 1977 [15]. This species was transferred to Imadateiella by Imadaté [25]. Three proturan species were described from the taiga belt near Turukhansk City, close to the Arctic Circle [16,18,26], and Nienna chukotka Shrubovych, 2019, was described from Northern Chukotka above 69° N [27]. This record and a new record of Yamatentomon yamato Imadaté & Yosii, 1956, from northern Yakutia above 72° N, are currently the northernmost known Palearctic localities of Protura.
3. Materials and Methods
3.1. Faunistic Dataset
A dataset of the distribution of proturans in Arctic regions was created based on the publications concerning these species. In addition, new data from Northern Yakutia (Siberia) reported in the current work were also included. Geographically, the dataset covers all studies conducted in the Arctic, as defined by the Arctic Monitoring and Assessment Programme (AMAP) [28], including all the terrestrial areas north of the Arctic Circle (66°33′ N) as well as most areas north of 60° N in Asia and North America with continuous permafrost, and the Aleutian Islands. It should be borne in mind that a significant proportion of these areas are often defined as Subarctic, but we follow the AMAP terminology for simplicity.
The dataset (Table 1) includes: (1) the valid species names and synonyms, and their taxonomic position, (2) sites where proturans were collected and their coordinates (with the type locality marked with *), (3) type of habitat in which they were found, as given in the original descriptions, (4) remarks on their known distribution, and (5) literature sources of the original records and species re-descriptions if available.
3.2. Material Examined
Proturan specimens from Northern Yakutia (Siberia) were extracted from soil and moss samples with Berlese-Tullgren funnels into 95% ethanol. The specimens were mounted on glass slides in Faure’s medium [29] for taxonomic evaluation. The examined specimens were deposited in the collection of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków (ISEA). In addition, the type material (holotype female and paratype male) of Nipponentomon macleani, preserved in the National Museum of Natural History (Smithsonian Institution) in Washington, DC, USA, was also studied using oil immersion and phase contrast on an Olympus microscope.
The terminology for body chaetotaxy and porotaxy follows Szeptycki [30] and Shrubovych [18]. The head seta designations follow Rusek et al. [31]. Abbreviations used in the description are as follows: Abd. = abdominal segments, Th. = thoracic segments, A-setae = anterior setae, M-setae = medial setae, P-setae = posterior setae, al = anterolateral, sl = sublateral, psm = posterosubmedial, psl = posterosublateral, spm = sternal posteromedial, and spsm = sternal posterosubmedial cuticular pores.
4. Results
4.1. Faunistic Dataset
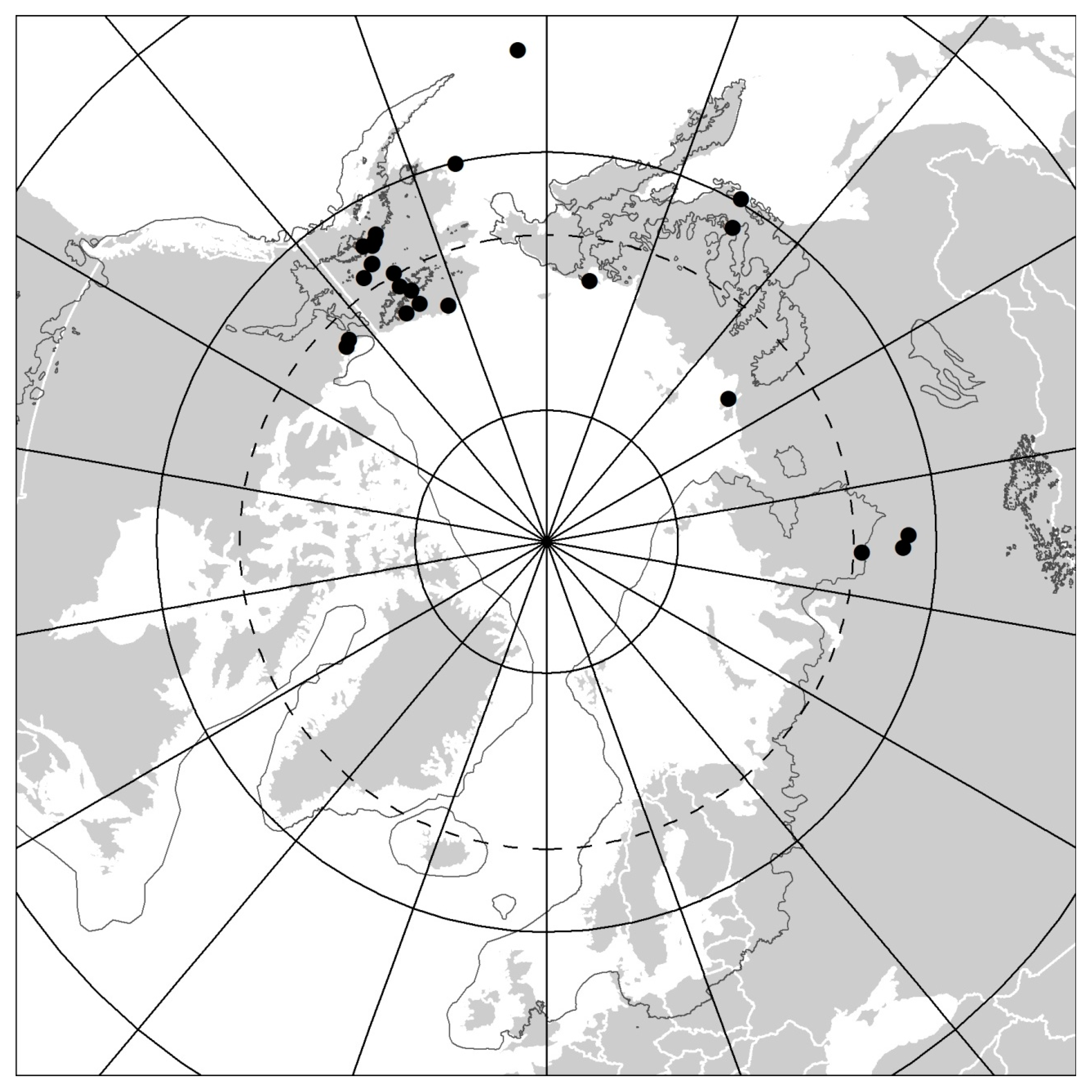
The faunistic dataset contains 37 records of Protura from 24 sites within the Arctic regions (Table 1), published originally in 11 papers. Of these, 26 records from 17 sites and 6 papers [10,11,12,13,14,23] concern Alaska and the Canadian Arctic and only 11 records from 7 sites and 5 papers [15,16,17,26,27] concern the Siberian Arctic. No records were found from the European Arctic (Figure 1). Overall, 23 proturan species from two orders (Eosentomata and Acerentomata) and 14 genera were identified from both the North American and Siberian Arctic (17 and 6 species, respectively). The order Eosentomata is represented by only four species, which all belong to the genus Eosentomon. Acerentomata are considerably more diverse, with 19 species belonging to 13 genera (eight Nipponentominae and five Acerentominae). Moreover, two records undetermined to species level, both reported as “Acerentulus sp. (Family Acerentomidae)” [23], were listed from Northern Canada. Unfortunately, Weber [23] did not provide any descriptions and whereabouts of these collections are unknown, so they could not be re-examined. It should also be remembered that, since that time, Acerentulus has been split into several genera. Thus, at present, these records can be assigned, at best, to Acerentomidae only.
Species richness for particular sites ranged from one to four even though the majority of sites (18) hosted only single species and only three sites (i.e., Chena Ridge and Denali National Park in Alaska, and Turukhansk in Siberia) had four species. However, it should be borne in mind that these estimates are mostly based on historical data taken from the literature and they might be biased by the sampling effort. Nearly all species reported from Alaska and the Canadian Arctic (i.e., 13 of 17 species) are known only from their type localities and only four were also found in other sites across this region. On the other hand, among Siberian species, only Nienna chukotka Shrubovych, 2019 is known exclusively from its type locality. The other five species seem to have broader distributions across Siberia and records of Yamatentomon yamato (Imadaté and Yosii, 1956), reported in the present paper from the northernmost locality, span a wide latitudinal range across the Northeastern Palearctic. It is also remarkable that none of the species have been found in both regions.
4.2. New Record from the Arctic Region
Yamatentomon yamato (Imadaté & Yosii, 1956).
Material Examined: 2 females, 9 males, 1 maturus junior, 4 larvae II, 7 larvae I, soil and moss in tundra. Russia, Sakha-Yakutia, Tyallakh, Lena Delta Nature Reserve, sedge/moss tundra near the reserve station, 72°19′36″ N, 128°07′27″ E, 16.08. 2015., coll. A. Nekhaeva.
Distribution: Japan, Korea, Northeastern China, Russian Far East [4,18] and Northern Siberia (current study).
Remarks: The specimens are identical with more southern specimens (see References [18,33]) in body length, chaetotaxy, and porotaxy. The sensilla pattern on the foretarsus is similar, with the very long sensillum b surpassing the seta γ4 insertion point. Unlike southern specimens, foretarsal sensillum a is shorter. Its apex reaches to the sensillum t2 insertion, and does not reach the base of sensillum d. This small character difference is not enough to justify a description of a new species.
4.3. Systematics
4.3.1. Mastodonentomon Shrubovych, New Genus
Characterized by the presence of four pairs of A-setae on meso- and metanota (A1, A2, A3, A4), setae M2 and M3 on metanotum, six pairs of A-setae on tergites I–III (additional setae A1’ present), five pairs of A-setae on tergites IV–VII, filiform foretarsal sensillum t1, sensillum b’ missing, position of foretarsal sensillum b proximal to c insertion, d situated closer to e than to c, sensillum a’ in distal position, close to c’ insertion, well-developed maxillary, and labial palps with terminal tuft of setae, posterior position of P3 setae on tergites II–VI, well-developed striate band on segment VIII with distinct parallel-sided striae, 4/2 setae on sternite VIII, and presence of two setae of equal length on abdominal legs II and III.
Type species:Mastodonentomon macleani (Nosek, 1977).
Remarks: The new genus Mastodonentomon is placed into subfamily Nipponentominae due to possessing a distinct calyx with large vesicle and racemose appendices on its surface, presence of four pairs of A-setae on metanotum, presence of 4/2 setae on sternite VIII, well-developed striate band on segment VIII, and two setae of equal length on abdominal legs II and III [5]. The new genus is similar to Nipponentomon and Imadateiella in having four pairs of A-setae on metanotum and filiform foretarsal sensillum t1. The genus differs in the presence of five pairs of A-setae on tergite VII and in the position of foretarsal sensillum a’, which is placed far distally, close to c’ insertion, as in some Acerentomon spp. [34,35]. The positions of sensillum a’ on the foretarsus clearly separates most of the genera within Nipponentominae [19]. This new genus differs from all other Acerentomidae in the presence of four pairs of A-setae on the mesonotum (A1, A2, A3, A4) and three pairs of M-setae (M1, M2, M3) on the metanotum. Other Protura have only two or three pairs of A-setae on mesonotum and one pair of M-setae on the metanotum. These characters are important at the generic level and justify the erection of Mastodonentomon n. gen. within the subfamily Nipponentominae.
4.3.2. Mastodonentomon macleani (Nosek, 1977), New Combination
syn. Nipponentomon macleani Nosek, 1977
Material Examined: Holotype female (no. 75801), collected in Dryas on exposed cliff, Mastodon Dome at Eagle Creek, 1200 m elev., Alaska, 1.VIII.1976, coll. A. Fjellberg. Paratype male collected together with the holotype. The slides are mounted in Swan’s medium and stored in the National Museum of Natural History (Smithsonian Institution) in Washington, USA.
Diagnosis: Large species with a body length of more than 1500 μm. Setae A1 present on mesonotum, metanotum, and metasternum, setae M2 and M3 on metanotum, six pairs of A-setae on tergites I–III, five pairs of A-setae on tergite VII, five A-setae on sternites I–VI and three A-setae on sternite VII. Additional cephalic seta d6 very long; sensilla f and c’ the longest on foretarsus. Pores al and sl present on meso- and metanota, group of sc pores present on mesosternum, two separate sc pores present on metasternum, psl present on tergites I–VII, pore spm present on sternites I–VII, a pair of sal pores present on sternite I, and a pair of spsm pores on sternites VI–VII.
Re-description: The maxillary and labial palpi, canal of the maxillary gland, arrangement of the foretarsal sensilla, setae of the third abdominal legs, striate band and comb on abdominal segment VIII, female squama genitalis were all accurately depicted by Nosek [11] to which the reader is referred for illustrations.
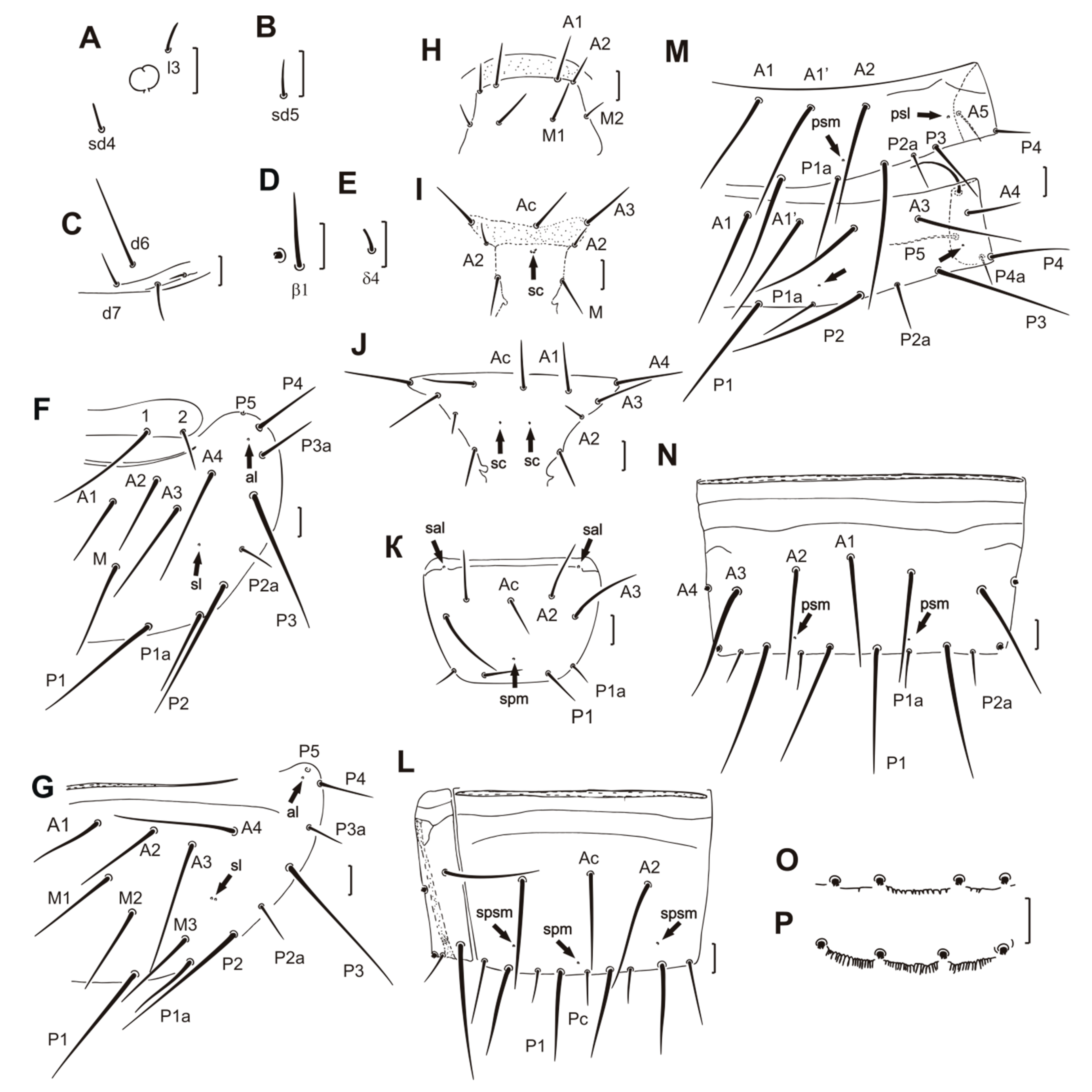
Head setae l3, sd4, and sd 5 setiform, 15, 13, and 16 μm in length, respectively (Figure 2A,B). Additional setae d6 very long, 65–70 μm (Figure 2C). Labrum not protruded. LR = 16. Pseudoculus round with a short posterior extension. PR = 21. Calyx of maxillary gland with two large racemose appendices. Posterior filament long, with simple posterior dilation. CF = 5.0–5.2 (see Reference [11]: Figure 6E). Maxillary palp with terminal tuft of setae. Sensilla short and pointed apically, similar in length and shape (see Reference [11]: Figure 6C). Labial palps with long, broad basal sensillum (see Reference [11]: Figure 6C).
On foretarsus, sensillum t1 filiform, t3 small and leaf-like. f and c’ very long, nearly setiform (see Reference [11]: Figure 6A,B). Sensillum b shorter than c and situated proximally to level of c insertion, g short and slender. Sensillum d closer to e than to c, a’ in distal position, close to c’ insertion, b’ missing. Length formula of sensilla: t3 < t1 < (b = g) < (a = d) < (t2 = c = e = a’) < c’ < f. Seta β1 setiform and long, seta δ4 short and stout, their length 32 μm and 10 μm, respectively (Figure 2D,E). Claw long, with long inner tooth, empodial appendage long. BS = 0.6–0.8, TR = 2.4–2.6, EU = 0.14–0.15. Single pores near bases of sensilla c and t3.
Chaetotaxy formula given in Table 2. Setae on nota differing in length (Figure 2F). Pronotal seta 1 three times longer than seta 2. Meso- and metanota with seta A1. Seta P1a on meso- and metanota long, setiform, seta P2a short, half length of seta P1a; P2a situated midway between P2 and P3. Length ratio of mesonotal setae P1:P1a:P2 as 1.6:1:1.8. Mesonotum with a pair of single sl and al pores and metanotum with a pair of doubled sl pores and a pair of al pores (Figure 2F,G). Prosternum without pores, mesosternum with a group of three closely placed sc pores, metasternum with a pair of separately placed sc pores (Figure 2H–J). Seta A2 on thoracic sterna and P4 on tergite I short, setiform (Figure 2I,J,M).
Tergites I–III with additional A1’ setae (Figure 2M). All accessory setae on tergites I–VI setiform. Setae P1a longest on tergite I (60 μm), shorter on tergites II–VI (about 43 μm), and shortest on tergite VII (37 μm). Setae P2a shorter than P1a, their length on tergite I 27 μm, on tergites II–VI 34 μm. Setae P4a the shortest when compared to other accessory setae, their length 20 μm on tergite II and 25 μm on tergites III–VII. Setae P2a and P4a on tergite VII 25 μm long. Pores psm and psl present on tergites I–VII anterior to P1a, al on tergites II–VII (Figure 2M,N).
Abdominal legs with 4, 2, 2 setae. Subapical and lateral apical setae on second and third pairs of abdominal legs nearly equal in length, 25 and 23 μm, respectively (see Reference [11]: Figure 6F). Lengths of accessory setae P1a on sternites I–III 17, 20, and 23 μm, respectively, of equal length (26 μm) on sternites IV–VII. Sternites I–V with single spm pores, sternite I with a pair of sal pores (Figure 2K). Sternites VI–VII with spm pores and a pair of spsm pores anterior to P2 (Figure 2L).
Abdominal segment VIII with a distinct striate band. Tergite and sternite each with anterior, irregular, transverse row of small teeth (see Reference [11]: Figure 7). Pore psm with several accompanying teeth. Posterior margin of sternite VIII and laterotergites smooth. Comb VIII with 9–10 long teeth (see Reference [11]: Figure 6G). Sternites IX–X with central setae half the length of lateral setae (see Reference [11]: Figure 6H). Tergite IX with minute serrations in the central part of the hind margin (Figure 2O), tergites X–XI, and sternite X with distinct serrations on the entire posterior margin (Figure 2P). Sternite IX with serrations on the lateral part of the hind margin, sternite X with serrations on the entire posterior margin (see Reference [11]: Figure 6H). Dorsal lobe of tergite XII with a median pore and a ventral lobe with 1+1 anterolateral pores.
Female squama genitalis with long, apically bipartite acrostyli (see Reference [11]: Figure 6I). Male squama genitalis with 6 + 6 setae.
Measurements: 2 adults (in μm): total length 1570–1660, head 235, pseudoculus 11, posterior part of maxillary gland 45–47, posterior cephalic setae: seta d6 65–70, d7 21–22, seta sd7 25–26, seta l5 9, pronotal seta 1 84–80, pronotal seta 2 27; mesonotal seta P1 92, mesonotal seta P1a 54–56, mesonotal seta P2 103, foretarsus 150–151, claw 58–60, empodial appendage 10–12.
Chaetal variability: In the holotype, seta A1 doubled on metasternum, Pc doubled on sternite III, A1 absent asymmetrically on tergite VII. In paratype, seta A1’ present symmetrically on tergite IV.
Etymology: The generic name is taken from the general locality (Mastodon Dome) where the specimens were collected.
Remarks: In this re-description of Nipponentomon macleani, morphological characters are added to the previous description of the species, such as chaetotaxy of head, length of seta on head, nota and foretarsus, and porotaxy on tergites and sternites. According to Nosek [11] tergite I has 6 A-setae and 14 P-setae. However, re-examination of the type material clearly showed that tergite I has 8 A-setae (seta A5 present) and 12 P-setae.
4.4. Key to Arctic Protura
| 1. | Meso- and metanotum with a pair of spiracles | ………………(Eosentomon Berlese, 1908) … 20 |
| – | Meso-metanotum without spiracles | …………(Acerentomidae Silvestri, 1907) … 2 |
| 2. | Metanotum with two pairs of A-setae | ………………………… 3 |
| – | Metanotum with three or four pairs of A-setae | ……… 8 |
| 3. | Abdominal legs II and III with two setae | ……………………………………… 4 |
| – | Abdominal legs II and III with three setae | ………… Fjellbergella tuxeni Nosek, 1980 |
| 4. | Striate band on tergite VIII well developedwith distinct striae | ………………………… 5 |
| – | Striate band reduced to two lineswithout striae | …………… Tuxenentulus boedvarssoni Nosek, 1981 |
| 5. | Foretarsal sensillum t1 filiform | ………………(Alaskaentomon Nosek, 1977) … 6 |
| – | Foretarsal sensillum t1 claviform | ……………(Sugaentulus Imadaté, 1978) … 7 |
| 6. | Sternites II–III with five A-setaetergite VII without seta P3a | ………… Alaskaentomon condei Nosek, 1981 |
| – | Sternites II–III with three A-setaetergite VII with seta P3a | ……… Alaskaentomon fjellbergi Nosek, 1977 |
| 7. | Tergites I–VI with seta P1a, sternites IV and VI with four A-setae | …………………Sugaentulus hoogstraali (Nosek, 1980) |
| – | Tergites I–VI without seta P1a, sternites IV and VI with three A-setae | …………………Sugaentulus andrzeji Shrubovych & Rusek, 2010 |
| 8. | Metanotum with three pairs of A-setae | ………………………… 9 |
| – | Metanotum with four pairs of A-setae | ………………………… 15 |
| 9. | Foretarsal sensillum t1 filiform | ………………………… 10 |
| – | Foretarsal sensillum t1 claviform | ………………………… 13 |
| 10. | Calyx of a maxillary gland with small granulated appendicesforetarsal sensillum t3 globular | ………………………… Nienna chukotka Shrubovych, 2019 |
| – | Calyx of a maxillary gland with large granulated appendicesforetarsal sensillum t3 leaf-like | ………………………… (Verrucoentomon Rusek, 1974) … 11 |
| 11. | Sternite I without setae P1a, sternite VII with seta Pc | ………………………… 12 |
| – | Sternite I with setae P1a, sternite VII without seta Pc | ……… Verrucoentomon anatoli Shrubovych & Bernard, 2012 |
| 12. | Foretarsal sensilla b and c equal in length | ……… Verrucoentomon canadense (Tuxen, 1955) |
| – | Foretarsal sensillum b clearly longer than c | ……… Verrucoentomon imadatei Nosek, 1977 |
| 13. | Maxillary gland with smooth globular vesicle near calyx | ……… Nosekientomon ruseki (Nosek, 1977) |
| – | Maxillary gland without vesicle near calyx | ………(Yavanna Szeptycki, 1988) … 14 |
| 14. | Foretarsal sensillum a longits apex reaching the base of seta γ3 sensillum g thin | ……… Yavanna behanae (Nosek, 1977) |
| – | Foretarsal sensillum a shorterits apex not reaching base of seta γ3 sensillum g thickened | ……… Yavanna babenkoi ShrubovychRusek & Bernard, 2012 |
| 15. | Foretarsal sensillum t1 filiform | ……… 16 |
| – | Foretarsal sensillum t1 baculiform or claviform | ……… 18 |
| 16. | Foretarsal sensillum a’ in distal positioninsertion close to base of sensillum c’ | ……… Mastodonentomon macleani (Nosek, 1977) |
| – | Foretarsal sensillum a’ in proximal positioninsertion near base of sensillum t2 | ……… (Imadateiella Rusek, 1974) … 17 |
| 17. | Tergite VII with three pairs of A-setae, sternite VI with Pc setasternite VII with six setae | ……… Imadateiella sharovi (Martynova, 1977) |
| – | Tergite VII with two pairs of A-setae, sternite VI without Pc setasternite VII with four setae | ……… Imadateiella mixta (Nosek, 1981) |
| 18. | Foretarsal sensillum t1 baculiform | ……… 19 |
| – | Foretarsal sensillum t1 claviform | ……… Orinentomon greenbergi (Nosek, 1980) |
| 19. | Sternite VII with six setaesternite VI without Pc seta | ……… Vesiculentomon condei (Tuxen, 1955) |
| – | Sternite VII with four setaesternite VI with Pc seta | ……… Yamatentomon yamato Imadaté & Yosii, 1956 |
| 20. | Foretarsal sensillum b’1 presentsternite VIII without A-setae | ……… 21 |
| – | Foretarsal sensillum b’1 absentsternite VIII with a pair of A-setae | ……… Eosentomon aquilinum Nosek, 1980 |
| 21. | Tergite VIII with three pairs of A-setae | ……… 22 |
| – | Tergite VIII with two pairs of A-setae | ……… Eosentomon adakense Bernard, 1985 |
| 22. | Foretarsal sensilla t2 and b’2 spatulate | ……… Eosentomon copelandi Nosek, 1980 |
| – | Foretarsal sensilla t2 and b’2 setiform | ……… Eosentomon alaskaense Nosek, 1977 |
5. Discussion
The proturan fauna of the Arctic regions is not rich but is quite diverse, represented by two orders (Eosentomata and Acerentomata), 14 genera and 23 species (Table 1). The order Sinentomata has not been recorded from this region, but only five species are known at present [4]. Considering over 800 species of Protura known worldwide [1,4,18], the proportion of the global proturan diversity found in the Arctic is very low (less than 3%). Species richness recorded for particular sites was also very low and the majority of sites hosted only single species. Nevertheless, nearly 20% of 76 genera known worldwide [5] have been found in the region. Members of the widespread genus Eosentomon, which are very abundant and diverse in the Americas and Europe [36,37,38], are poorly represented, with only four known species, and restricted in distribution to Alaska and Northern Canada (Table 1). However, the lack of Eosentomon in the Siberian Arctic is not surprising because species of this genus are generally rare and not numerous in neighboring regions [18,39]. Acerentomidae is very diverse in the Arctic at the generic level and comprise eight genera of Nipponentominae and five of Acerentominae. The genus Verrucoentomon is most diverse in the Arctic fauna. Its members have usually been collected in mountainous regions, such as V. montanum (Martynova, 1970) and V. rafalskii Szeptycki, 1997. Such apparently relict species may have been able to survive on non-glaciated mountaintops [18,40]. The subfamilies Berberentulinae and Acerellinae apparently are absent from the Arctic regions. Berberentulinae is rich in species and widely distributed. Acerellinae is restricted to four species know only from Europe [4]. Members of these subfamilies are characterized by a reduction of some morphological characters, e.g., shape of labial palps, striate band, number of setae on body segments, and sensilliform accessory setae on the body. The acerentomids on the northern edge of proturan distribution possess well-developed labial palps with a terminal tuft of setae, well-developed striate band on segment VIII (except Fjellbergella and Tuxenentulus spp.), and a larger number of setae on the body than species with a more southern distribution.
In general, Protura can be found wherever decaying organic matter and sufficient moisture levels are available [1,8], and often are restricted to habitats characterized by luxuriant vegetation growth [7]. It is believed that proturans are fungivorous and some species may feed preferentially on ectomycorrhizal fungi [41]. It is not surprising, therefore, that proturans have rarely been collected in tundra biome, which marks the northern limit of their distribution. They have never been collected in the taiga biome in the European part of Russia, but proturans occur in Siberian taiga and tundra biotopes [42].
Alaska, Yukon, and Siberia are a part of the Beringian region that was mostly ice-free during the Pleistocene glaciations [32] and, therefore, proturans are found further north there than in the rest of the Holarctic. This suggests the importance of historical biogeography in the distribution of proturan species, which was a hypothesis highlighted by Tuxen [43]. Furthermore, records suggest pronounced biogeographical differences in distribution of proturan species within the Beringian region. For instance, no species were found in both the North American and Siberian Arctic. This disparity implies the existence of a strong intercontinental disjunction in proturan distributions between these two Arctic regions. Moreover, the majority of species seem to have regionally restricted distributions or are known only from their type localities. This pattern is particularly pronounced for the proturan fauna in North American Arctic, but also among Siberian species. Only Yamatentomon yamato is widely distributed over a broad latitudinal range extending far beyond the Arctic region (see Reference [18]). While substantial northward migration and post-glacial recolonization from the south is apparent from datasets for some previously glaciated regions, e.g., Southern Canada [44] or Southern Fennoscandia [4], it appears that further north in the Arctic regions distribution of proturans is still limited to the areas of Beringia that were unglaciated during the Pleistocene (Figure 1). Moreover, the restricted distributions and morphological differences among some taxa, when compared to species with a more southern distribution (as indicated above), imply a relict origin and a potentially very high level of endemism in proturan fauna of that region. This hypothesis is consistent with distribution patterns reported for some other arthropod taxa, particularly those with limited dispersal capabilities (see Reference [45]), which were presumably eliminated or prevented from dispersing by glaciers elsewhere, but for which Beringia acted as a refugium. However, the available data on proturan occurrence are fragmentary and restricted to very few specific collection localities. Many areas have not been sampled at all. Thus, the apparent restricted distributions may, in part, reflect the limited sampling intensity and location of specific collection sites rather than the real distribution of these animals. While our work provides both a benchmark for the region and the foundation for future research, considerably greater survey efforts are necessary to elucidate the true diversity of proturans and biogeographical patterns of their distribution in the Arctic regions.
6. Conclusions
Although proturans are often believed to be absent in polar regions, their occurrence above the Arctic Circle was noted more than 70 years ago with the northernmost records exceeding 72° N. While early works focused on documenting and describing new species, primarily in Alaska and the Canadian Arctic, recent advances in proturan taxonomy have facilitated re-descriptions of several taxa and documented the occurrence of proturans throughout Northern Siberia. Accordingly, in the current work, we re-describe Nipponentomon macleani, supplementing the original description with new characters, including head chaetotaxy, seta lengths, and porotaxy, and, on the basis of several unique characters, establish a new genus, Mastodonentomon, for this species. Nonetheless, the Arctic literature dataset contains only 35 records representing 23 species, i.e., less than 3% of the global proturan diversity. In contrast, generic diversity is relatively high but dominated by Acerentomidae. Another striking feature of the collected dataset is that occurrence of proturans in the Arctic regions appears to be limited to regions of Beringia that were glacier-free during the Pleistocene. Emerging patterns in species distribution and their distinct morphology also imply a relict origin and a potentially high level of endemism within the proturan fauna of that region. It is clear, however, that proturans in the Arctic regions have been studied very selectively. Hence, consideration of the importance of biogeographical history and the concept of endemism is itself strongly hampered by the limited extent of available data, which highlights the need for considerably greater survey efforts. Still, this work provides a strong baseline needed to facilitate further studies addressing diversity and biogeography of these little-known arthropods in the Arctic regions and elsewhere.
Author Contributions
Conceptualization, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Methodology, J.S. (Julia Shrubovych). Validation, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Formal analysis, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Investigation, J.S. (Julia Shrubovych) and J.S. (Jerzy Smykla). Resources, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Data curation, J.S. (Julia Shrubovych). Writing—original draft preparation, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Writing—review and editing, J.S. (Julia Shrubovych), J.S. (Jerzy Smykla), and E.C.B. Visualization, J.S. (Julia Shrubovych) and J.S. (Jerzy Smykla). Supervision, J.S. (Julia Shrubovych) and J.S. (Jerzy Smykla). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors are very grateful to Anatoly Babenko (Severtsov Institute of Ecology and Evolution) for Protura material from Yakutia (Siberia) and his valuable comments. We also thank the curators of the collections of Smithsonian Institution, Natural Museum of National History, Washington, DC, for lending us the type material of Nipponentomon macleani for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pass, G.; Szucsich, N. 100 years of research on the Protura: Many secrets still retained. Soil Org. 2011, 83, 309–334. [Google Scholar]
- Silvestri, F. Descrizioni di un nuovo genere di Insetti Apterigoti, rappresantante di un novo oridine. Boll. Lab. Zool. Gen. E Agrar. Della R Scuola Agric. Portici 1907, 1, 296–311. [Google Scholar]
- Carapelli, A.; Bu, Y.; Chen, W.-J.; Nardi, F.; Leo, C.; Frati, F.; Luan, Y.-X. Going deeper into high and low phylogenetic relationships of Protura. Genes 2019, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeptycki, A. Catalogue of the world Protura. Acta Zool. Cracoviensa 2007, 50B, 1–210. [Google Scholar]
- Galli, L.; Shrubovych, J.; Bu, Y.; Zinni, M. Genera of the Protura of the world: Diagnosis, distribution, and key. ZooKeys 2018, 772, 1–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeptycki, A. The taxonomy of Protura present status and future problems: Proceedings of the Xth international colloquium on Apterygota, České Budějovice 2000: Apterygota at the beginning of the third millennium. Pedobiologia 2002, 46, 209–214. [Google Scholar]
- Galli, L.; Capurro, M.; Colasanto, E.; Molyneux, T.; Murray, A.; Torti, C.; Zinni, M. A synopsis of the ecology of Protura (Arthropoda: Hexapoda). Rev. Suisse Zool. 2020, 126, 155–164. [Google Scholar]
- Nosek, J. Niches of Protura in biogeocenoses. Pedobiologia 1975, 15, 290–298. [Google Scholar]
- Szeptycki, A. The present knowledge of Protura. Fragm. Faun. 1997, 40, 307–311. [Google Scholar] [CrossRef] [Green Version]
- Tuxen, S.L. The first record of Canadian Protura. With systematic notes on Acerentulus. Entomol. Medd. 1955, 27, 113–128. [Google Scholar]
- Nosek, J. A new genus and six new species of Protura from Alaska (Protura: Acerentomidae, Eosentomidae). Entomol. Scand. 1977, 8, 271–284. [Google Scholar] [CrossRef]
- Nosek, J. A new genus and five species of Protura from Alaska. Entomol. Scand. 1980, 11, 265–273. [Google Scholar] [CrossRef]
- Nosek, J. Three new Protura from Alaska with key to known Alaskan species (Insecta: Protura). Entomol. Scand. 1981, 12, 158–162. [Google Scholar] [CrossRef]
- Bernard, E.C. Two new species of Protura (Insecta) from North America. Proc. Biol. Soc. Wash. 1985, 98, 72–80. [Google Scholar]
- Martynova, E.F. Acerella sharovi sp. n. (Protura, Acerentomidae) from Magadan district. Zool. Zhurnal 1977, 56, 164–167. [Google Scholar]
- Shrubovych, J.; Bernard, E.C. Two new species of Verrucoentomon (Protura: Acerentomidae, Nipponentominae) and a key to species. Ann. Entomol. Soc. Am. 2012, 105, 628–637. [Google Scholar] [CrossRef]
- Shrubovych, J.; Rusek, J.; Bernard, E.C. Redefinition and four new species of Yavanna Szeptycki and comparison with Nosekiella Rusek (Protura: Acerentomidae: Nipponentominae). Ann. Entomol. Soc. Am. 2012, 105, 3–19. [Google Scholar] [CrossRef]
- Shrubovych, J. Identification and character analysis of the Acerentomidae (Protura) of the northeastern Palearctic (Protura: Acerentomidae). Zootaxa 2014, 3755, 136–164. [Google Scholar] [CrossRef] [Green Version]
- Shrubovych, J.; Rusek, J.; Bernard, E.C. Revision of Vesiculentomon, Nosekientomon n. g. (Protura: Acerentomidae, Nipponentominae), and a key to genera of Nipponentominae. Ann. Entomol. Soc. Am. 2014, 107, 74–80. [Google Scholar] [CrossRef]
- Shrubovych, J.; Rusek, J.; Bernard, E.C. Revision of Nosekiella (Protura: Acerentomidae, Nipponentominae). Ann. Entomol. Soc. Am. 2014, 107, 721–727. [Google Scholar] [CrossRef]
- Shrubovych, J.; Rusek, J.; Smykla, J.; Bernard, E.C. Review of Alaskaentomon (Protura: Acerentomidae, Nipponentominae). Ann. Entomol. Soc. Am. 2014, 107, 728–733. [Google Scholar] [CrossRef]
- Shrubovych, J.; Rusek, J.; Smykla, J.; Bernard, E.C. Review of North American Verrucoentomon species (Protura: Acerentomidae, Nipponentominae), with a key to species of the related genus Imadateiella Rusek. Fla. Entomol. 2015, 98, 215–222. [Google Scholar] [CrossRef]
- Weber, N.A. A survey of the insects and related arthropods of Arctic Alaska. Part I. Trans. Am. Entomol. Soc. 1890- 1950, 76, 147–206. [Google Scholar]
- Yin, W.Y.; Xie, R.D. A new genus, two new species and two new records of Protura from the Northeast China. Contrib. Shanghai Inst. Entomol. 1993, 11, 65–73. [Google Scholar]
- Imadate, G. Occurrence of Nosekiella (Protura, Acerentomidae) in Japan. Annot. Zool. Jpn. 1981, 54, 142–146. [Google Scholar]
- Shrubovych, J.; Rusek, J. Sugaentulus andrzeji sp. nov. from Siberia, Russia (Protura: Acerentomidae: Acerentominae) and key to Acerentominae genera. Zootaxa 2010, 2720, 59–68. [Google Scholar] [CrossRef]
- Shrubovych, J. Nienna chukotka sp. nov. (Protura, Acerentomidae, Nipponentominae) from the Arctic region, with a key to species of the genus. ZooKeys 2019, 899, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.L.; Hacquebord, L.; Gregor, D.J.; Loeng, H. Physical/geographical characteristics of the Arctic. In AMAP Assessment Report: Arctic Pollution Issues. Chapter 2; Arctic Monitoring and Assessment Programme (AMAP): Oslo, Norway, 1998; pp. 9–24. [Google Scholar]
- Dunger, W.; Fiedler, H.J. Methoden der Bodenbiologie; Gustav Fischer: Jena, Germany, 1989. [Google Scholar]
- Szeptycki, A. New genera and species of Protura from the Altai Mts. Acta Zool. Cracoviensa 1988, 31, 297–362. [Google Scholar]
- Rusek, J.; Shrubovych, J.; Szeptycki, A. Head porotaxy and chaetotaxy of order Acerentomata (Protura). Zootaxa 2012, 3262, 54–61. [Google Scholar] [CrossRef]
- Batchelor, C.L.; Margold, M.; Krapp, M.; Murton, D.K.; Dalton, A.S.; Gibbard, P.L.; Stokes, C.R.; Murton, J.B.; Manica, A. The configuration of Northern Hemisphere ice sheets through the Quaternary. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Bu, Y.; Wu, D.H. Revision of Chinese Yamatentomon (Protura: Acerentomata: Acerentomidae), with description of one new species, redescription of Yamatentomon yamato, and key to world species. Fla. Entomol. 2012, 95, 839–847. [Google Scholar] [CrossRef]
- Szeptycki, A. Polish Protura. I. Genus Acerentomon Silvestri, 1907. Pol. Pismo Entomol. 1980, 50, 311–392. [Google Scholar]
- Shrubovych, J.; Bartel, D.; Szucsich, N.U.; Resch, M.C.; Pass, G. Morphological and genetic analysis of the Acerentomon doderoi group (Protura: Acerentomidae) with description of A. christiani sp. nov. PLoS ONE 2016, 11, e0148033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, R.T. Studies on the North American Protura 1: Catalogue and atlas of the Protura of North America; description of new species; key to the species of Eosentomon. Proc. Acad. Nat. Sci. Phila. 2007, 156, 97–116. [Google Scholar] [CrossRef]
- Shrubovych, J.; Schneider, C.; D’Haese, C.A. Revision of the genus Andinentulus (Protura: Acerentomidae: Berberentulinae), with a key to South and Central American Acerentomidae species. Ann. Entomol. Soc. Am. 2014, 107, 567–574. [Google Scholar] [CrossRef]
- Shrubovych, J.; Bernard, E.C. A key for the determination of European species of Eosentomon Berlese, 1909 (Protura, Eosentomata, Eosentomidae). ZooKeys 2018, 742, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bu, Y.; Potapov, M.B.; Yin, W.Y. Systematic and biogeographical study of Protura (Hexapoda) in Russian Far East: New data on high endemism of the group. ZooKeys 2014, 424, 19–57. [Google Scholar] [CrossRef] [Green Version]
- Shrubovych, J. Redescription of Verrucoentomon montanum new status (= Acerella montana Martynova) (Protura: Acerentomidae, Nipponentominae). Zootaxa 2011, 2743, 63–67. [Google Scholar] [CrossRef]
- Bluhm, S.L.; Potapov, A.M.; Shrubovych, J.; Ammerschubert, S.; Polle, A.; Scheu, S. Protura are unique: First evidence of specialized feeding on ectomycorrhizal fungi in soil invertebrates. BMC Ecol. 2019, 19, 10. [Google Scholar] [CrossRef] [Green Version]
- Babenko, A. Personal Communication; Severtsov Institute of Ecology and Evolution: Moscow, Russia, 2019. [Google Scholar]
- Tuxen, S.L. Protura (Insecta) and Brazil during 400 million years of continental drift. Stud. Neotrop. Fauna Environ. 1978, 13, 23–50. [Google Scholar] [CrossRef]
- Sikes, D.S. Diplura and Protura of Canada. ZooKeys 2019, 819, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Sikes, D.S.; Allen, R.T. First Alaskan records and a significant northern range extension for two species of Diplura (Diplura, Campodeidae). ZooKeys 2016, 563, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Map showing all known sites of Protura occurrence within the Arctic regions. The dark grey irregular line shows the largest ice-sheet extent during the Pleistocene Last Glacial Cycle (redrawn after Reference [32]).
Figure 1.
Map showing all known sites of Protura occurrence within the Arctic regions. The dark grey irregular line shows the largest ice-sheet extent during the Pleistocene Last Glacial Cycle (redrawn after Reference [32]).

Figure 2.
Mastodonentomon macleani (Nosek, 1977). (A) Pseudoculus with setae l3 and sd4. (B) Cephalic seta sd 5. (C) Hind part of head. (D) Foretarsal seta β1. (E) Foretarsal seta γ4. (F) Pronotum and mesonotum, right side. (G) Metanotum, right side. (H) Anterior part of the prosternum. (I) Anterior part of the mesosternum. (J) Anterior part of the metasternum. (K) Sternite I. (L) Sternite VII. (M) Tergites I and II, right side. (N) Tergite VII. (O) Hind margin of tergite IX. (P) Hind margin of tergite X. Arrows indicate pores (sl = sublateral, psm = posterosubmedial, psl = posterosublateral, sc = sternal central, spm = sternal posteromedial, spsm = sternal posterosubmedial). Figures A–E, J: paratype. Figures F–I, K–P: holotype. Scale bars = 20 µm.
Figure 2.
Mastodonentomon macleani (Nosek, 1977). (A) Pseudoculus with setae l3 and sd4. (B) Cephalic seta sd 5. (C) Hind part of head. (D) Foretarsal seta β1. (E) Foretarsal seta γ4. (F) Pronotum and mesonotum, right side. (G) Metanotum, right side. (H) Anterior part of the prosternum. (I) Anterior part of the mesosternum. (J) Anterior part of the metasternum. (K) Sternite I. (L) Sternite VII. (M) Tergites I and II, right side. (N) Tergite VII. (O) Hind margin of tergite IX. (P) Hind margin of tergite X. Arrows indicate pores (sl = sublateral, psm = posterosubmedial, psl = posterosublateral, sc = sternal central, spm = sternal posteromedial, spsm = sternal posterosubmedial). Figures A–E, J: paratype. Figures F–I, K–P: holotype. Scale bars = 20 µm.


{kind=link}
{kind=link}
Table 1.
A list of all records of Protura from the Arctic regions with information on their taxonomic position, and geographical and environmental distribution. – * type locality.
Table 1.
A list of all records of Protura from the Arctic regions with information on their taxonomic position, and geographical and environmental distribution. – * type locality.
| Species and Their Taxonomic Position | Records from the Arctic | Environmental Settings | Distribution | References |
|---|---|---|---|---|
| Order Eosentomata | ||||
| Family Eosentomidae Berlese, 1909 | ||||
| Genus Eosentomon Berlese, 1908 | ||||
| Eosentomon adakense Bernard, 1985 | Kuluk Bay *, Adak Island, Aleutian Islands (Bering Sea), Alaska, USA (51°50′ N, 176°40′ W). | Soil, beach terrace with dune grass (Elymus mollis). | Alaska (type area only). | [14] |
| Eosentomon alaskaense Nosek, 1977 | Finger Mountain (671 m elev.), Yukon-Koyukuk, Alaska, USA (66°21′30″ N, 150°27′30″ W). Mastodon Dome * at Eagle Creek, Crazy Mountains, Yukon-Koyukuk, Alaska, USA (65°26′ N, 145°21′ W). Nunivak Island (Bering Sea), Alaska, USA (60°06′ N, 166°30′ W). | * 1200 m elev., on exposed cliff in Dryas sp. | Alaska. | [11] |
| Eosentomon aquilinum Nosek, 1980 | Denali National Park and Preserve * (as Mt. McKinley N.P.) between Teklanika River and Sanctuary River, Alaska, USA (63°40′ N, 149°30′ W). | Soil, relatively dry slope with dead aspen. | Alaska (type area only). | [12] |
| Eosentomon copelandi Nosek, 1980 | Denali National Park and Preserve * (as Mt. McKinley N.P.), Eielson, Alaska, USA (63°25′ N, 150°20′ W). | Soil, alpine tundra below talus, grass tussocks and moss. | Alaska (type area only). | [12] |
| Eosentomon sp. (“larva 2”, indeterminable to species) | Reindeer Station, Northwest Territories, northern Canada (68°42′ N, 134°08′ W). | Moist soil with lichens. | – | [10] |
| Order Acerentomata | ||||
| Family Acerentomidae Silvestri, 1907 | ||||
| Acerentulus sp. (undetermined species). | Umiat Mt., Alaska, USA (69°23′30″ N, 152°01′18″ W). | Humus, below the summit, above the Colville River at elevation 900 feet (ca.275 m), the warmest exposure on the south side of the bluff. | – | [23] |
| Acerentulus sp. (undetermined species). | Anaktuvuk Pass, Alaska, USA (68°10′ N, 151°45′ W). | Humus, about lichens, mosses, and higher plant roots, at elevation 3950 feet (ca.1200 m) on fully exposed jagged rocks facing the pass. | – | [23] |
| Subfamily Acerentominae Silvestri, 1907 | ||||
| Genus Fjellbergella Nosek, 1978 | ||||
| Fjellbergella tuxeni Nosek, 1980 | Chena Ridge, Fairbanks, Alaska, USA (64°50′ N, 147°57′ W). Brushkana River E of Cantwell *, Denali, Alaska, USA (63°20′ N, 148°10′ W). Denali National Park and Preserve (as Mt. McKinley N.P.), Denali, Alaska, USA (63°00′ N, 151°00′ W). | *Soil, tundra with moss, lichens, grass, Ledum sp., Vaccinium sp., thick turf. | Alaska. | [12] |
| Genus Orinentomon Yin, Xie, 1993 | ||||
| Orinentomon greenbergi (Nosek, 1980) syn. Yamatentomon greenbergi Nosek, 1980 | Denali National Park and Preserve * (as Mt. McKinley N.P.), Eielson, Alaska, USA (63°25′ N, 150°20′ W). | Soil; moss, grass, Salix in alpine tundra talus. | Alaska (type area only). | [12] |
| Genus Sugaentulus Imadaté, 1978 | ||||
| Sugaentulus hoogstraali (Nosek, 1980) syn. Nosekiella hoogstraali Nosek, 1980 | Smith Lake *, Fairbanks, Alaska, USA (64°52′ N, 147°52′ W). Denali National Park and Preserve (as Mt. McKinley N.P.), Eielson, Alaska, USA (63°25′ N, 150°20′ W). | * Soil and litter, deep rotten layer in squirrel mound of white spruce cones. Soil, moss, grass, Salix in alpine tundra below talus. | Alaska. | [12,20] |
| Sugaentulus andrzeji Shrubovych & Rusek, 2010 | Turukhansk area *, Evenkia, Krasnoyarskiy Kray, Siberia, Russia (65°48′ N, 88°00′ E). | Soil, litter, and moss in birch, mixed and montane coniferous forests with dense herbaceous cover. | Northern* and Southern Siberia. | [18,26] |
| Genus Tuxenentulus Imadaté, 1973 | ||||
| Tuxenentulus boedvarssoni Nosek, 1981 | Chena Ridge *, Fairbanks, Alaska, USA (64°50′ N, 147°57′ W). | Humus and dry litter in a mixed alder, aspen, birch forest. | Alaska (type area only). | [13] |
| Genus Yamatentomon Imadaté, 1964 | ||||
| Yamatentomon yamato (Imadaté & Yosii, 1956) syn. Acerentulus yamato Imadaté & Yosii, 1956 Acerentomon yamato, Acerella yamato, Yamatentomon tokapchupi Imadaté, 1964 | Lena Delta Nature Reserve, Tyallakh, Sakha (Yakutia), Siberia, Russia (72°19′36″ N, 128°07′27″ E). | Soil and moss in sedge/moss tundra. | Japan *, Korea, Northern China, Russian Far East, and Northern Siberia. | [3,17,31] Current study. |
| Subfamily Nipponentominae Yin, 1983 | ||||
| Genus Alaskaentomon Nosek, 1977 | ||||
| Alaskaentomon condei Nosek, 1981 | Chena Ridge *, Fairbanks, Alaska, USA (64°50′ N, 147°57′ W). | Dry alder litter in a mixed alder-aspen-birch forest. | Alaska (type area only). | [13,21] |
| Alaskaentomon fjellbergi Nosek, 1977 | Sagwon Upland *, CRREL 12, Alaska, USA (69°30′ N, 148°30′ W). | Dry exposed hill with Dryas sp. and Carex rupestris. | Alaska (type area only). | [11,21] |
| Genus Imadateiella Rusek, 1974 | ||||
| Imadateiella mixta (Nosek, 1981) syn. Verrucoentomon mixtum Nosek, 1981 | Chena Ridge *, Fairbanks, Alaska, USA (64°50′ N, 147°57′ W). | Litter in shrubs with Betula sp. and Equisetum sp., N slope, rather moist. | Alaska (type area only). | [13,22] |
| Imadateiella sharovi (Martynova, 1977) syn. Acerella sharovi Martynova, 1977 | Turukhansk area, Evenkia, Krasnoyarskiy Kray, Siberia, Russia (65°48′ N, 88°00′ E). Aborigen Station, Kolyma River, Magadan District, Russia (61°59′ N, 149°20′ E). Snow Valley *, Magadan area, Magadan District, Russia (59°40′ N, 150°25′ E). | Soil, litter, and turf in dry localities, under lichens and shrubs. | Northern Siberia and Russian Far East. | [15,18] |
| Genus Nienna Szeptycki, 1988 | ||||
| Nienna chukotka Shrubovych, 2019 | Akanotenmeem hill *, near Apapelgino village, Chaunskiy district, Chukotka, Russia (69°48′40″ N, 170°35′51″ E). | Soil in dry locality with Dryas sp., 20 m elev. | Northern Siberia (type area only). | [27] |
| Genus Mastodonentomon n. gen. | ||||
| Mastodonentomon macleani (Nosek, 1977) syn. Nipponentomon macleani Nosek, 1977 | Mastodon Dome * at Eagle Creek, Crazy Mountains, Yukon-Koyukuk, Alaska, USA (65°26′ N, 145°21′ W). | 1200 m elev., on exposed cliff in Dryas sp. | Alaska (type area only). | [11] |
| Genus Vesiculentomon Rusek, 1974 | ||||
| Vesiculentomon condei (Tuxen, 1955) syn. Acerentulus condei Tuxen, 1955; Acerella condei; Nosekiella condei. | Richardson Mountains *, Yukon, northern Canada (68°24′ N, 135°37′ W). | 600 m elev., dry locality with Dryas sp. | Northern Canada (type area only). | [10,20] |
| Genus Nosekientomon Shrubovych, Rusek, Bernard 2014 | ||||
| Nosekientomon ruseki (Nosek, 1977) syn. Vesiculentomon ruseki Nosek, 1977 | Mastodon Dome * at Eagle Creek, Crazy Mountains, Yukon-Koyukuk, Alaska, USA (65°26′ N, 145°21′ W). | 1200 m elev., on exposed cliff in Dryas sp. | Alaska (type area only). | [11,19] |
| Genus Verrucoentomon Rusek, 1974 | ||||
| Verrucoentomon canadense (Tuxen, 1955) syn. Acerentulus canadensis Tuxen, 1955, Acerella canadensis; Nosekiella canadensis; Verrucoentomon canadensis. | Reindeer Station, Northwest Territories, northern Canada (68°42′ N, 134°08′ W). Richardson Mountains *, Yukon, northern Canada, (68°24′ N, 135°37′ W). | 130 m elev., moist localities with moss, Cladinia, Empetrum, cowberries and Betula nana. *600 m elev., dry locality with Dryas sp. | Northern Canada. | [10,22] |
| Verrucoentomon imadatei Nosek, 1977 syn. Acerentulus imadatei Nosek, 1977 | Meade River at Atqasuk *, North Slope, Alaska, USA (70°29′ N, 157°25′ W). | Litter and humus in Salix-shrub on sandy river slope. | Alaska (type area only). | [11,22] |
| Verrucoentomon anatoli Shrubovych & Bernard, 2012 | Turukhansk area *, Evenkia, Krasnoyarskiy Kray, Siberia, Russia (65°48′ N, 88°00′ E). Mirnoye area, Turukhanskiy District, Krasnoyarskiy Kray, Siberia, Russia (62°35′ N, 89°03′ E). Podkamennaya Tunguska River (bank), Borskiy District, Krasnoyarskiy Kray, Siberia, Russia (61°70′ N, 90°63′ E). | Soil, moss, and litter in mixed taiga forests. | Northern and southern Siberia. | [16,18] |
| Genus Yavanna Szeptycki, 1988 | ||||
| Yavanna babenkoi Shrubovych, Rusek & Bernard 2012 | Turukhansk area *, Evenkia, Krasnoyarskiy Kray, Siberia, Russia (65°48′ N, 88°00′ E). Mirnoye area, Turukhanskiy District, Krasnoyarskiy Kray, Siberia, Russia (62°35′ N, 89°03′ E). | Soil, litter, and moss in mixed and taiga forest. | Northern and southern Siberia. | [17,18] |
| Yavanna behanae (Nosek, 1977) syn Nosekiella behanae Nosek, 1977 | Gobbler Knobb * (920 m elev.), Yukon-Koyukuk, Alaska, USA (67°28′30″ N, 150°11′30″ W). | Moist lichen slope in paper birch stand. | Alaska (type area only). | [11] |
Table 2.
Body chaetotaxy of Mastodonentomon macleani (Nosek, 1977).
| Dorsal | Ventral | |||
|---|---|---|---|---|
| Segment | Formula | Setal Composition | Formula | Setal Composition |
| Th. I | 4 | 1, 2 | 4 + 4 6 | A1, 2, M1, 2 P1, 2, 3 |
| Th. II | 8 + 2 16 | A1, 2, 3, 4, M P1, 1a, 2, 2a, 3, 3a, 4, 5 | 5 + 2 4 | Ac, 2, 3, M P1, 3 |
| Th. III | 8 + 6 16 | A1, 2, 3, 4, M1, 2, 3 P1, 1a, 2, 2a, 3, 3a, 4, 5 | 9 + 2 4 | Ac, 1, 2, 3, 4, M P1, 3 |
| Abd. I | 8 12 | A1, 1’, 2, 5 P1, 1a, 2, 2a, 3, 4 | 5 4 | Ac, 2, 3 P1, 1a |
| Abd. II-III | 12 16 | A1, 1’, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 | 5 5 | Ac, 1, 2 Pc, 1a, 2 |
| Abd. IV-VI | 10 16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 | 5 8 | Ac, 2, 3 P1, 1a, 2, 3 |
| Abd. VII | 10 16 | A1, 2, 3, 4, 5 P1, 1a, 2, 2a, 3, 4, 4a, 5 | 3 9 | Ac, 2 Pc, 1, 1a, 2, 3 |
| Abd. VIII | 6 15 | A1, 4, 5 Pc, 1, 1a, 2, 2a, 3, 3a, 5 | 4 2 | 1, 2 P1a |
| Abd. IX | 12 | 1, 1a, 2, 2a, 3, 4 | 4 | |
| Abd. X | 10 | 1, 2, 2a, 3, 4 | 4 | |
| Abd. XI | 6 | 1, 3, 4 | 6 | |
| Abd. XII | 9 | 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shrubovych, J.; Smykla, J.; Bernard, E.C. Protura in Arctic Regions, with Description of Mastodonentomon n. gen. (Acerentomidae, Nipponentominae) and a Key to Known Arctic Taxa. Insects 2020, 11, 173. https://doi.org/10.3390/insects11030173
AMA Style
Shrubovych J, Smykla J, Bernard EC. Protura in Arctic Regions, with Description of Mastodonentomon n. gen. (Acerentomidae, Nipponentominae) and a Key to Known Arctic Taxa. Insects. 2020; 11(3):173. https://doi.org/10.3390/insects11030173
Chicago/Turabian StyleShrubovych, Julia, Jerzy Smykla, and Ernest C. Bernard. 2020. "Protura in Arctic Regions, with Description of Mastodonentomon n. gen. (Acerentomidae, Nipponentominae) and a Key to Known Arctic Taxa" Insects 11, no. 3: 173. https://doi.org/10.3390/insects11030173
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.