Transcriptome Analyses of Diaphorina citri Midgut Responses to Candidatus Liberibacter Asiaticus Infection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing, Tissue Collection and Total RNA Extraction
2.2. Library Preparation and Illumina Sequencing
2.3. Read Mapping and Identification of DEGs
2.4. Gene Ontology (GO) and KEGG Enrichment Analysis of DEGs
2.5. Reverse Transcription Quantitative PCR (RT-qPCR) Analysis
3. Results
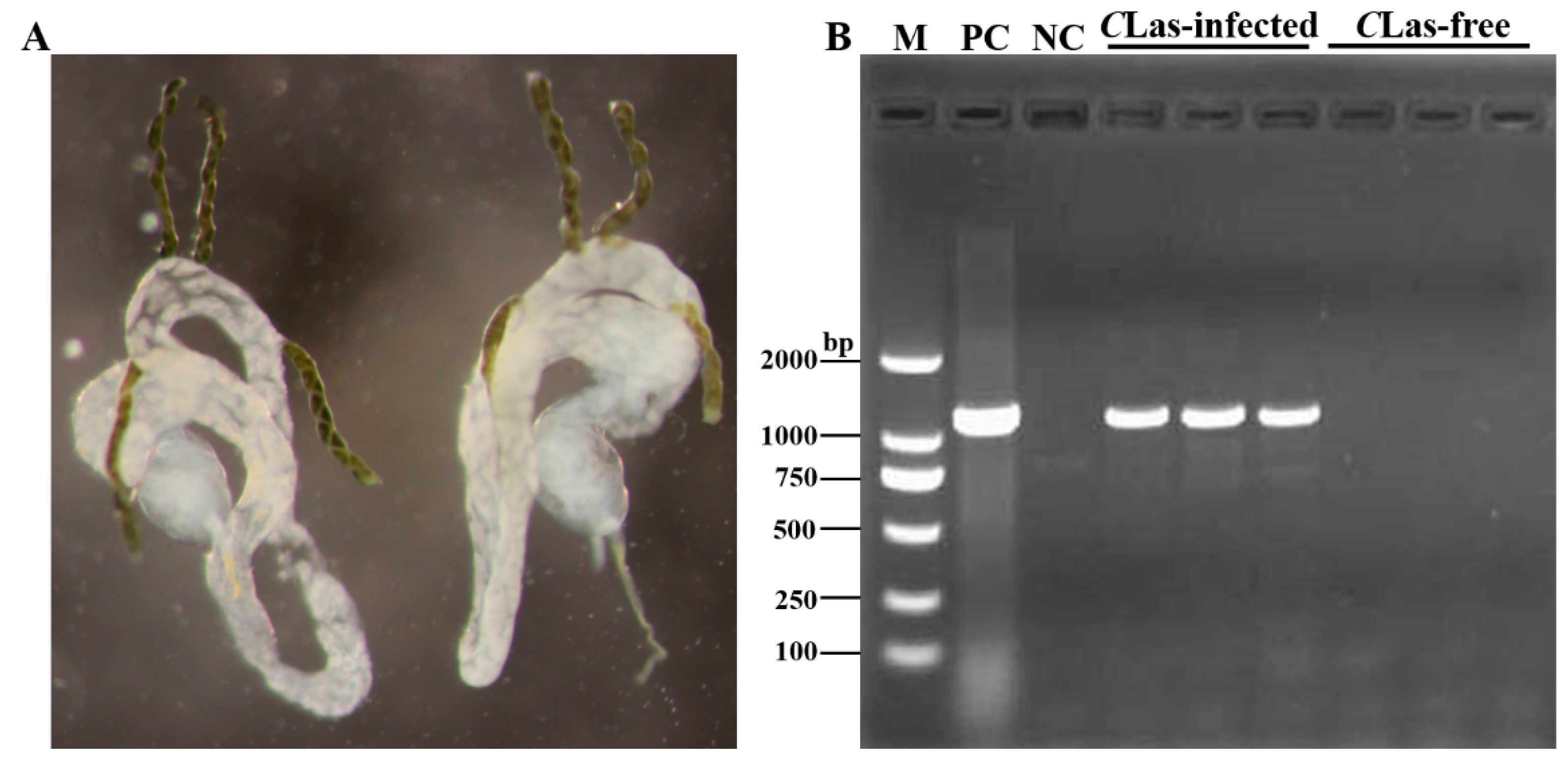
3.1. Detection of CLas Infection in the Midgut of D. citri
3.2. Illumina Sequencing and Read Assembly
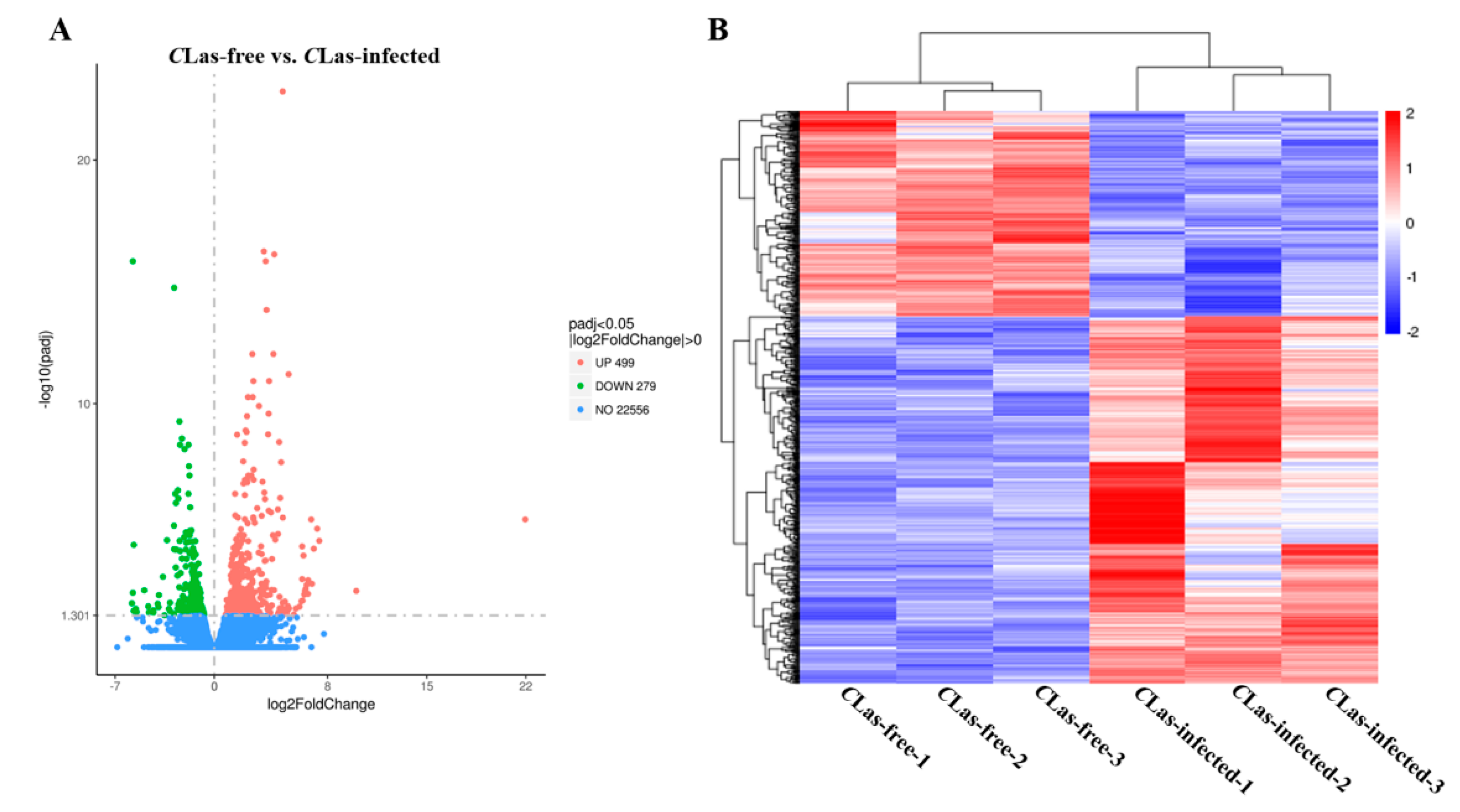
3.3. Identification of DEGs in Response to CLas Infection
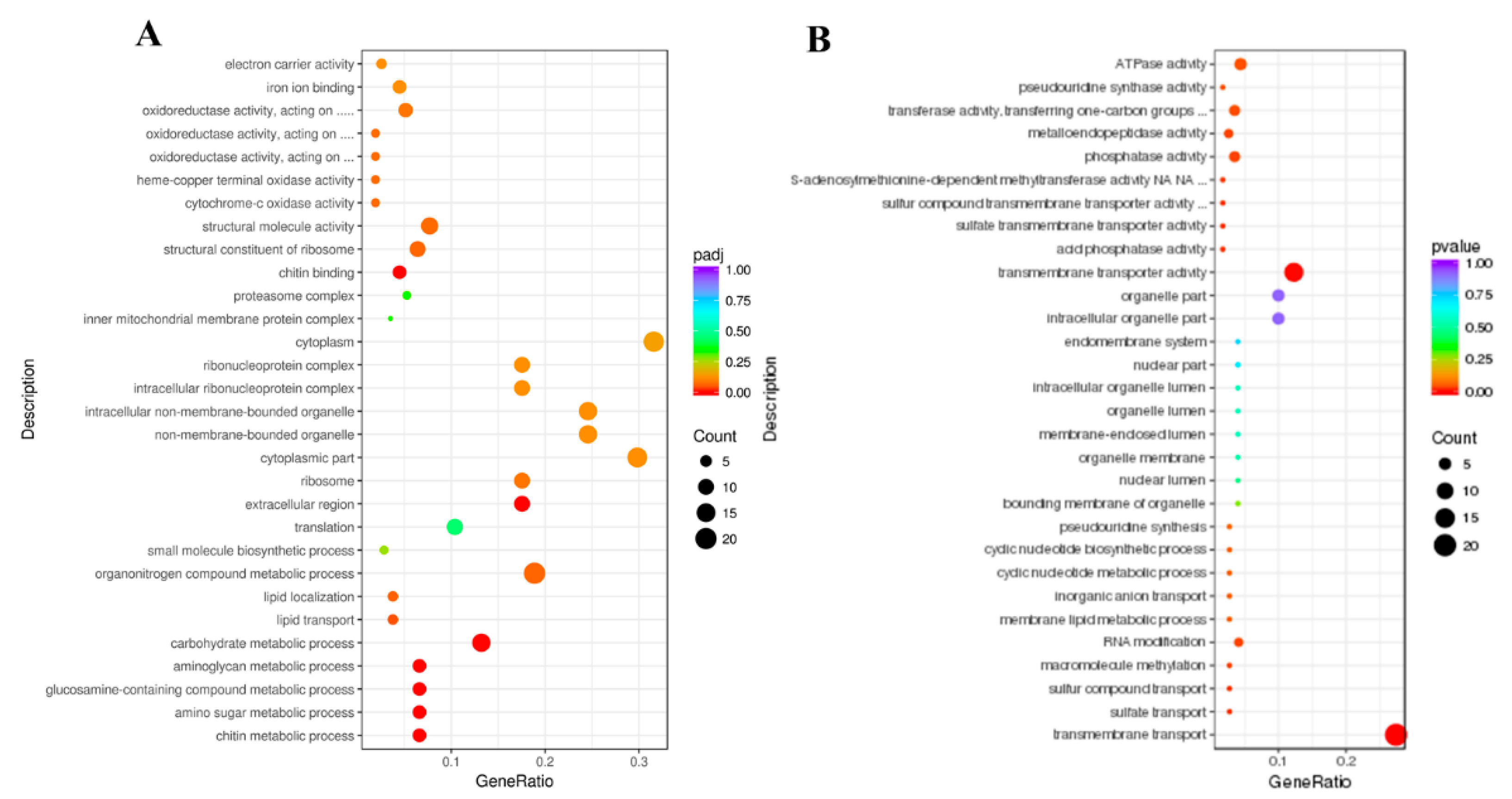
3.4. GO and KEGG Enrichment Analysis
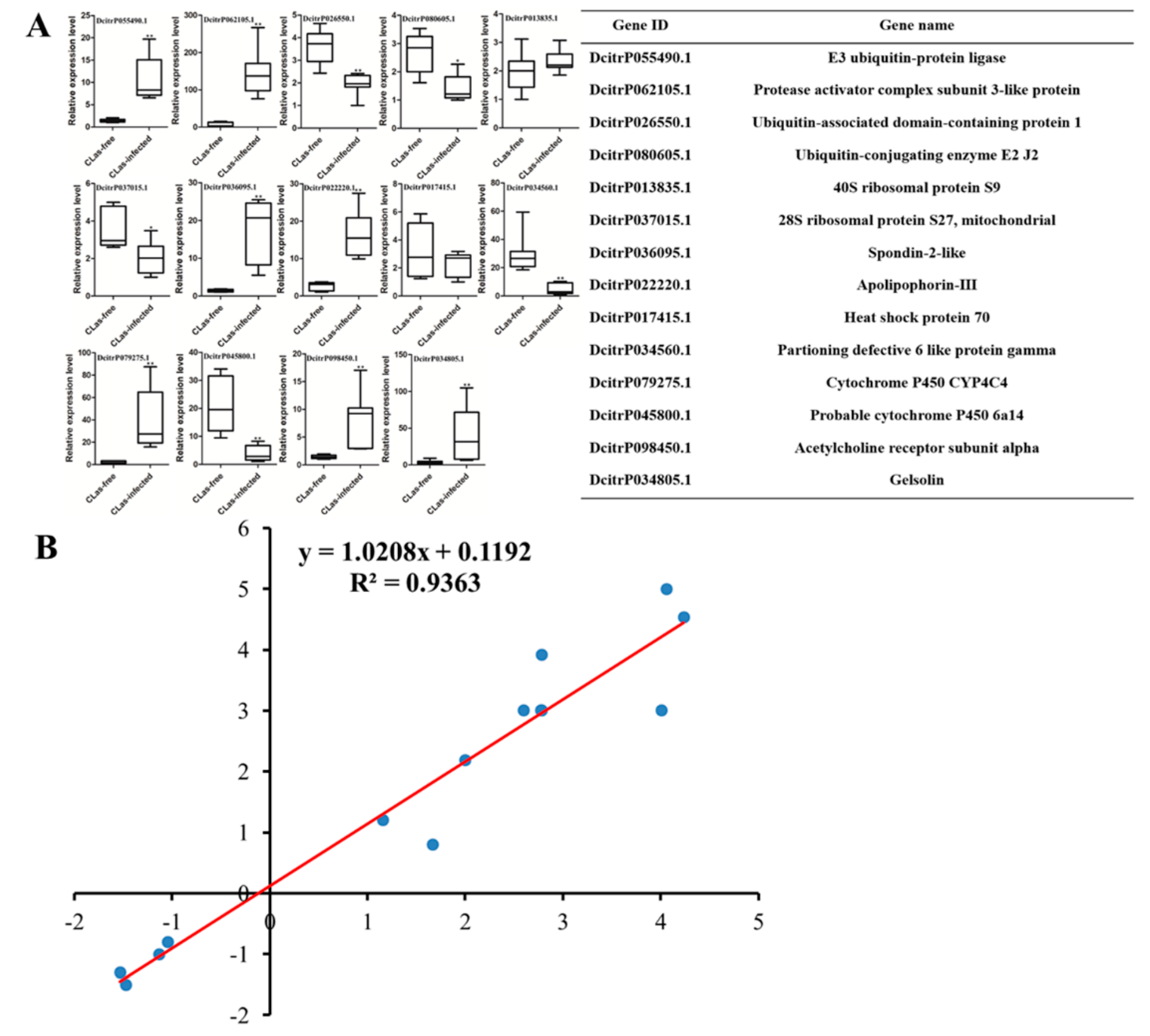
3.5. Validation of DEGs at the Transcriptional Level
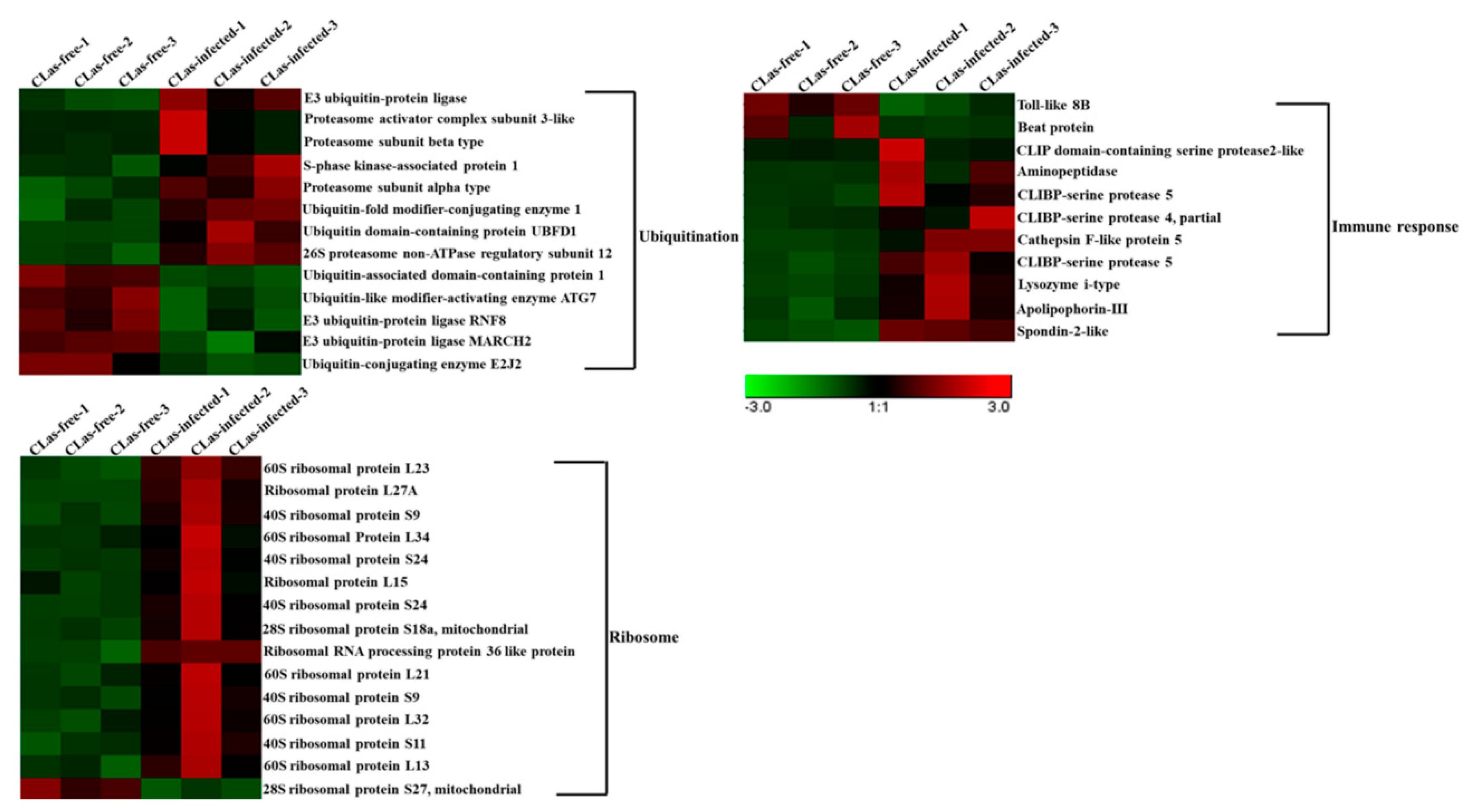
3.6. Analysis of DEGs Associated with Ubiquitination, the Immune Response and the Ribosome
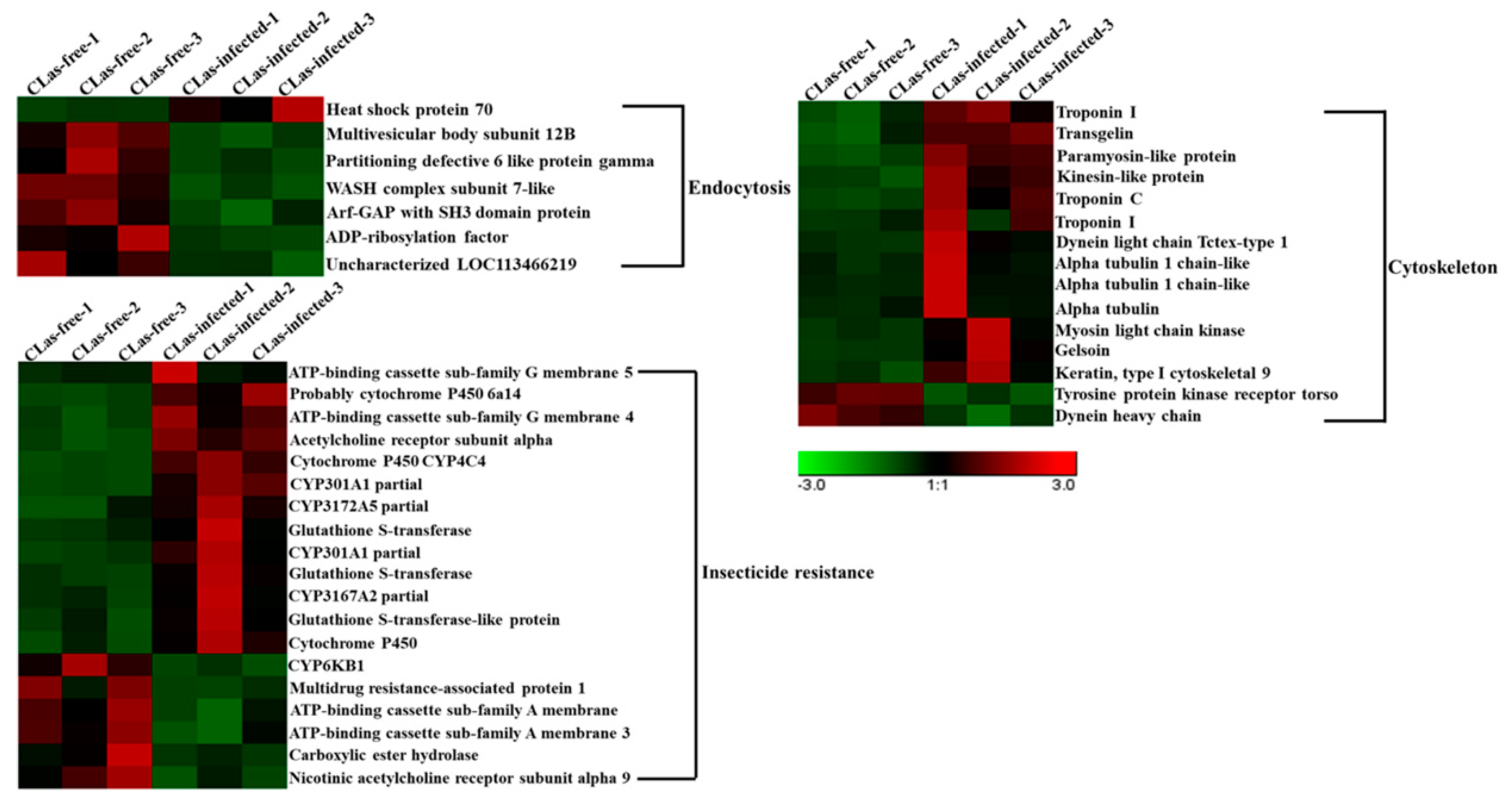
3.7. Analysis of DEGs Involved in Endocytosis, the Cytoskeleton and Insecticide Resistance
4. Discussion
4.1. Ubiquitination, the Immune Response and Ribosomes May Play Important Roles in the Midgut Response to CLas Infection
4.2. Endocytosis, the Cytoskeleton and Insecticide Resistance May Play Crucial Roles in the Midgut Response to CLas Infection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bove, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J.J. The Candidatus Liberibacter-host interface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef] [PubMed]
- Slisz, A.M.; Breksa, A.P.; Mishchuk, D.O.; McCollum, G.; Slupsky, C.M. Metabolomic analysis of citrus infection by ‘Candidatus Liberibacter’ reveals insights into pathogenicity. J. Proteome Res. 2012, 22, 4223–4230. [Google Scholar] [CrossRef]
- Hu, J.H.; Jiang, J.X.; Wang, N. Control of citrus Huanglongbing (HLB) via trunk injection of plant activators and antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, J.A.; Kostyk, B.C.; Stansly, P.A. Insecticidal suppression of Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) vector of huanglongbing pathogens. PLoS ONE 2014, 9, e112331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolora, L.D.; Powell, C.M.; Hunter, W.; Bextine, B.; Lauzon, C.R. Internal extracellular bacteria of Diaphorina citri Kuwayama (Hemiptera: Psyllidae), the Asian citrus psyllid. Curr. Microbiol. 2015, 70, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Boina, D.R.; Bloomquist, J.R. Chemical control of the Asian citrus psyllid and of huanglongbing disease in citrus. Pest Manag. Sci. 2015, 71, 808–823. [Google Scholar] [CrossRef] [PubMed]
- Boina, D.R.; Onagbola, E.O.; Salyani, M.; Stelinski, L.L. Antifeedant and sublethal effects of imidacloprid on Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2010, 65, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.K.; Hu, R.F.; Pray, C.; Qiao, F.B.; Rozelle, S. Biotechnology as an alternative to chemical pesticides: A case study of Bt cotton in China. Agric. Econ. 2015, 29, 55–67. [Google Scholar] [CrossRef]
- Coutinho-Abreu, I.V.; Forster, L.; Guda, T.; Ray, A. Odorants for surveillance and control of the Asian citrus psyllid (Diaphorina citri). PLoS ONE 2014, 9, e109236. [Google Scholar] [CrossRef]
- Yu, X.D.; Gowda, S.; Killiny, N. Double-stranded RNA delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid. Diaphorina citri. Pest Manag. Sci. 2017, 73, 1846–1853. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.J.; Zhou, C.H.; Yu, H.Z.; Huang, Y.L.; Liu, Y.X.; Xie, Y.X.; Wang, J.; Hu, W.; Huang, A.J.; Su, H.N.; et al. Potential roles of insect Tropomyosin1-X1 isoform in the process of Candidatus Liberibacter asiaticus infection of Diaphorina citri. J. Insect Physiol. 2019, 114, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Ohnishi, J.; Ito, T.; Tomimura, K.; Miyata, S.; Iwanami, T.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2010, 155, 29–36. [Google Scholar] [CrossRef]
- Coletta-Filho, H.D.; Daugherty, M.P.; Ferreira, C.; Lopes, J.R. Temporal progression of ‘Candidatus Liberibacter asiaticus’ infection in citrus and acquisition efficiency by Diaphorina citri. Phytopathology 2014, 104, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Ammar, E.D.; Shatters, R.G., Jr.; Hall, D.G. Localization of Candidatus Liberibacter asiaticus, associated with citrus huanglongbing disease, in its psyllid vector using fluorescence in situ hybridization. J. Phytopathol. 2011, 159, 726–734. [Google Scholar] [CrossRef]
- Gakhar, S.K.; Shandilya, H.K. Midgut specific immune response of vector mosquito Anopheles stephensi to malaria parasite Plasmodium. Indian J. Exp. Biol. 2011, 39, 287–290. [Google Scholar]
- Vincendeau, P.; Gobert, A.P.; Daulouede, S.; Moynet, D.; Mossalayi, M.D. Arginases in parasitic diseases. Trends Parasitol. 2003, 19, 9–12. [Google Scholar] [CrossRef]
- Rider, M.A.; Zou, J.; Vanlandingham, D.; Nuckols, J.T.; Higgs, S.; Zhang, Q.; Lacey, M.; Kim, J.; Wang, G.; Hong, Y.S. Quantitative proteomic analysis of the Anopheles gambiae (Diptera: Culicidae) midgut infected with o’nyong-nyong virus. J. Med. Entomol. 2013, 50, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Yu, H.Z.; Geng, L.; Xu, J.P.; Yu, D.; Zhang, S.Z.; Ma, Y.; Fei, D.Q. Comparative transcriptome analysis of Bombyx mori (Lepidoptera) larval midgut response to BmNPV in susceptible and near-isogenic resistant strains. PLoS ONE 2016, 11, e0155341. [Google Scholar] [CrossRef]
- Bao, Y.Y.; Wang, Y.; Wu, W.J.; Zhao, D.; Xue, J.; Zhang, B.Q.; Shen, Z.C.; Zhang, C.X. De novo intestine-specific transcriptome of the brown planthopper Nilaparvata lugens revealed potential functions in digestion, detoxification and immune response. Genomics 2012, 99, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Kruse, A.; Fattah-Hosseini, S.; Saha, S.; Johnson, R.; Warwick, E.; Sturgeon, K.; Mueller, L.; MacCoss, M.J.; Shatters, R.G.J.; Cilia Heck, M. Combining ‘omics and microscopy to visualize interactions between the Asian citrus psyllid vector and the Huanglongbing pathogen Candidatus Liberibacter asiaticus in the insect gut. PLoS ONE 2017, 12, e0179531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.J.; Killiny, N. Huanglongbing pathogen Candidatus Liberibacter asiaticus exploits the energy metabolism and host defence responses of its vector Diaphorina citri. Physiol. Entomol. 2017, 42, 319–335. [Google Scholar] [CrossRef]
- Kruse, A.; Ramsey, J.S.; Johnson, R.; Hall, D.G.; MacCoss, M.J.; Heck, M. Candidatus Liberibacter asiaticus’ minimally alters expression of immunity and metabolism proteins in the hemolymph of Diaphorina citri, the insect vector of Huanglongbing. J. Proteome Res. 2018, 17, 2995–3011. [Google Scholar] [CrossRef] [PubMed]
- Shokrollah, H.; Abdullah, T.L.; Sijam, K.; Abdullah, S.N.A. Ultrastructure of Candidatus Liberibacter asiaticus and its damage in huanglongbing (HLB) infected citrus. Afr. J. Biotechnol. 2010, 9, 5897–5901. [Google Scholar]
- Saha, S.; Hosmani, P.S.; Villalobos-Ayala, K.; Miller, S.; Shippy, T.; Flores, M.; Rosendale, A.; Cordola, C.; Bell, T.; Mann, H.; et al. Improved annotation of the insect vector of citrus greening disease: Biocuration by a diverse genomics community. Database 2017, 2017, bax032. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHatL discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwiqht, S.S.; Eppiq, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Zheng, Q.; Wang, X.J. GOEAST: A web-based software toolkit for Gene Ontology enrichment analysis. Nucleic Acids Res. 2008, 36, 358–363. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.C.; Wang, L.G.; Han, Y.Y.; He, Q.Y. Clusterprofiler: An R Package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Cheraghi, E.; Mehranjani, M.S.; Shariatzadeh, S.M.A.; Esfahani, M.H.N.; Alani, B. N-acetylchsteine compared to metformin, improves the expression profiles of growth differentiation factor-9 and receptor tyrosine kinase c-kit in the oocytes of patients with polycystic ovarian syndrome. Int. J. Fertil. Steril. 2018, 11, 270–278. [Google Scholar] [PubMed]
- Camon, E.; Magrane, M.; Barrell, D.; Lee, V.; Dimmer, E.; Maslen, J.; Binns, D.; Harte, N.; Lopez, R.; Apweiler, R. The gene ontology annotation (GOA) database: Sharing knowledge in uniprot with gene ontology. Nucleic Acids Res. 2004, 32, D262–D266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. The KEGG databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xu, M.R.; Bao, M.L.; Wu, F.N.; Chen, J.C.; Deng, X.L. Unusual five copies and dual Forms of nrdB in “Candidatus Liberibacter asiaticus”: Biological implications and PCR detection application. Sci. Rep. 2016, 6, 39020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arp, A.P.; Hunter, W.B.; Pelz-Stelinski, K.S. Annotation of the Asian citrus psyllid genome reveals a reduced innate immune system. Front. Physiol. 2016, 7, 570. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 48, 1–5. [Google Scholar] [CrossRef]
- Rajkumar, A.P.; Qvist, P.; Lazarus, R.; Lescai, F.; Ju, J.; Nyegaard, M.; Mors, O.; Børglum, A.D.; Li, Q.B.; Christensen, J.H. Experimental validation of methods for differential gene expression analysis and sample pooling in RNA-seq. BMC Genom. 2015, 16, 548. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.H.; Xing, L.S.; Lin, Z.; Saha, T.T.; Wang, C.; Jiang, H.B.; Zou, Z. High throughput profiling of the cotton bollworm Helicoverpa armigera immunotranscriptome during the fungal and bacterial infections. BMC Genom. 2015, 16, 321. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Li, X.; Yang, X.; Yu, X.; Wang, J.H.; Liu, F.S.; Huang, D.W. Transcriptional response of Musca domestica larvae to bacterial infection. PLoS ONE 2014, 9, e104867. [Google Scholar] [CrossRef] [Green Version]
- Koegl, M.; Hoppe, T.; Schlenker, S.; Ulrich, H.D.; Mayer, T.U.; Jentsch, S. A novel ubiquitination factor, E4, is involved in multiubiquitin chain assembly. Cell 1999, 96, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Dupont, N.; Temime-Smaali, N.; Lafont, F. How ubiquitination and autophagy participate in the regulation of the cell response to bacterial infection. Biol. Cell 2010, 102, 621–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.D.; Yan, J.; Niu, H.; Huang, R.; Wu, S.Y. Autophagy and ubiquitination in Salmonella infection and the related inflammatory responses. Front. Cell Infect Microbiol. 2018, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Zhao, H.; Chen, Y.Y.; Luo, H.Y.; Yang, P.L.; Yao, B. A zebrafish (Danio rerio) bloodthirsty member 20 with E3 ubiquitin ligase activity involved in immune response against bacterial infection. Biochem. Biophys. Res. Commun. 2015, 457, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Grice, G.L.; Nathan, J.A. The recognition of ubiquitinated proteins by the proteasome. Cell Mol. Life Sci. 2016, 73, 3497–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacson, M.K.; Ploegh, H.L. Ubiquitination, ubiquitin-like modifiers, and deubiquitination in viral infection. Cell Host Microbe 2009, 5, 559–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.Z.; Wang, X.Y.; Xu, J.P.; Ma, Y.; Zhang, S.Z.; Yu, D.; Fei, D.Q.A. Muhammad, iTRAQ-based quantitative proteomics analysis of molecular mechanisms associated with Bombyx mori (Lepodoptera) larval midgut response to BmNPV in susceptible and near-isogenic strains. J. Proteom. 2017, 8, 35–50. [Google Scholar] [CrossRef]
- Wang, N.; Trivedi, P. Citrus huanglongbing: A newly relevant disease presents unprecedented challenges. Phytopathology 2013, 103, 652–665. [Google Scholar] [CrossRef] [Green Version]
- Crava, C.M.; Jacubowska, A.K.; Escriche, B.; Herrero, S.; Bel, Y. Dissimilar regulation of antimicrobial proteins in the midgut of Spodoptera exigua larvae challenged with Bacillus thuringiensis toxins or baculovirus. PLoS ONE 2015, 10, e0125991. [Google Scholar] [CrossRef]
- Chu, Y.; Liu, Y.; Shen, D.X.; Hong, F.; Wang, G.R.; An, C.J. Serine proteases SP1 and SP13 mediate the melanization response of Asian corn borer, Ostrinia furnacalis, against entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2015, 128, 64–72. [Google Scholar] [CrossRef]
- Rodnina, M.V.; Wolfgang, W. The ribosome as a molecular machine: The mechanism of tRNA-mRNA movement in translocation. Biochem. Soc. Trans. 2011, 39, 658–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodsky, F.M.; Guagliardi, L.E.; Galloway, C.J.; Koppelman, B.; Acton, S.L.; Wong, D.H.; Nathke, I. Endocytosis: Molecular mechanism and role in the immune response. Endocytosis 1992, 62, 343–353. [Google Scholar]
- Nonnenmacher, M.; Weber, T. Adeno-associated virus 2 infection requires endocytosis through the CLIC/GEEC pathway. Cell Host Microbe 2011, 10, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauza, C.D.; Price, T.M. Human immunodeficiency virus infection of T cells and monocytes proceeds via receptor-mediated endocytosis. J. Cell Biol. 1988, 107, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Branza-Nichita, N.; Macovei, A.; Lazar, C. Caveolae-Dependent Endocytosis in Viral Infection; Ceresa, B., Ed.; Intech: London, UK, 2012. [Google Scholar]
- Kumar, Y.; Valdivia, R.H. Leading a sheltered life: Intracellular pathogens and maintenance of vacuolar compartments. Cell Host Microbe 2009, 5, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Calvo, A.; Sobrino, F.; Martin-Acebes, M.A. Plasma membrane phosphatidylinositol 4, 5 bisphosphate is required for internalization of foot-and mouth disease virus and vesicular stomatitis virus. PLoS ONE 2012, 7, e45172. [Google Scholar] [CrossRef] [Green Version]
- Cicero, J.M.; Fisher, T.W.; Qureshi, J.A.; Stansly, P.A.; Brown, J.K. Colonization and intrusive invasion of potato psyllid by ‘Candidatus Liberibacter solanacearum’. Phytopathology 2017, 107, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Harries, P.A.; Schoelz, J.E.; Nelson, R.S. Intracellular transport of viruses and their components: Utilizing the cytoskeleton and membrane highways. Mol. Plant Microbe Interact. 2010, 23, 1381–1393. [Google Scholar] [CrossRef]
- Colonne, P.M.; Winchell, C.G.; Voth, D.E. Hijacking host cell highways: Manipulation of the host actin cytoskeleton by obligate intracellular bacterial pathogens. Front. Cell. Infect. Microbiol. 2016, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.J.; Hu, H.; Killiny, N. Proteomic maps of subcellular protein fractions of the Asian citrus psyllid Diaphorina citri, the vector of citrus huanglongbing. Physiol. Entomol. 2017, 42, 36–64. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CLas-Free-1 | CLas-Free-2 | CLas-Free-3 | CLas-Infected-1 | CLas-Infected-2 | CLas-Infected-3 |
|---|---|---|---|---|---|---|
| Total raw reads | 62,582,728 | 58,399,847 | 56,575,636 | 57,180,938 | 63,335,414 | 54,887,010 |
| Total clean reads | 61,521,780 | 57,557,156 | 55,768,652 | 55,707,048 | 62,309,800 | 53,907,044 |
| Q20 | 97.25% | 97.14% | 97.37% | 97.59% | 97.16% | 97.48% |
| Q30 | 92.45% | 92.20% | 92.72% | 93.13% | 92.24% | 92.90% |
| GC percent | 40.57% | 40.46% | 40.91% | 37.74% | 39.94% | 39.39% |
| Total map | 43,131,403 (70.11%) | 40,754,962 (70.81%) | 40,200,207 (72.08%) | 32,534,301 (58.4%) | 41,738,828 (66.99%) | 35,207,811 (65.31%) |
| Unique map | 34,876,548 (56.69%) | 32,834,545 (57.05%) | 32,347,622 (58.0%) | 26,881,942 (48.26%) | 33,556,647 (53.85%) | 28,376,185 (52.64%) |
| Multiple map | 8,254,855 (13.42%) | 7,920,417 (13.76%) | 7,852,585 (14.08%) | 5,652,359 (10.15%) | 8,182,181 (13.13%) | 6,831,626 (12.67%) |
| Read 1 map | 17,503,161 (28.45%) | 16,493,012 (28.66%) | 16,205,676 (29.06%) | 13,455,363 (24.15%) | 16,865,739 (27.07%) | 14,200,948 (26.34%) |
| Read 2 map | 17,373,387 (28.24%) | 16,341,533 (28.39%) | 16,141,946 (28.94%) | 13,426,579 (24.1%) | 16,690,908 (26.79%) | 14,175,237 (26.3%) |
| Positive map | 17,505,192 (28.45%) | 16,467,596 (28.61%) | 16,228,967 (29.1%) | 13,459,583 (24.16%) | 16,829,779 (27.01%) | 14,230,468 (26.4%) |
| Gene ID | Gene Description | CLas-Free FPKM | CLas-Infected FPKM | Log2Fold Change |
|---|---|---|---|---|
| Ubiquitination | ||||
| DcitrP024565.1 | Proteasome subunit alpha type | 8.590624 | 18.91098 | 1.142678 |
| DcitrP057820.1 | S-phase kinase-associated protein 1 | 5.07435 | 12.83314 | 1.338850457 |
| DcitrP057935.1 | Ubiquitin-fold modifier-conjugating enzyme 1 | 7.195371 | 14.55307 | 1.015273471 |
| DcitrP026550.1 | Ubiquitin-associated domain-containing protein 1 | 10.65557 | 3.702037 | −1.531855619 |
| DcitrP066965.1 | Ubiquitin-like modifier-activating enzyme ATG7 | 29.43184 | 14.80442 | −0.993594896 |
| DcitrP055490.1 | E3 ubiquitin-protein ligase | 0.333095 | 2.28688 | 2.782229875 |
| DcitrP062105.1 | Proteasome activator complex subunit 3-like protein | 2.290002 | 13.83469 | 2.595512129 |
| DcitrP059695.1 | Proteasome subunit beta type | 1.753526 | 3.587919 | 2.414010482 |
| DcitrP081240.1 | Ubiquitin domain-containing protein UBFD1 | 8.095103 | 15.50153 | 0.93331621 |
| DcitrP077930.1 | 26S proteasome non-ATPase regulatory subunit 12 | 11.5905 | 20.46284 | 0.818448568 |
| DcitrP069975.1 | E3 ubiquitin-protein ligase MARCH2 | 78.9481 | 38.47541 | −1.034157481 |
| DcitrP069805.1 | E3 ubiquitin-protein ligase RNF8 | 203.473 | 97.33118 | −1.064416343 |
| DcitrP080605.1 | Ubiquitin-conjugating enzyme E2 J2 | 24.049 | 10.95165 | −1.132484873 |
| Immune response | ||||
| DcitrP022770.1 | CLIP domain-containing serine protease 2-like | 0.15049 | 2.354921 | 3.964562534 |
| DcitrP022775.1 | CLIPB-serine protease 4 | 0.569921 | 5.485922 | 3.275711152 |
| DcitrP046955.1 | Aminopeptidase | 0.223217 | 1.070738 | 2.276052354 |
| DcitrP016435.1 | Lysozyme i-type | 4.803546 | 19.42617 | 2.011353707 |
| DcitrP097315.1 | CLIPB-serine protease 5 | 2.744837 | 6.396924 | 1.223892409 |
| DcitrP029315.1 | CLIPB-serine protease 5-RA | 1.551907 | 3.488869 | 1.163879263 |
| DcitrP022220.1 | Apolipophorin-Ⅲ | 82.6109 | 184.9353 | 1.162289806 |
| DcitrP036745.1 | Toll-like 8B | 66.77792 | 39.1133 | −0.771504993 |
| DcitrP077945.1 | Beat protein | 1.732096 | 0.063739 | −4.609706023 |
| DcitrP077305.1 | Cathespin F-like protein 5-RA | 0.014575 | 0.0578843 | 5.280394143 |
| DcitrP036095.1 | Spondin-2-like | 0.036088 | 0.612192 | 4.060891064 |
| Ribosome | ||||
| DcitrP073005.1 | 60S ribosomal protein L23 | 28.24808 | 126.5357 | 2.162205397 |
| DcitrP093430.1 | Ribosomal protein L27A | 98.15065 | 435.6547 | 2.152761801 |
| DcitrP013835.1 | 40S ribosomal protein S9 | 4.209662 | 13.43044 | 1.666951923 |
| DcitrP008150.1 | 60S ribosomal protein L34 | 17.65276 | 51.35397 | 1.536118663 |
| DcitrP059870.1 | 40S ribosomal protein S24 | 155.3869 | 427.2229 | 1.458686099 |
| DcitrP062275.1 | Ribosomal protein L15 | 101.5892 | 271.7571 | 1.418811677 |
| DcitrP059845.1 | 40S ribosomal protein S24 | 21.11473 | 55.26206 | 1.385121791 |
| DcitrP068410.1 | 28S ribosomal protein S18a, mitochondrial | 43.2703 | 102.078 | 1.235994788 |
| DcitrP007765.1 | Ribosomal RNA processing protein 36 like protein | 4.525509 | 10.3174 | 1.186528712 |
| DcitrP019375.1 | 60S ribosomal protein L21 | 206.8503 | 463.4805 | 1.16374505 |
| DcitrP027780.1 | 40S ribosomal protein S9 | 419.7915 | 883.7374 | 1.073779521 |
| DcitrP025515.1 | 60S ribosomal protein L32 | 533.5668 | 1100.274 | 1.044015195 |
| DcitrP002745.1 | 40S ribosomal protein S11 | 102.8649 | 207.1952 | 1.009873361 |
| DcitrP094130.1 | 60S ribosomal protein L13 | 48.45802 | 96.80903 | 0.997246375 |
| DcitrP037015.1 | 28S ribosomal protein S27, mitochondrial | 6.245021 | 3.043301 | −1.04281238 |
| Gene ID | Gene Description | CLas-Free FPKM | CLas-Infected FPKM | Log2Fold Change |
|---|---|---|---|---|
| Endocytosis | ||||
| DcitrP017415.1 | Heat shock protein 70 | 4.970658 | 34.09295 | 2.779406588 |
| DcitrP097875.1 | Multivesicular body subunit 12B | 24.38566 | 8.356045 | −1.541534978 |
| DcitrP031105.1 | WASH complex subunit 7-like protein | 17.45894 | 7.028736 | −1.315473724 |
| DcitrP034560.1 | Partioning defective 6 like protein gamma | 26.1217 | 9.444002 | −1.46961334 |
| DcitrP007925.1 | ADP-ribosylation factor | 16.66552 | 6.056527 | −1.459442252 |
| DcitrP079685.1 | Arf-GAP with SH3 domain | 6.149781 | 2.706943 | −1.176262825 |
| novel.2039 | Uncharacterized LOC113466219 | 48.38901 | 23.34858 | −1.053542718 |
| Cytoskeleton | ||||
| DcitrP027020.1 | Paramyosin-like protein | 6.933056 | 15.75305 | 1.185025088 |
| DcitrP028800.1 | Troponin I | 29.9198 | 67.87614 | 1.18133546 |
| DcitrP031990.1 | Tyrosine protein kinase receptor torso | 2.771104 | 1.295568 | −1.104829478 |
| DcitrP084355.1 | Troponin I | 0.433339 | 3.163564 | 2.881484544 |
| DcitrP056675.1 | Alpha tubulin 1 chain-like | 9.993052 | 47.73973 | 2.256687099 |
| DcitrP062610.1 | Myosin light chain kinase | 0.72003 | 2.930362 | 2.021891929 |
| DcitrP046925.1 | Transgelin | 9.711448 | 22.08303 | 1.186590264 |
| DcitrP015360.1 | Kinesin-like protein | 1.97132 | 4.45914 | 1.180135096 |
| DcitrP016785.1 | Alpha tubulin 1 chain-like | 6.096831 | 53.22063 | 3.126504327 |
| DcitrP034805.1 | Gelsolin | 17.55555 | 120.6549 | 2.780374779 |
| DcitrP011480.1 | Troponin C | 0.928841 | 6.230001 | 2.755828695 |
| DcitrP027235.1 | Dynein light chain Tctex-type 1 | 0.84466 | 5.503671 | 2.705884305 |
| DcitrP081065.1 | Alpha tubulin 1 chain-like | 1.522667 | 7.639856 | 2.330954816 |
| DcitrP062790.1 | Keratin, type I cytoskeletal 9 | 3.797606 | 10.28138 | 1.432892752 |
| DcitrP002545.1 | Dynein heavy chain | 1.317832 | 0.439357 | −1.557423656 |
| Insecticide resistance | ||||
| DcitrP079275.1 | Cytochrome P450 CYP4C4 | 0.331885 | 6.304564 | 4.238246725 |
| DcitrP045800.1 | Probable cytochrome P450 6a14 | 0.172821 | 2.734463 | 4.010391446 |
| DcitrP079270.1 | CYP301A1 partial | 1.019659 | 11.5746 | 3.500092719 |
| DcitrP079255.1 | CYP301A1 partial | 2.045632 | 13.91084 | 2.763107035 |
| DcitrP039115.1 | ATP-binding cassette sub-family G member 5 | 0.349581 | 1.914687 | 2.457939089 |
| DcitrP019570.1 | Glutathione S-transferase | 108.29 | 310.5219 | 1.519243483 |
| DcitrP022100.1 | CYP3167A2 partial | 3.10311 | 8.045237 | 1.366858366 |
| DcitrP071995.1 | CYP3172A5 partial | 1.847749 | 4.73452 | 1.355174187 |
| DcitrP019590.1 | Glutathione S-transferase | 14.94143 | 37.15758 | 1.311635196 |
| DcitrP069025.1 | ATP-binding cassette sub-family G member 4 | 3.412652 | 8.348752 | 1.294131073 |
| DcitrP042025.1 | ATP-binding cassette sub-family A member | 21.79537 | 9.547165 | −1.187067336 |
| DcitrP090960.1 | Multidrug resistance-associated protein 1 | 56.71604 | 22.4627 | −1.335398633 |
| DcitrP013660.1 | CYP6KB1 | 17.94354 | 6.599974 | −1.448082102 |
| DcitrP078955.1 | ATP-binding cassette sub-family A member 3 | 81.79196 | 9.226527 | −1.549025159 |
| DcitrP051650.1 | Carboxylic ester hydrolase | 8.538477 | 2.508855 | −1.773370392 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-Z.; Li, N.-Y.; Zeng, X.-D.; Song, J.-C.; Yu, X.-D.; Su, H.-N.; Chen, C.-X.; Yi, L.; Lu, Z.-J. Transcriptome Analyses of Diaphorina citri Midgut Responses to Candidatus Liberibacter Asiaticus Infection. Insects 2020, 11, 171. https://doi.org/10.3390/insects11030171
Yu H-Z, Li N-Y, Zeng X-D, Song J-C, Yu X-D, Su H-N, Chen C-X, Yi L, Lu Z-J. Transcriptome Analyses of Diaphorina citri Midgut Responses to Candidatus Liberibacter Asiaticus Infection. Insects. 2020; 11(3):171. https://doi.org/10.3390/insects11030171
Chicago/Turabian StyleYu, Hai-Zhong, Ning-Yan Li, Xiang-Dong Zeng, Jian-Chun Song, Xiu-Dao Yu, Hua-Nan Su, Ci-Xiang Chen, Long Yi, and Zhan-Jun Lu. 2020. "Transcriptome Analyses of Diaphorina citri Midgut Responses to Candidatus Liberibacter Asiaticus Infection" Insects 11, no. 3: 171. https://doi.org/10.3390/insects11030171