First Report of Aleurocanthus spiniferus on Ailanthus altissima: Profiling of the Insect Microbiome and MicroRNAs

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Identification
2.2. Denaturing Gradient Gel Electrophoresis (DGGE)
2.3. Deep Sequencing of Small RNAs
2.4. NGS Data Analysis
2.4.1. Microbiomics
2.4.2. Classification of Small RNAs
2.4.3. Identification of Known and Novel MicroRNAs Including IsomiRs
3. Results and Discussion
3.1. Aleurocanthus spiniferus Was Found for the First Time on Ailanthus altissima in Italy
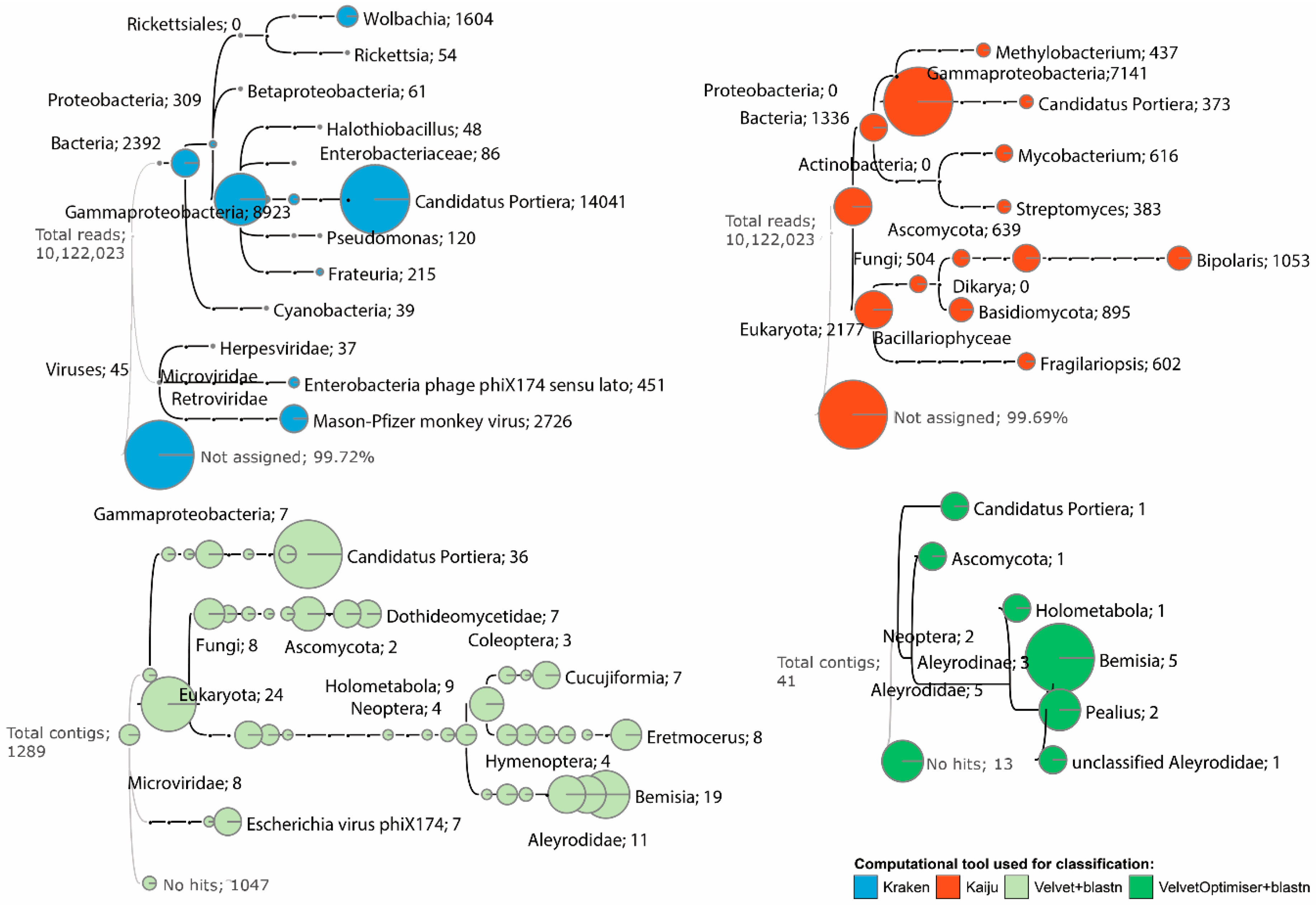
3.2. “Candidatus Portiera”, Wolbachia, and Rickettsia Were Identified in the OSW Microbiome
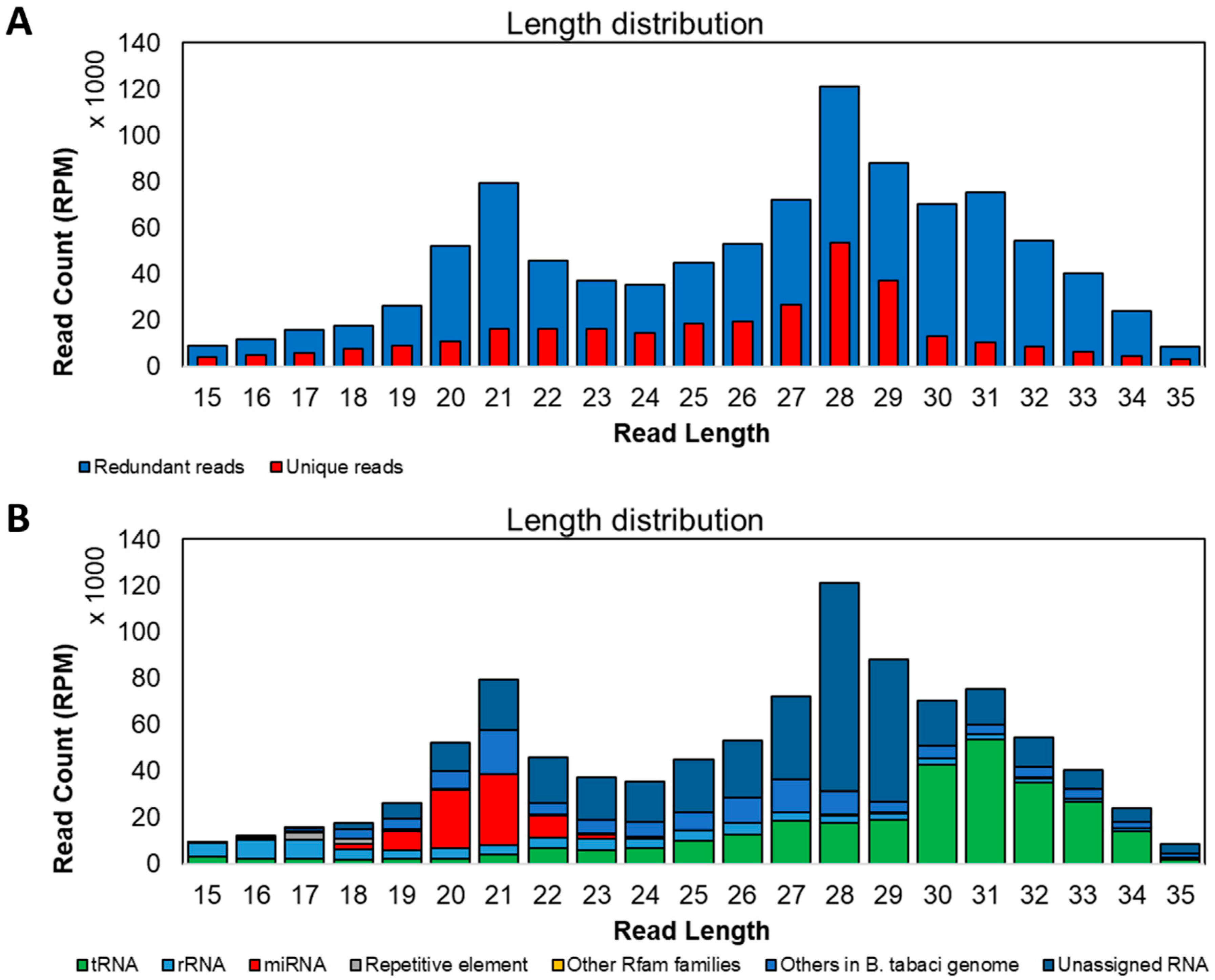
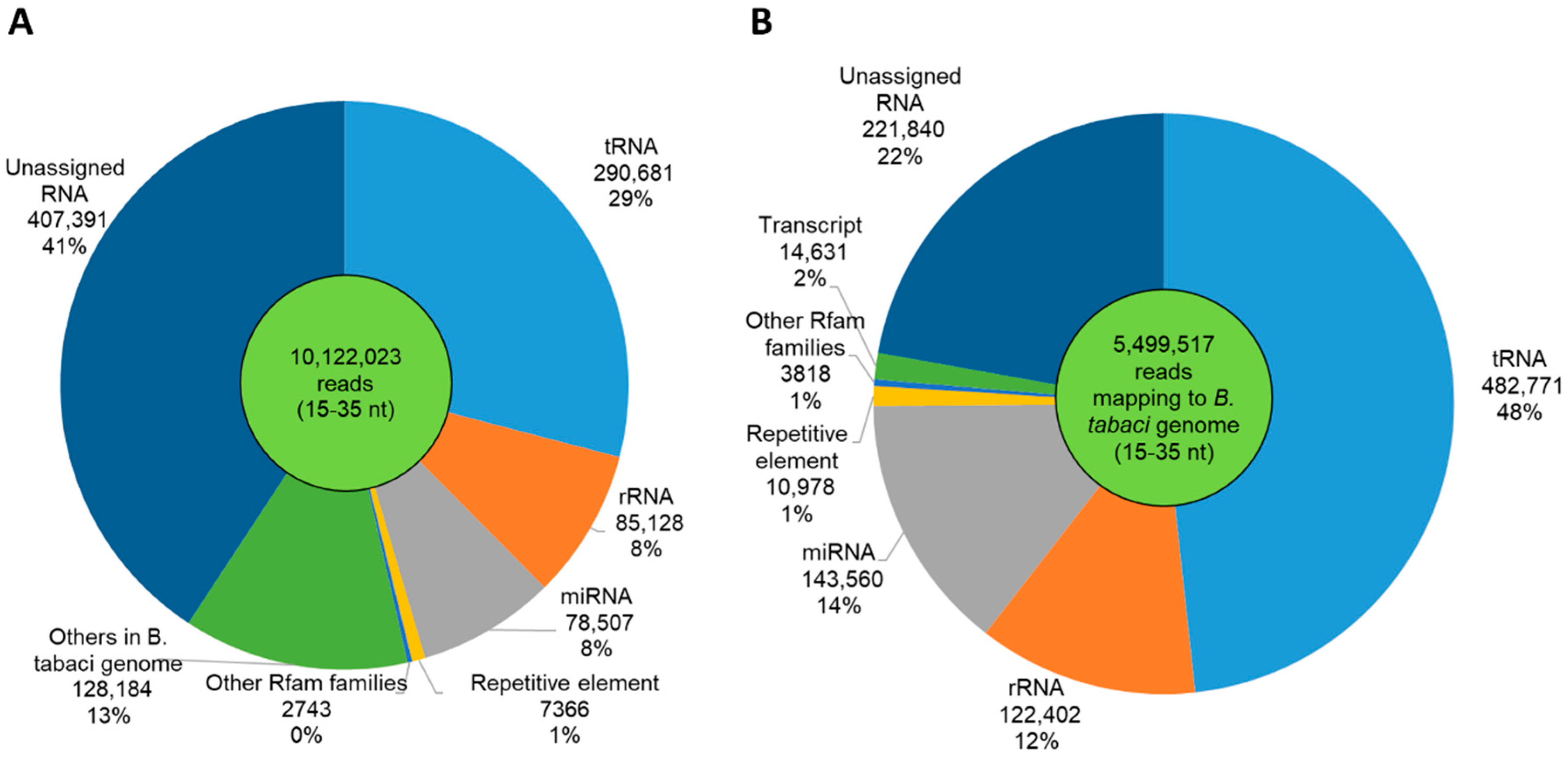
3.3. Features of Small RNAs
3.4. MicroRNAs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Porcelli, F. First record of Aleurocanthus spiniferus (Homoptera: Aleyrodidae) in Apulia, Southern Italy. EPPO Bull. 2008, 38, 516–518. [Google Scholar] [CrossRef]
- Kapantaidaki, D.E.; Antonatos, S.; Kontodimas, D.; Milonas, P.; Papachristos, D.P. Presence of the invasive whitefly Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Greece. EPPO Bull. 2019, 49, 127–131. [Google Scholar] [CrossRef]
- Nugnes, F.; Laudonia, S.; Jesu, G.; Jansen, M.G.M.; Porcelli, F.; Bernardo, U. Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Europe: Diffusion, hosts, molecular characterization, and its natural enemies. Insects 2020, 11, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, M.; Porcelli, F. Aleurocanthus camelliae (Hemiptera: Aleyrodidae), a species possibly new for the European fauna of a genus in great need of revision. Tijdschr. Entomol. 2018, 161, 63–78. [Google Scholar] [CrossRef]
- EPPO. EPPO Global Database. Aleurocanthus spiniferus (ALECSN). Available online: https://gd.eppo.int/taxon/ALECSN (accessed on 23 October 2019).
- Cioffi, M.; Cornara, D.; Corrado, I.; Jansen, M.G.M.; Porcelli, F. The status of Aleurocanthus spiniferus from its unwanted introduction in Italy to date. B. Insectol. 2013, 66, 273–281. [Google Scholar]
- EFSA; Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Jaques Miret, J.A.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; et al. Pest categorisation of Aleurocanthus spp. EFSA J. 2018, 16, e05436. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium. Ailanthus altissima. Datasheet 3889. Available online: https://www.cabi.org/isc/datasheet/3889 (accessed on 23 October 2019).
- EPPO. EPPO Global Database. Ailanthus altissima (AILAL). Available online: https://gd.eppo.int/taxon/AILAL (accessed on 23 October 2019).
- Maschek, O.; Halmschlager, E. Effects of Verticillium nonalfalfae on Ailanthus altissima and associated indigenous and invasive tree species in eastern Austria. Eur. J. For. Res. 2018, 137, 197–209. [Google Scholar] [CrossRef]
- Schall, M.J.; Davis, D.D. Verticillium wilt of Ailanthus altissima: Susceptibility of associated tree species. Plant Dis. 2009, 93, 1158–1162. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Wu, Y.; Zheng, H.; Fu, W.; Reardon, R.; Liu, M. Assessing potential biological control of the invasive plant, tree-of-heaven, Ailanthus altissima. Biocontrol Sci. Techn. 2006, 16. [Google Scholar] [CrossRef]
- O’Neal, E.S.; Davis, D.D. Biocontrol of Ailanthus altissima: Inoculation protocol and risk assessment for Verticillium nonalfalfae (Plectosphaerellaceae: Phyllachorales). Biocontrol Sci. Techn. 2015, 25, 950–969. [Google Scholar] [CrossRef]
- Su, Q.; Xie, W.; Wang, S.; Wu, Q.; Ghanim, M.; Zhang, Y. Location of symbionts in the whitefly Bemisia tabaci affects their densities during host development and environmental stress. PLoS ONE 2014, 9, e91802. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Xie, W.; Wang, S.; Wu, Q.; Liu, B.; Fang, Y.; Xu, B.; Zhang, Y. The endosymbiont Hamiltonella increases the growth rate of its host Bemisia tabaci during periods of nutritional stress. PLoS ONE 2014, 9, e89002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnegan, E.J.; Matzke, M.A. The small RNA world. J. Cell Sci. 2003, 116, 4689–4693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Osimani, A.; Milanović, V.; Garofalo, C.; Cardinali, F.; Roncolini, A.; Sabbatini, R.; De Filippis, F.; Ercolini, D.; Gabucci, C.; Petruzzelli, A.; et al. Revealing the microbiota of marketed edible insects through PCR-DGGE, metagenomic sequencing and real-time PCR. Int. J. Food Microbiol. 2018, 276, 54–62. [Google Scholar] [CrossRef]
- Nugnes, F.; Gebiola, M.; Monti, M.M.; Gualtieri, L.; Giorgini, M.; Wang, J.; Bernardo, U. Genetic diversity of the invasive gall wasp Leptocybe invasa (Hymenoptera: Eulophidae) and of its Rickettsia endosymbiont, and associated sex-ratio differences. PLoS ONE 2015, 10, e0124660. [Google Scholar] [CrossRef] [Green Version]
- Gualtieri, L.; Nugnes, F.; Nappo, A.G.; Gebiola, M.; Bernardo, U. Life inside a gall: Closeness does not favour horizontal transmission of Rickettsia between a gall wasp and its parasitoid. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Monti, M.M.; Nugnes, F.; Gualtieri, L.; Gebiola, M.; Bernardo, U. No evidence of parthenogenesis-inducing bacteria involved in Thripoctenus javae thelytoky: An unusual finding in Chalcidoidea. Entomol. Exp. Appl. 2016, 160, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Yu, T.; Xie, S.; Du, K.; Liang, X.; Lan, Y.; Sun, C.; Lu, X.; Shao, Y. Comparative shotgun metagenomic data of the silkworm Bombyx mori gut microbiome. Sci. Data 2018, 5, 180285. [Google Scholar] [CrossRef]
- Krishnan, M.; Bharathiraja, C.; Pandiarajan, J.; Prasanna, V.A.; Rajendhran, J.; Gunasekaran, P. Insect gut microbiome - An unexploited reserve for biotechnological application. Asian Pac. J. Trop. Biomed. 2014, 4, S16–S21. [Google Scholar] [CrossRef]
- Malacrinò, A. Meta-omics tools in the world of insect-microorganism interactions. Biology 2018, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.J.; Chen, Y.T.; Bonning, B.C. RNA virus discovery in insects. Curr. Opin. Insect Sci. 2015, 8, 54–61. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.G.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLlife 2015, 4, e05378. [Google Scholar] [CrossRef]
- Liu, S.; Vijayendran, D.; Bonning, B.C. Next generation sequencing technologies for insect virus discovery. Viruses 2011, 3, 1849–1869. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.X.; Ding, S.W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcelli, F. A Method to Slide-Mount Insects and Other Arthropods in a Water-Soluble Medium. Available online: https://zenodo.org/record/3471649#.XbiFD-hKjIU (accessed on 29 October 2019).
- Gebiola, M.; Bernardo, U.; Monti, M.M.; Navone, P.; Viggiani, G. Pnigalio agraules (Walker) and Pnigalio mediterraneus Ferrière and Delucchi (Hymenoptera: Eulophidae): Two closely related valid species. J. Nat. Hist. 2009, 43, 2465–2480. [Google Scholar] [CrossRef]
- Dar, S.A.; Kuenen, J.G.; Muyzer, G. Nested PCR-denaturing gradient gel electrophoresis approach to determine the diversity of sulfate-reducing bacteria in complex microbial communities. Appl. Environ. Microbiol. 2005, 71, 2325–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assaf, G. FASTQ/A short-reads pre-processing tools. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 23 October 2019).
- Blankenberg, D.; Gordon, A.; Von Kuster, G.; Coraor, N.; Taylor, J.; Nekrutenko, A.; The Galaxy Team. Manipulation of FASTQ data with Galaxy. Bioinformatics 2010, 26, 1783–1785. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Gladman, S. VelvetOptimiser. Available online: https://github.com/slugger70/VelvetOptimiser (accessed on 23 October 2019).
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.-J.; Tappu, R. MEGAN Community Edition—Interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2018, 47, D155–D162. [Google Scholar] [CrossRef]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.M.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A uniform system for microRNA annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Axtell, M.J.; Bartel, B.; Bartel, D.P.; Baulcombe, D.; Bowman, J.L.; Cao, X.; Carrington, J.C.; Chen, X.M.; Green, P.J.; et al. Criteria for annotation of plant microRNAs. Plant Cell 2008, 20, 3186–3190. [Google Scholar] [CrossRef]
- Morin, R.D.; O’Connor, M.D.; Griffith, M.; Kuchenbauer, F.; Delaney, A.; Prabhu, A.-L.; Zhao, Y.; McDonald, H.; Zeng, T.; Hirst, M.; et al. Application of massively parallel sequencing to microRNA profiling and discovery in human embryonic stem cells. Genome Res. 2008, 18, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, L.F.; Christoff, A.P.; Margis, R. isomiRID: A framework to identify microRNA isoforms. Bioinformatics 2013, 29, 2521–2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, N.; Singh, A.; Rana, V.S.; Rajagopal, R. Molecular characterization and analysis of bacterial diversity in Aleurocanthus woglumi (Hemiptera: Aleyrodidae). Environ. Entomol. 2013, 42, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Sinkins, S.P. Wolbachia and cytoplasmic incompatibility in mosquitoes. Insect. Biochem. Mol. Biol. 2004, 34, 723–729. [Google Scholar] [CrossRef]
- Weinert, L.A.; Werren, J.H.; Aebi, A.; Stone, G.N.; Jiggins, F.M. Evolution and diversity of Rickettsia bacteria. BMC Biol. 2009, 7, 6. [Google Scholar] [CrossRef]
- Wang, H.-L.; Lei, T.; Xia, W.-Q.; Cameron, S.L.; Liu, Y.-Q.; Zhang, Z.; Gowda, M.M.N.; De Barro, P.; Navas-Castillo, J.; Omongo, C.A.; et al. Insight into the microbial world of Bemisia tabaci cryptic species complex and its relationships with its host. Sci. Rep. 2019, 9, 6568. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.F.; Xia, F.; Johnson, K.W.; Brown, C.D.; Bartom, E.; Tuteja, J.H.; Stevens, R.; Grossman, R.L.; Brumin, M.; White, K.P.; et al. Comparison of the genome sequences of “Candidatus Portiera aleyrodidarum” primary endosymbionts of the whitefly Bemisia tabaci B and Q biotypes. Appl. Environ. Microbiol. 2013, 79, 1757–1759. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, Y.; Ghanim, M.; Gueguen, G.; Kontsedalov, S.; Vavre, F.; Fleury, F.; Zchori-Fein, E. Inherited intracellular ecosystem: Symbiotic bacteria share bacteriocytes in whiteflies. FASEB J. 2008, 22, 2591–2599. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Su, Q.; Jiao, X.; Zhou, L.; Liu, B.; Xie, W.; Wang, S.; Wu, Q.; Xu, B.; Zhang, Y. Relative amount of symbionts in Bemisia tabaci (Gennadius) Q changes with host plant and establishing the method of analyzing free amino acid in B. tabaci. Commun. Integr. Biol. 2013, 6, e23397. [Google Scholar] [CrossRef]
- Rosell, R.C.; Davidson, E.W.; Jancovich, J.K.; Hendrix, D.L.; Brown, J.K. Size limitations in the filter chamber and digestive tract of nymphal and adult Bemisia tabaci whiteflies (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2003, 96, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Hanan, A.; He, X.Z.; Wang, Q. Insight into the success of whitefly biological control using parasitoids: Evidence from the Eretmocerus warrae−Trialeurodes vaporariorum system. Pest Manag. Sci. 2017, 73, 2294–2301. [Google Scholar] [CrossRef] [PubMed]
- Leschen, R.A.B.; Ślipiński, A. 7. Cucujiformia Lameere, 1938. Introduction, phylogeny. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera Beetles Vol. 2. Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim); Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; Walter de Gruyter GmbH & Co. KG: Berlin/New York, NY, USA, 2010; pp. 227–228. [Google Scholar]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Family-herpesviridae. In Virus Taxonomy; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 111–122. [Google Scholar]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Family-herpesviridae. In Virus Taxonomy; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 477–495. [Google Scholar]
- Montiel, N.A. An updated review of simian betaretrovirus (SRV) in macaque hosts. J. Med. Primatol. 2010, 39, 303–314. [Google Scholar] [CrossRef]
- Junglen, S.; Drosten, C. Virus discovery and recent insights into virus diversity in arthropods. Curr. Opin. Microbiol. 2013, 16, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Huntemann, M.; Ivanova, N.; Kyrpides, N.C.; Pati, A. Large-scale contamination of microbial isolate genomes by Illumina PhiX control. Stand. Genomic Sci. 2015, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wang, L.; Chen, F.; Yang, X.; Ding, M.; Zhang, Z.; Liu, S.-S.; Wang, X.-W.; Zhou, X. MicroRNA profiling of the whitefly Bemisia tabaci Middle East-Aisa Minor I following the acquisition of Tomato yellow leaf curl China virus. Virol. J. 2016, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Tao, Y.L.; Chu, D. Characterization and comparative profiling of miRNAs in invasive Bemisia tabaci (Gennadius) B and Q. PLoS ONE 2013, 8, e59884. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhao, J.; Wang, J.; Hu, C.; Peng, J.; Luo, R.; Zhou, C.; Liu, J.; Lin, J.; Jin, Y.; et al. MicroRNAs are involved in the regulation of ovary development in the pathogenic blood fluke Schistosoma japonicum. PLoS Pathog. 2016, 12, e1005423. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Han, S.; Chen, T.; Qin, G.; Li, L.; Guo, X. Involvement of microRNAs in infection of silkworm with Bombyx mori cytoplasmic polyhedrosis virus (BmCPV). PLoS ONE 2013, 8, e68209. [Google Scholar] [CrossRef]
- Ruby, J.G.; Stark, A.; Johnston, W.K.; Kellis, M.; Bartel, D.P.; Lai, E.C. Evolution, biogenesis, expression, and target predictions of a substantially expanded set of Drosophila microRNAs. Genome Res. 2007, 17, 1850–1864. [Google Scholar] [CrossRef] [Green Version]
- Stark, A.; Bushati, N.; Jan, C.H.; Kheradpour, P.; Hodges, E.; Brennecke, J.; Bartel, D.P.; Cohen, S.M.; Kellis, M. A single Hox locus in Drosophila produces functional microRNAs from opposite DNA strands. Genes Dev. 2008, 22, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Ylla, G.; Fromm, B.; Piulachs, M.D.; Belles, X. The microRNA toolkit of insects. Sci. Rep. 2016, 6, 37736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smibert, P.; Lai, E.C. A view from Drosophila: Multiple biological functions for individual microRNAs. Semin. Cell Dev. Biol. 2010, 21, 745–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. Bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Huang, Y.; Zhang, Q.; Zhou, H.; Jin, P.; Ma, F. The miR-317 functions as a negative regulator of Toll immune response and influences Drosophila survival. Dev. Comp. Immunol. 2019, 95, 19–27. [Google Scholar] [CrossRef]
- Lampe, L.; Jentzsch, M.; Levashina, E.A. Metabolic balancing by miR-276 shapes the mosquito reproductive cycle and Plasmodium falciparum development. bioRxiv 2019, 548784. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Chen, Q.; Wei, Y.; Jiang, F.; Yang, M.; Hao, S.; Guo, X.; Chen, D.; Kang, L. MicroRNA-276 promotes egg-hatching synchrony by up-regulating brm in locusts. Proc. Natl. Acad. Sci. USA 2016, 113, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Ronshaugen, M.; Biemar, F.; Piel, J.; Levine, M.; Lai, E.C. The Drosophila microRNA iab-4 causes a dominant homeotic transformation of halteres to wings. Genes Dev. 2005, 19, 2947–2952. [Google Scholar] [CrossRef] [Green Version]
- Rubio, M.; de Horna, A.; Belles, X. MicroRNAs in metamorphic and non-metamorphic transitions in hemimetabolan insect metamorphosis. BMC Genomics 2012, 13, 386. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.; Cui, T.; Grigoriev, A. Visualization of nucleotide substitutions in the (micro) transcriptome. BMC Genomics 2014, 15 (Suppl. 4), S9. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provisional MiRNA Name (Mature/Star) | miRDeep2 ID | miRDeep2 Score | Read Count (RPM) | Example of miRBase miRNA with the Same Seed | Alignment between A. spiniferus miRNA (up) and miRbase miRNA (Bottom) | Precursor Coordinate in the Bemisia tabaci Genome | |
|---|---|---|---|---|---|---|---|
| Mature | Star | ||||||
| asp-bantam-3p asp-bantam-5p | NW_017547202.1_5067 | 5.2 | 115,812.7 | 5.5 | dme-bantam-3p | UGAGAUCAUCGUGAAAGCUG .........UU.........AUU | NW_017547202.1:1932321..1932378: + |
| asp-miR-276a-3p asp-miR-276a-5p | NW_017547107.1_1308 | 5 × 104 | 17,936.8 | 10.8 | dme-miR-276a-3p | UAGGAACUUCAUACCGUGCUC .....................U | NW_017547107.1:5694695..5694839: − b |
| asp-miR-317-3p asp-miR-317-5p | NW_017550575.1_11879 | 1.1 × 104 | 4034.0 | 4.0 | dqu-miR-317-3p | UGAACACAGCUGGUGGUAUCUCAG ........................ | NW_017550575.1:773861..774000: + b |
| asp-miR-750-3p asp-miR-750-5p | NW_017547443.1_8884 | 9.4 × 103 | 3348.7 | 18.5 | ame-miR-750-3p | CCAGAUCUAACUCUUCCAGCUC ...................... | NW_017547443.1:370244..370308: − |
| asp-mir-8-3p asp-mir-8-5p | NW_017547115.1_2104 | 4.4 | 3101.0 | 3.7 | api-mir-8 | UAAUACUGUCAGGUAAUGAU ....................GUC | NW_017547115.1:1766399..1766456: − |
| asp-miR-184-3p asp-miR-184-5p | NW_017563531.1_15559 | 5.1 | 26.2 | 0 | dme-miR-184-3p | UGGACGGAAAACUGAUAAGGGU ........G............C | NW_017563531.1:2205..2265: + |
| NW_017552810.1_13306 | 4.8 | 26.2 | 0 | UGGACGGAAAACUGAUAAGGGU ........G............C | NW_017552810.1:1753402..1753462: − | ||
| NW_017547113.1_1681 | 5 | 3074.1 | 0.6 | UGGACGGAGAACUGAUAAGGG .....................C | NW_017547113.1:1014029..1014184: + b | ||
| asp-miR-2765-5p asp-miR-2765-3p | NW_017547098.1_412 | 3.8 × 103 | 1355.4 | 5.1 | ame-miR-2765-5p | UGGUAACUCCACCACCGUUGG .....................C | NW_017547098.1:2423456..2423512: + |
| NW_017547098.1_410 | 4.8 | 2424.9 | 0 | UGGUAACUCCACACCACCGUUGG ............--.........C | NW_017547098.1:2422100..2422160: + | ||
| asp-miR-100-5p asp-miR-100-3p | NW_017547299.1_7860 | 4.9 × 103 | 1758.5 | 2.0 | dme-miR-100-5p | AACCCGUAGAUCCGAACUUGU .....................G | NW_017547299.1:803546..803601: + |
| asp-miR-13a-3p asp-miR-13a-5p | NW_017547352.1_8563 | 4.6 × 103 | 1617.4 | 49.2 | tca-miR-13a-3p | UAUCACAGCCACUUUGAUGAAC ....................G. | NW_017547352.1:37273..37334: + |
| asp-miR-2a-3p asp-miR-2a-5p | NW_017547352.1_8561 | 2.9 × 103 | 1039.5 | 0.2 | mse-miR-2a | UCACAGCCAGCUUUGAUGAGCA ...................... | NW_017547352.1:36468..36531: + |
| NW_017547352.1_8569 | 1.7 × 103 | 636.6 | 0.9 | dme-miR-2a-3p | UAUCACAGCCAGCUUUGAUGA .....................GC | NW_017547352.1:40230..40282: + | |
| NW_017547352.1_8567 | 5.4 | 638.8 | 1.1 | NW_017547352.1:40048..40201: + b | |||
| NW_017561640.1_15257 | 1.7 × 103 | 636.6 | 0.9 | NW_017561640.1:532..683: + b | |||
| NW_017561640.1_15255 | 5.4 | 638.8 | 1.1 | NW_017561640.1:382..438: + | |||
| asp-miR-315-5p asp-miR-315-3p | NW_017547334.1_8451 | 4.8 | 1501.8 | 0 | dme-miR-315-5p | UUUUGAUUGUUGCUCAGAAAGC ...................... | NW_017547334.1:1004963..1005023: − |
| asp-miR-9a-5p asp-miR-9a-3p | NW_017551177.1_12283 | 3.9 × 103 | 1395.9 | 22.2 | dme-miR-9a-5p | UCUUUGGUUACCUAGCUGUAUG ..........T........... | NW_017551177.1:699591..699652: − |
| asp-miR-277-3p asp-miR-277-5p | NW_017550575.1_11881 | 2.9 × 103 | 1062.8 | 2.0 | dme-miR-277-3p | UAAAUGCACUAUCUGGUACGAC ......................A | NW_017550575.1:818061..818121: + |
| asp-miR-71-3p asp-miR-71-5p | NW_017547352.1_8559 | 2.8 × 103 | 860.6 | 139.8 | tca-miR-71-3p | UCUCACUACCUUGUCUUUCAU .....................G | NW_017547352.1:36323..36463: + b |
| asp-miR-14-3p asp-miR-14-5p | NW_017547098.1_493 | 2.5 × 103 | 847.3 | 57.8 | dme-miR-14-3p | UCAGUCUUUUUCUCUCUCCUA .....................U | NW_017547098.1:8908667..8908729: + |
| asp-miR-275-3p asp-miR-275-5p | NW_017547297.1_7629 | 5.6 | 820.1 | 101.1 | dme-miR-275-3p | UCAGGUACCUGAAGUAGCGCG .....................CG | NW_017547297.1:1465090..1465268: + b |
| asp-let-7-5p asp-let-7-3p | NW_017547299.1_7862 | 4.8 | 745.2 | 6.2 | dme-let-7-5p | UGAGGUAGUAGGUUGUAUAG ....................U | NW_017547299.1:803734..803802: + |
| asp-miR-263a-5p asp-miR-263a-3p | NW_017547161.1_3192 | 5.1 | 696.1 | 4.8 | dme-miR-263a-5p | AAUGGCACUAGAAGAAUUCACG .........G............GG | NW_017547161.1:912282..912340: + |
| asp-miR-31-5p asp-miR-31-5p | NW_017547107.1_1345 | 5 | 654.4 | 0 | aca-miR-31-5p | AGGCAAGAUGUUGGCAUAGC ....................U | NW_017547107.1:9048192..9048254: − |
| asp-miR-306-5p asp-miR-306-3p | NW_017547320.1_8138 | 5.3 | 597.3 | 1.5 | dme-miR-306-5p | UCAGGUACUGAGUGACUCU .........G......... | NW_017547320.1:2159894..2159951: + |
| asp-miR-305-5p asp-miR-305-3p | NW_017547297.1_7632 | 5.1 | 522.0 | 2.4 | dme-miR-305-5p | AUUGUACUUCAUCAGGUGCUC .....................UG | NW_017547297.1:1466885..1466944: + |
| asp-miR-92b-3p asp-miR-92b-5p | NW_017564582.1_15992 | 0.6 a | 488.4 | 0 | dme-miR-92b-3p | AAUUGCACUAGUCCCGGCCUG .....................C | NW_017564582.1:513918..513972: − |
| asp-miR-13b-3p asp-miR-13b-5p | NW_017561640.1_15253 | 1.1 × 103 | 311.8 | 109.2 | bmo-miR-13b | UAUCACAGCCAUUUUUGACGUGCC ....................A.U | NW_017561640.1:204..265:+ |
| NW_017547352.1_8565 | NW_017547352.1:39883..39944: + | ||||||
| asp-miR-34-5p asp-miR-34-3p | NW_017550575.1_11884 | 5.1 | 297.1 | 15.4 | dme-miR-34-5p | UGGCAGUGUGGUUAGCUGGUUG ......................UG | NW_017550575.1:854739..854841: + b |
| asp-miR-87a-3p asp-miR-87a-5p | NW_017547298.1_7717 | 2.8 × 102 | 98.7 | 2.8 | tca-miR-87a-3p | GUGAGCAAAGUUUCAGGUGUG ..................... | NW_017547298.1:2017127..2017227: + |
| NW_017547298.1_7719 | 8.2 × 102 | 285.8 | 5.7 | tca-miR-87a-3p | GUGAGCAACGUAUCAGGUGUC ........A..U........GA | NW_017547298.1:2017271..2017332: + | |
| asp-miR-993-3p asp-miR-993-5p | NW_017547290.1_7299 | 9.5 × 102 | 276.9 | 64.2 | dme-miR-993-3p | GAAGCUCGUCUCUACAGGUAUC ......................U | NW_017547290.1:727642..727815: + b |
| asp-miR-125-5p asp-miR-125-3p | NW_017547299.1_7864 | 7.8 × 102 | 262.4 | 14.9 | dme-miR-125-5p | UCCCUGAGACCCUAAUUUGUG ...............C.....A | NW_017547299.1:822960..823021: + |
| asp-miR-927-3p asp-miR-927-5p | NW_017547282.1_7013 | 8.7 × 102 | 227.3 | 83.3 | dme-miR-927-3p | CAAAGCGUUUGAAUUCUGGAA ...........G......A..C | NW_017547282.1:1940947..1941008: + |
| asp-miR-281-2-5p asp-miR-281-2-3p | NW_017561619.1_15248 | 8.1 × 102 | 219.8 | 68.8 | dme-miR-281-2-5p | AAGAGAGCUAUCCUUCGACAG .............G.......U | NW_017561619.1:12115..12174: + |
| asp-miR-252a-5p asp-miR-252a-3p | NW_017554870.1_13884 | 5.9 × 102a | 212.4 | 0.2 | dpu-miR-252a | CUAAGUACUCCGUGCCGCAGG ..........-..........AG | NW_017554870.1:193643..193702: + |
| asp-miR-1-3p asp-miR-1-5p | NW_017547904.1_9568 | 3.9 × 102 | 137.8 | 2.2 | dme-miR-1-3p | UGGAAUGUAAAGAAGUAUGGA .....................G | NW_017547904.1:1209882..1209940: − |
| asp-miR-190-5p asp-miR-190-3p | NW_017547096.1_359 | 3.3 × 102 | 117.3 | 0.6 | dme-miR-190-5p | AGAUAUGUUUGAUAUUCUUGGUU .......................G | NW_017547096.1:1449782..1449864: − b |
| asp-miR-279b-3p asp-miR-279b-5p | NW_017547145.1_3172 | 5.2 | 114.4 | 0 | pxy-miR-279b-3p | UGACUAGAUUUUCACUCAUUC ...................C. | NW_017547145.1:2285467..2285523: − |
| asp-miR-92a-3p asp-miR-92a-5p | NW_017564582.1_15994 | 5.1 | 86.9 | 0 | dme-miR-92a-3p | UAUUGCACUUGUCCCGGCC C..................UAU | NW_017564582.1:514087..514138: − |
| asp-miR-133-3p asp-miR-133-5p | NW_017547904.1_9564 | 2.3 × 102 | 83.8 | 0.2 | dme-miR-133-3p | UUGGUCCCCUUCAACCAGCUG .....................U | NW_017547904.1:1012752..1012934: − b |
| asp-miR-263b-5p asp-miR-263b-3p | NW_017547161.1_3196 | 2.4 × 102 | 81.3 | 3.7 | api-miR-263b | CUUGGCACUGGAAGAAUUCACAG .......................A | NW_017547161.1:950587..950647: + |
| asp-miR-252b-5p asp-miR-252b-3p | NW_017554870.1_13882 | 2 × 10 2a | 72.9 | 0.9 | api-miR-252b | CUAAGUAGUAGCGCCGAAGGUG ...............A.C....A | NW_017554870.1:149158..149217: + |
| asp-miR-124-3p asp-miR-124-5p | NW_017557672.1_14877 | 0.3 a | 64.0 | 37.4 | dme-miR-124-3p | UAAGGCACGCGGUGAAUGCC ....................AAG | NW_017557672.1:97524..97666: + b |
| asp-miR-375-3p asp-miR-375-5p | NW_017547090.1_76 | 1.5 × 102 | 53.5 | 1.5 | ame-miR-375-3p | UUUGUUCGUUCGGCUCGAGUU .....................A | NW_017547090.1:1647645..1647829: − b |
| asp-miR-10-5p asp-miR-10-3p | NW_017547290.1_7361 | 4.4 | 51.8 | 0.9 | oar-miR-10a | UACCCUGUAGAUCCGAAAUU .................U..G | NW_017547290.1:727642..727815: − b |
| asp-miR-996-3p asp-miR-996-5p | NW_017553907.1_13657 | 1.5 × 102 | 46.2 | 9.2 | api-miR-996 | UGACUAGAGUUACACUCGUC ....................A | NW_017553907.1:824153..824211: − |
| asp-miR-7-5p asp-miR-7-3p | NW_017556795.1_14461 | 1.1 × 102 | 38.2 | 1.1 | dme-miR-7-5p | UGGAAGACUAGUGAUUUUGUUGU ....................... | NW_017556795.1:161510..161612: − b |
| asp-miR-210-3p asp-miR-210-5p | NW_017550873.1_12017 | 4.8 | 36.5 | 0.4 | dme-miR-210-3p | CUUGUGCGUGUGACAGCGGC ....................UAU | NW_017550873.1:911604..911762: + b |
| asp-miR-1000-5p asp-miR-1000-3p | NW_017547246.1_6234 | 5 | 32.0 | 0.9 | dme-miR-1000-5p | AUAUUGUCCUGUCACAGCAG ....................U | NW_017547246.1:2709831..2709911: + b |
| asp-miR-929-5p asp-miR-929-3p | NW_017547330.1_8238 | 5 | 25.6 | 0.2 | dme-miR-929-5p | AAAUUGACUCUAGUAGGGAG ....................UC | NW_017547330.1:415939..415997: + |
| asp-miR-137-3p asp-miR-137-5p | NW_017547181.1_4201 | 73 | 25.1 | 0.4 | dme-miR-137-3p | UAUUGCUUGAGAAUACACGUA .....................G | NW_017547181.1:4906197..4906365: + b |
| asp-miR-29b-3p asp-miR-29b-5p | NW_017547261.1_6573 | 4.3 | 23.6 | 0 | aca-miR-29b | UAGCACCAUUUGAAAUCAGUG ..................... | NW_017547261.1:4373495..4373551: + |
| NW_017548020.1_10048 | NW_017548020.1:5969402..5969450: − | ||||||
| asp-miR-971b-5p asp-miR-971b-3p | NW_017547177.1_3772 | 72 | 21.6 | 2.8 | tca-miR-971b-5p | CACUCUAAGCUCGAACAUCAAG .........U.U..........C | NW_017547177.1:2847368..2847428: + |
| asp-miR-307a-3p asp-miR-307a-5p | NW_017547096.1_364 | 4.8 | 20.7 | 0 | dme-miR-307a-3p | UCACAACCUCCUUGAGUGAG ....................CGA | NW_017547096.1:1789984..1790139: − b |
| asp-miR-3049-3p asp-miR-3049-5p | NW_017547142.1_3035 | 69 | 16.2 | 7.0 | ame-miR-3049-3p | UCCGUCCAACUCUUUUCCGCC ............C......U.U | NW_017547142.1:3651122..3651182: + |
| asp-miR-2796-3p asp-miR-2796-5p | NW_017547276.1_6782 | 50 | 15.8 | 0.7 | ame-miR-2796-3p | GUAGGCCGGCGGAAACUACUUG ......................C | NW_017547276.1:164674..164899: − b |
| asp-miR-998-3p asp-miR-998-5p | NW_017547112.1_1536 | 5.3 | 14.5 | 0 | api-miR-998 | UAGCACCAUGGAAUUCAGCUU ....................G | NW_017547112.1:1633093..1633150: − |
| asp-miR-1175-3p asp-miR-1175-5p | NW_017547443.1_8882 | 3.3 | 13.5 | 0 | dme-miR-958-3p | UGAGAUUCAACUCCUCCAUC .................... | NW_017547443.1:363797..363852: − |
| asp-miR-965-3p asp-miR-965-5p | NW_017551483.1_12566 | 4.4 | 10.7 | 3.7 | dme-miR-965-3p | UAAGCGUAUAGCUUUUCCCC ....................UU | NW_017551483.1:4663002..4663057: − |
| asp-miR-iab-4-5p asp-miR-iab-4-3p | NW_017549609.1_11445 | 4.7 | 4.7 | 0.7 | dme-miR-iab-4-5p | ACGUAUACUAAAUGUAUCCU .........G..........GA | NW_017549609.1:2987912..2987972: − |
| asp-miR-iab-8-3p asp-miR-iab-8-5p | NW_017549609.1_11368 | 1.5 | 0.4 | 0.6 | dps-miR-iab-8-3p | AGGAUACAUUUAGUAUACGUC ..........C.........AUA | NW_017549609.1:2987914..2987973: + |
| Provisional miRNA Name (Mature/Star) | miRDeep2 ID | miRDeep2 Score | Read Count (RPM) | Example of miRBase miRNA with the Same Seed | Alignment between A. spiniferus miRNA (up) and miRbase miRNA (Bottom) | Precursor Coordinate in the Bemisia tabaci Genome | |
|---|---|---|---|---|---|---|---|
| Mature | Star | ||||||
| asp-miR-111690-5p asp-miR-111690-3p | NW_017549178.1_11169 | 2.2 × 102 | 68.9 | 9.0 | efu-mir-9235b | UGUGAUGUGCCUGUGGGCUUU UAAU.U.......G.......AAG | NW_017549178.1:3495258..3495320: − |
| asp-miR-131950-5p asp-miR-131950-3p | NW_017552654.1_13195 | 1 × 102 | 29.6 | 6.8 | dme-miR-304-5p | UAAUCUCAUACAGUUAUCCAUGA ..........UUUG..AAUG...G | NW_017552654.1:189212..189276: − |
| asp-miR-73390-3p asp-miR-73390-5p | NW_017547290.1_7339 | 11 | 2.0 | 0.6 | rno-miR-3590-3p | UGUCACAUUCGAAAAGAGCUG UA.CACA..GU..........C | NW_017547290.1:2474877..2474936: + |
| asp-miR-131930-5p asp-miR-131930-3p | NW_017552654.1_13193 | 4.5 | 1467.2 | 22.2 | dme-miR-12-5p | UGAGUAUUACAUCAGAUAUUU ...............G..C.GGU | NW_017552654.1:186778..186837: − |
| asp-miR-81360-5p asp-miR-81360-3p | NW_017547320.1_8136 | 4.3 | 405.1 | 0 | dme-miR-9a-5p | UCUUUGGUAUUCUAGCUGUAG ........UA..........UGA | NW_017547320.1:2154975..2155034: + |
| asp-miR-36450-5p asp-miR-36450-3p | NW_017547174.1_3645 | 5.2 | 343.1 | 0 | dme-miR-5-5p | UAAGGAACUAGGAAUGAGAUG A.......G.UCGU..U...AUG | NW_017547174.1:1937885..1937946: − |
| asp-miR-129380-3p asp-miR-129380-5p | NW_017552322.1_12938 | 5 | 65.8 | 0 | dme-miR-981-3p | UUCGUUGUCGCCGAAAACUCGUC ..........A.....C..GCA | NW_017552322.1:646402..646467: − |
| asp-miR-38410-3p asp-miR-38410-5p | NW_017547177.1_3841 | 2.8 | 46.9 | 0 | zma-miR396e-3p | ACUUUGGCCGUGAAAGCCGUG ..U.AA..........GGAAG | NW_017547177.1:5603872..5603954: + |
| asp-miR-161110-3p asp-miR-161110-5p | NW_017564655.1_16111 | 5.2 | 36.5 | 0 | api-miR-3016 | UUUGGUAAAAGAUAGCCGGUA A.......C.C.....UCU.UAG | NW_017564655.1:2288547..2288606: + |
| asp-miR-1011900-3p asp-miR-1011900-5p | NW_017548094.1_10119 | 5.1 | 33.3 | 0 | dme-miR-993-3p | CAAGCUCGUUGAAGUAUACCC G........CUCUAC.GGUAUCU | NW_017548094.1:36089..36137: + |
| asp-miR-64150-3p asp-miR-64150-5p | NW_017547257.1_6415 | 1.3 a | 26.7 | 0 | bmo-miR-3322 | CCUCGUCGGCGUCGGCGGC U........UU.GAAGUUGGCUAG | NW_017547257.1:1451280..1451335: + |
| asp-miR-22650-3p asp-miR-22650-5p | NW_017547116.1_2265 | 0.4 a | 11.8 | 0 | ame-miR-9883-5p | GUCGGGCGUAGUUAGUACU U.......GGC.CG.GCGAGA | NW_017547116.1:7925742..7925790: + |
| NW_017548020.1_10048 | 0 | 0 | NW_017548020.1:5969402..5969450: − | ||||
| NW_017547090.1_19 | 0 a | 21.8 | 0 | GUCGGGCGUAGUUAGUACUUAGAU U.......GGC.CG.GCGAGA | NW_017547090.1:1477845..1477898: + | ||
| asp-miR-33410-5p asp-miR-33410-3p | NW_017547161.1_3341 | 3.2 | 18.9 | 0 | mmu-miR-6976-5p | CAGGGAAUUUUUGACCAAAA .......G..GA..GG....UUG | NW_017547161.1:1252883..1252946: − |
| asp-miR-161130-3p asp-miR-161130-5p | NW_017564655.1_16113 | 5 | 13.5 | 0 | dme-miR-9c-3p | ACUUUGGUAAAAACAGCUGUG UAAAG....UA..CC..AGCUC | NW_017564655.1:2292773..2292828: + |
| asp-miR-144320-5p asp-miR-144320-3p | NW_017556720.1_14432 | 5.8 | 12.5 | 0 | eca-miR-9128 | ACUCGCGCGGGCACCGUCGU CUGGUG......U....CA....AC | NW_017556720.1:451897..451955: + |
| NW_017549581.1_11314 | 0 | 0 | NW_017549581.1:39602..39660: + | ||||
| NW_017555149.1_13938 | 0 | 0 | NW_017555149.1:1072638..1072696: + | ||||
| asp-miR-169840-3p asp-miR-169840-5p | NW_017566817.1_16984 | 3.6 | 7.5 | 0 | eca-miR-9047 | AGUUCCCGGGUUUCGGCACC U........C.GA.CCUGG.UAGG | NW_017566817.1:2966919..2966963: − |
| asp-miR-81390-3p asp-miR-81390-5p | NW_017547320.1_8139 | 4.7 | 5.3 | 0 | dme-miR-4-3p | AUAAAGCUGGAUUACCAAAGC ........A..CA....UU.A | NW_017547320.1:2163341..2163400: + |
| asp-miR-147030-3p asp-miR-147030-5p | NW_017557621.1_14703 | 0.8 a | 5.3 | 0 | mmu-miR-7648-3p | CGGCGCGGCCCGGGCGGCGG A...UG........AC....G | NW_017557621.1:10736677..10736742: + |
| asp-miR-1850-5p asp-miR-1850-3p | NW_017547093.1_185 | 1.4 a | 4.7 | 0 | hbv-miR-B4 | UCGGGGCGGCCGUUGCCG A.......UGGU.G.UG.GCG | NW_017547093.1:1632977..1633017: − |
| asp-miR-129190-3p asp-miR-129190-5p | NW_017552322.1_12919 | 3.8 | 4.4 | 0 | mdv1-miR-M10-3p | UCCCGGAGAAAUUUCGAGCU ..U..U...G.......UAACA | NW_017552322.1:437934..437995: + |
| asp-miR-29250-5p asp-miR-29250-3p | NW_017547136.1_2925 | 0 a | 4.4 | 0 | mdo-miR-7380-5p | AGGUCCCAGGUUCGAUCCCU .........C.CUAC...UCAU | NW_017547136.1:599807..599880: − |
| NW_017547136.1_2927 | 0 a | 0 | 0 | AGGUCCCAGGUUCGAUCCCU ...........CUAC...UCAU | NW_017547136.1:603764..603837: − | ||
| asp-miR-75080-3p asp-miR-75080-5p | NW_017547292.1_7508 | 0.4 a | 4.0 | 0 | gga-miR-6592-3p | CGCGUCUCCUCCCUCGGA A........CG...GACGGCC | NW_017547292.1:2958777..2958832: − |
| asp-miR-87590-3p asp-miR-87590-5p | NW_017547427.1_8759 | 5 | 3.8 | 0 | atr-miR8572 | CGACCCGGUCGGCGUCGG U..........UU.GAAUCCUCUC | NW_017547427.1:2779263..2779325: + |
| asp-miR-140930-5p asp-miR-140930-3p | NW_017555629.1_14093 | 0 a | 3.6 | 0 | dme-miR-959-5p | UUAGUACUUAGAUGGGAGAC U.......CG.G.U.AUA.AG | NW_017555629.1:127516..127600: − |
| asp-miR-151900-3p asp-miR-151900-5p | NW_017560566.1_15190 | 0.8 | 3.3 | 0 | bta-miR-11982 | AUUACCCGGCGCCUCCACC UU.............CUGCGGGU | NW_017560566.1:657..718: − |
| asp-miR-8520-5p asp-miR-8520-3p | NW_017547103.1_852 | 0.8 a | 3.1 | 0 | mmu-miR-3081-5p | GGCCGCGAGCGUGGAGUGGUGA .A.UG....U.....C..... | NW_017547103.1:943997..944071: + |
| asp-miR-35640-5p asp-miR-35640-3p | NW_017547174.1_3564 | 5 | 2.9 | 0 | dme-miR-5-5p | UAAGGAACUGUUUGAUGUGGU A.......GA.CGUUGUGAUAUG | NW_017547174.1:1945099..1945157: + |
| asp-miR-70610-3p asp-miR-70610-5p | NW_017547282.1_7061 | 3.3 | 2.9 | 0 | ame-miR-750-3p | CAGCUCUAACUCUUAAAGCUG .CAGA..........CCAG.C | NW_017547282.1:4449832..4449878: + |
| asp-miR-29460-5p asp-miR-29460-3p | NW_017547139.1_2946 | 1.2 | 2.9 | 0 | ame-bantam-3p | AGAUCCUUGUGACAGCUU UG.....A......A....GAUU | NW_017547139.1:273731..273785: + |
| asp-miR-46090-5p asp-miR-46090-3p | NW_017547184.1_4609 | 4.2 | 2.4 | 0 | dvi-miR-9709-5p | UUGCACUUCGGUAUCUCCAACCUAU ..........C.U.U.UAU..AACG | NW_017547184.1:3061811..3061874: − |
| asp-miR-109-5p asp-miR-42860-3p | NW_017547182.1_4286 | 3.9 | 2.4 | 0 | gsa-miR-10b-5p | UGUAGACCUGAAUCA AACCC........C....UUGA | NW_017547182.1:1044722..1044786: + |
| asp-miR-49480-3p asp-miR-49480-5p | NW_017547098.1_445 | 1.9 a | 2.4 | 0 | dme-miR-4948-5p | CGGCGGGCCGGGCGGCGU ........GU....UGCGUGAU | NW_017547098.1:4979831..4979950: + |
| asp-miR-125030-5p asp-miR-125030-3p | NW_017551483.1_12503 | 0.9 | 2.2 | 0 | efu-miR-9212 | UUCGCAUGGAGUCAUCUGUUAU ACA........GG.....GAGA | NW_017551483.1:448169..448229: − |
| asp-miR-165650-5p asp-miR-165650-3p | NW_017566689.1_16565 | 2.9 | 2.0 | 0 | hsa-miR-10523-5p | GACAAUGAUGACAAAAUUUGGU ...........G..G.CC..AGGA | NW_017566689.1:494135..494184: − |
| asp-miR-54140-5p asp-miR-54140-3p | NW_017547211.1_5414 | 4.6 | 1.8 | 0 | hsa-miR-1538 | GGGCCCGGGGUUCGAUUCC C........C.G.UGC.GUUCC | NW_017547211.1:1082752..1082825: + |
| asp-miR-1077800-3p asp-miR-1077800-5p | NW_017549038.1_10778 | 4.4 | 1.5 | 0.4 | pxy-miR-8536b-3p | AGGUAUCUGAGCGAUGUCCCCAC CC.....C...U.....A.U. | NW_017549038.1:8023737..8023796: + |
| NW_017551711.1_12713 | 0 | 0 | NW_017551711.1:428822..428883: − | ||||
| asp-miR-131970-5p asp-miR-131970-3p | NW_017552654.1_13197 | 1.7 | 1.3 | 0 | dme-miR-283-5p | AAAUAUCAGCUAGAUGCCUG ...........G.UAAUUCUGG | NW_017552654.1:190394..190458: − |
| asp-miR-125790-3p asp-miR-125790-5p | NW_017551561.1_12579 | 2.1 | 0.9 | 0 | aca-miR-9573-3p | CAAAACUAGAAUUGCCUAGG .........G....U...U. | NW_017551561.1:47313..47366: + |
| NW_017549024.1_10631 | 1.9 | 0.9 | 0 | NW_017549024.1:233711..233804: + | |||
| asp-miR-56900-5p asp-miR-56900-3p | NW_017547229.1_5690 | 0.3 | 0.9 | 0 | hsa-miR-483-5p | AUAAGACUGGAGGAAUGUAG .....C.......A.G...GAG | NW_017547229.1:2771708..2771774: + |
| asp-miR-77520-5p asp-miR-77520-3p | NW_017547298.1_7752 | 1.8 | 0.7 | 0 | sfr-miR-10458-5p | CACUGGAUACAUGUUAAACG GC.G......G....U.C.. | NW_017547298.1:3903205..3903268: + |
| asp-miR-147690-5p asp-miR-147690-3p | NW_017557621.1_14769 | 0 | 0.7 | 0 | cre-miR1171 | AGGAGUGGAUUUUCGAAC U........G.GGA.UGGAGUGG | NW_017557621.1:3670592..3670638: − |
| asp-miR-102140-5p asp-miR-102140-3p | NW_017548287.1_10214 | 0.5 | 0.5 | 0 | ppc-miR-2263 | GUCUCGGGUUCGAUUCCCG ........C.UCU..GAG.AU | NW_017548287.1:869888..869958: + |
| asp-miR-64380-3p asp-miR-64380-5p | NW_017547257.1_6438 | 0.7 | 0.4 | 0.4 | spu-miR-4848a | AGGGUUGAGUAAGGACAUCAAC U........GCUUUUGGG..GGA | NW_017547257.1:3219088..3219147: + |
| asp-miR-134420-3p asp-miR-134420-5p | NW_017553232.1_13442 | 0.9 | 0.2 | 0 | bta-miR-2489 | AAGGGCUCAUGAGUUUUUG AAAUG.C....GA......... | NW_017553232.1:988520..988580: − |
| NW_017553232.1_13405 | 0.9 | 0.2 | 0 | NW_017553232.1:988522..988582: + | |||
| asp-miR-98810-5p asp-miR-98810-3p | NW_017548003.1_9881 | 0.4 | 0.2 | 0 | efu-miR-9304 | UAAGCCCCCCUCCCCUCCC G.......UUGAAGGG..AGGGG | NW_017548003.1:5351436..5351483: − |
| asp-miR-33140-5p asp-miR-33140-3p | NW_017547161.1_3314 | 0.1 | 0.2 | 0 | bta-miR-2460 | GAAACUCAUGAGCCCUGGC U..G...U....G......AU | NW_017547161.1:6988319..6988379: + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bubici, G.; Prigigallo, M.I.; Garganese, F.; Nugnes, F.; Jansen, M.; Porcelli, F. First Report of Aleurocanthus spiniferus on Ailanthus altissima: Profiling of the Insect Microbiome and MicroRNAs. Insects 2020, 11, 161. https://doi.org/10.3390/insects11030161
Bubici G, Prigigallo MI, Garganese F, Nugnes F, Jansen M, Porcelli F. First Report of Aleurocanthus spiniferus on Ailanthus altissima: Profiling of the Insect Microbiome and MicroRNAs. Insects. 2020; 11(3):161. https://doi.org/10.3390/insects11030161
Chicago/Turabian StyleBubici, Giovanni, Maria Isabella Prigigallo, Francesca Garganese, Francesco Nugnes, Maurice Jansen, and Francesco Porcelli. 2020. "First Report of Aleurocanthus spiniferus on Ailanthus altissima: Profiling of the Insect Microbiome and MicroRNAs" Insects 11, no. 3: 161. https://doi.org/10.3390/insects11030161