Anion Inhibition Studies of the β-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
- (i)
- The following anions did not significantly inhibit CAS3 up to a concentration of 100 mM in the assay system: the halides (except iodide); sulfate, hydrogensulfite and peroxydisulfate; diphosphate and divanadate; perchlorate, tetrafluoroborate, fluorosulfonate and iminodisulfonate (Table 2). Some of the anions, such as perchlorate and tetrafluoroborate, are known to possess a low affinity for binding metal ions, both in solution and within the metalloenzyme active sites [4]. However, the halides generally inhibit CAs, as seen in Table 2 in which millimolar inhibition was reported for CAS1 and hCA II (except fluoride).
- (ii)
- Most of the investigated anions were low millimolar, weak CAS3 inhibitors, with inhibition constants in the range of 3.2–9.9 mM. They include iodide, the pseudohalides, bicarbonate, carbonate, nitrate, nitrite, hydrogensulfide, stannate, selenate, tellurate, tetraborate, perrhenate, perruthenate, selenocyanide and trithiocarbonate.
- (iii)
- Submillimolar inhibition was observed with diethylditiocarbamate (KI of 0.89 mM), as well as sulfamide, sulfanate, phenyl boronic and phenyl arsonic acids (KIs of 91–97 µM). As expected, the weak inhibitor trithiocarbonate (KI of 8.6 mM) led to an order of magnitude more effective inhibitor when the diethylamino fragment was introduced, with N,N-diethyl-ditiocarbamate showing a KI of 0.89 mM (Table 2).
- (iv)
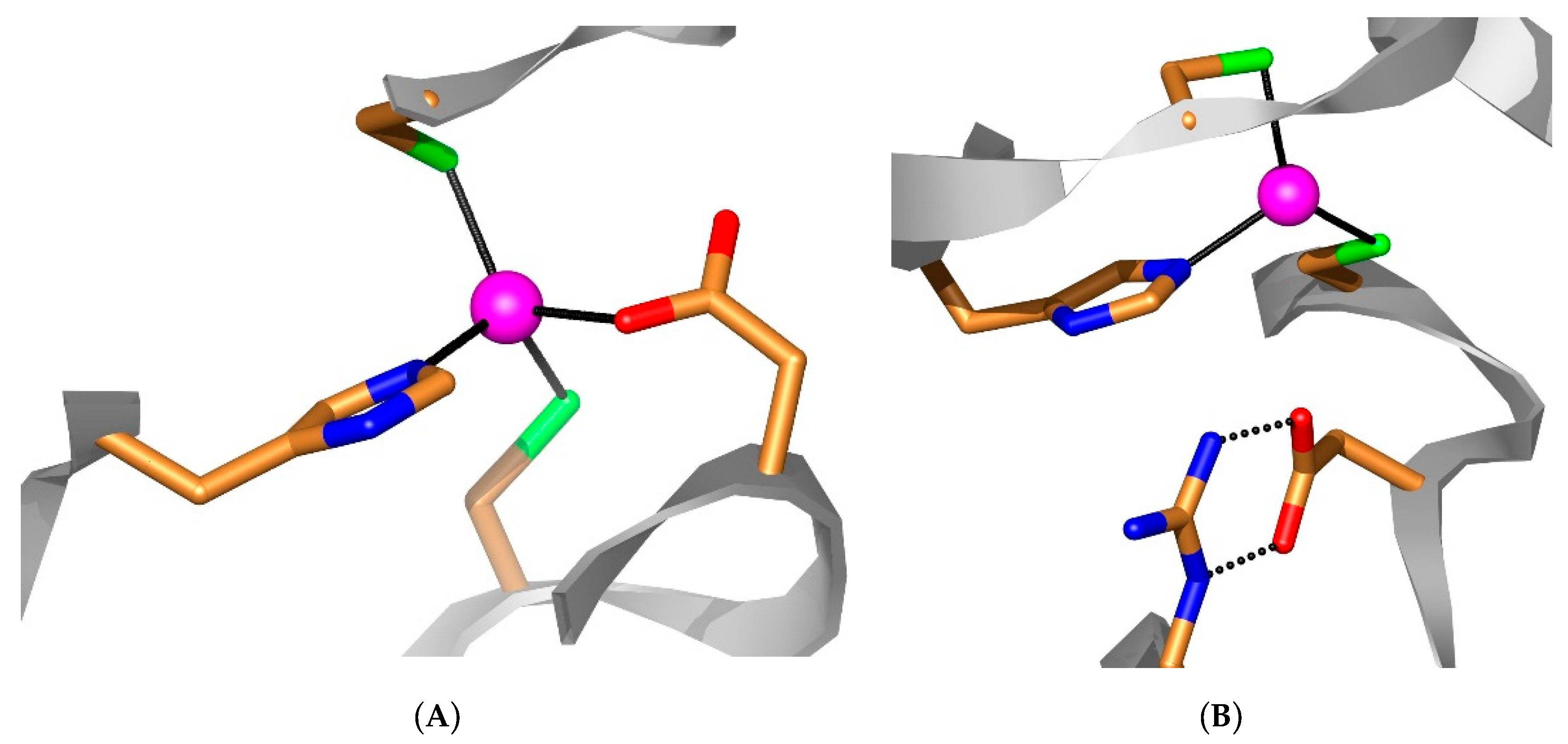
- The inhibition profile of CAS3 is rather different from that of the related isoforms, CAS1 and CAS2, and also from hCA II. However, no CAS3-selective inhibitors have been detected so far among these simple anions and small molecules. On the other hand, the identification of submillimolar or high micromolar inhibitors (diethyl-ditiocarbamate, sulfamide, sulfamate, phenyl boronic and phenyl arsonic acids) shows that it is probably possible to design compounds with an enhanced activity. In fact, acetazolamide (5-acetamido-1,3,4-thiadiazole-2-sulfonamide), which incorporates the sulfonamide fragment present in sulfamide and sulfamic acid, is a nanomolar CAS3 inhibitor (Table 1).
3. Materials and Methods
CA Activity and Inhibition Measurements
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fukuda, Y.; Tse, K.M.; Nakane, T.; Nakatsu, T.; Suzuki, M.; Sugahara, M.; Inoue, S.; Masuda, T.; Yumoto, F.; Matsugaki, N. Redox-coupled proton transfer mechanism in nitrite reductase revealed by femtosecond crystallography. Proc. Natl. Acad. Sci. USA 2016, 113, 2928–2933. [Google Scholar] [CrossRef] [Green Version]
- Schaffner, I.; Hofbauer, S.; Krutzler, M.; Pirker, K.F.; Bellei, M.; Stadlmayr, G.; Mlynek, G.; Djinovic-Carugo, K.; Battistuzzi, G.; Furtmüller, P.G.; et al. Dimeric chlorite dismutase from the nitrogen-fixing cyanobacterium Cyanothece sp. PCC7425. Mol Microbiol. 2015, 96, 1053–1068. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, A.; Ponnuraj, K. Crystal structure of the first plant urease from jack bean: 83 years of journey from its first crystal to molecular structure. J. Mol. Biol. 2010, 400, 274–283. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Supuran, C.T. (In)organic anions as carbonic anhydrase inhibitors. J. Inorg. Biochem. 2012, 111, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Winum, J.Y. Introduction to zinc enzymes as drug targets. In Drug Design of Zinc-Enzyme Inhibitors: Functional, Structural, and Disease Applications; Supuran, C.T., Winum, J.Y., Eds.; Wiley: Hoboken, NJ, USA, 2009; pp. 3–12. [Google Scholar]
- Temperini, C.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. X-Ray crystal studies of the carbonic anhydrase II—Trithiocarbonate adduct—An inhibitor mimicking the sulfonamide and urea binding to the enzyme. Bioorg. Med. Chem. Lett. 2010, 20, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Supuran, C.T. Dithiocarbamates: A new class of carbonic anhydrase inhibitors. Crystallographic and kinetic investigations. Chem. Commun. 2012, 48, 1868–1870. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Durante, M.; Di Leva, F.S.; Cosconati, S.; Masini, E.; Scozzafava, A.; Novellino, E.; Supuran, C.T.; Carta, F. Monothiocarbamates strongly inhibit carbonic anhydrases in vitro and possess intraocular pressure lowering activity in an animal model of glaucoma. J. Med. Chem. 2016, 59, 5857–5867. [Google Scholar] [CrossRef]
- Carta, F.; Akdemir, A.; Scozzafava, A.; Masini, E.; Supuran, C.T. Xanthates and trithiocarbonates strongly inhibit carbonic anhydrases and show antiglaucoma effects in vivo. J. Med. Chem. 2013, 56, 4691–4700. [Google Scholar] [CrossRef]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Masini, E.; Supuran, C.T. Dithiocarbamates strongly inhibit carbonic anhydrases and show antiglaucoma action in vivo. J. Med. Chem. 2012, 55, 1721–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspatwar, A.; Hammarén, M.; Koskinen, S.; Luukinen, B.; Barker, H.; Carta, F.; Supuran, C.T.; Parikka, M.; Parkkila, S. β-CA-specific inhibitor dithiocarbamate Fc14-584B: A novel antimycobacterial agent with potential to treat drug-resistant tuberculosis. J. Enzyme Inhib. Med. Chem. 2017, 32, 832–840. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, G.T.H.; Tran, T.N.; Podgorski, M.N.; Bell, S.G.; Supuran, C.T.; Donald, W.A. Nanoscale Ion Emitters in Native Mass Spectrometry for Measuring Ligand-Protein Binding Affinities. ACS Cent. Sci. 2019, 5, 308–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, A.B.; Aggarwal, M.; Pinard, M.; Vullo, D.; Patrauchan, M.; Supuran, C.T.; McKenna, R. Structural Mapping of Anion Inhibitors to β-Carbonic Anhydrase psCA3 from Pseudomonas aeruginosa. ChemMedChem 2018, 13, 2024–2029. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Donald, W.A.; Winum, J.Y.; Supuran, C.T.; Capasso, C. Anion inhibitors of the β-carbonic anhydrase from the pathogenic bacterium responsible of tularemia, Francisella tularensis. Bioorg. Med. Chem. 2017, 25, 4800–4804. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. Carbonic Anhydrase from Porphyromonas gingivalis as a Drug Target. Pathogens 2017, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Legionella pneumophila Carbonic Anhydrases: Underexplored Antibacterial Drug Targets. Pathogens 2016, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Haapanen, S.; Bua, S.; Kuuslahti, M.; Parkkila, S.; Supuran, C.T. Cloning, Characterization and Anion Inhibition Studies of a β-Carbonic Anhydrase from the Pathogenic Protozoan Entamoeba histolytica. Molecules 2018, 23, 3112. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Inhibition of carbonic anhydrase from Trypanosoma cruzi for the management of Chagas disease: An underexplored therapeutic opportunity. Future Med. Chem. 2016, 8, 311–324. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. The η-class carbonic anhydrases as drug targets for antimalarial agents. Expert Opin. Ther. Targets 2015, 19, 551–563. [Google Scholar] [CrossRef]
- Del Prete, S.; Bua, S.; Zoccola, D.; Alasmary, F.A.S.; AlOthman, Z.; Alqahtani, L.S.; Techer, N.; Supuran, C.T.; Tambutté, S.; Capasso, C. Comparison of the Anion Inhibition Profiles of the α-CA Isoforms (SpiCA1, SpiCA2 and SpiCA3) from the Scleractinian Coral Stylophora pistillata. Int. J. Mol. Sci. 2018, 19, 2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prete, S.D.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Anion Inhibition Profile of the β-Carbonic Anhydrase from the Opportunist Pathogenic Fungus Malassezia Restricta Involved in Dandruff and Seborrheic Dermatitis. Metabolites 2019, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
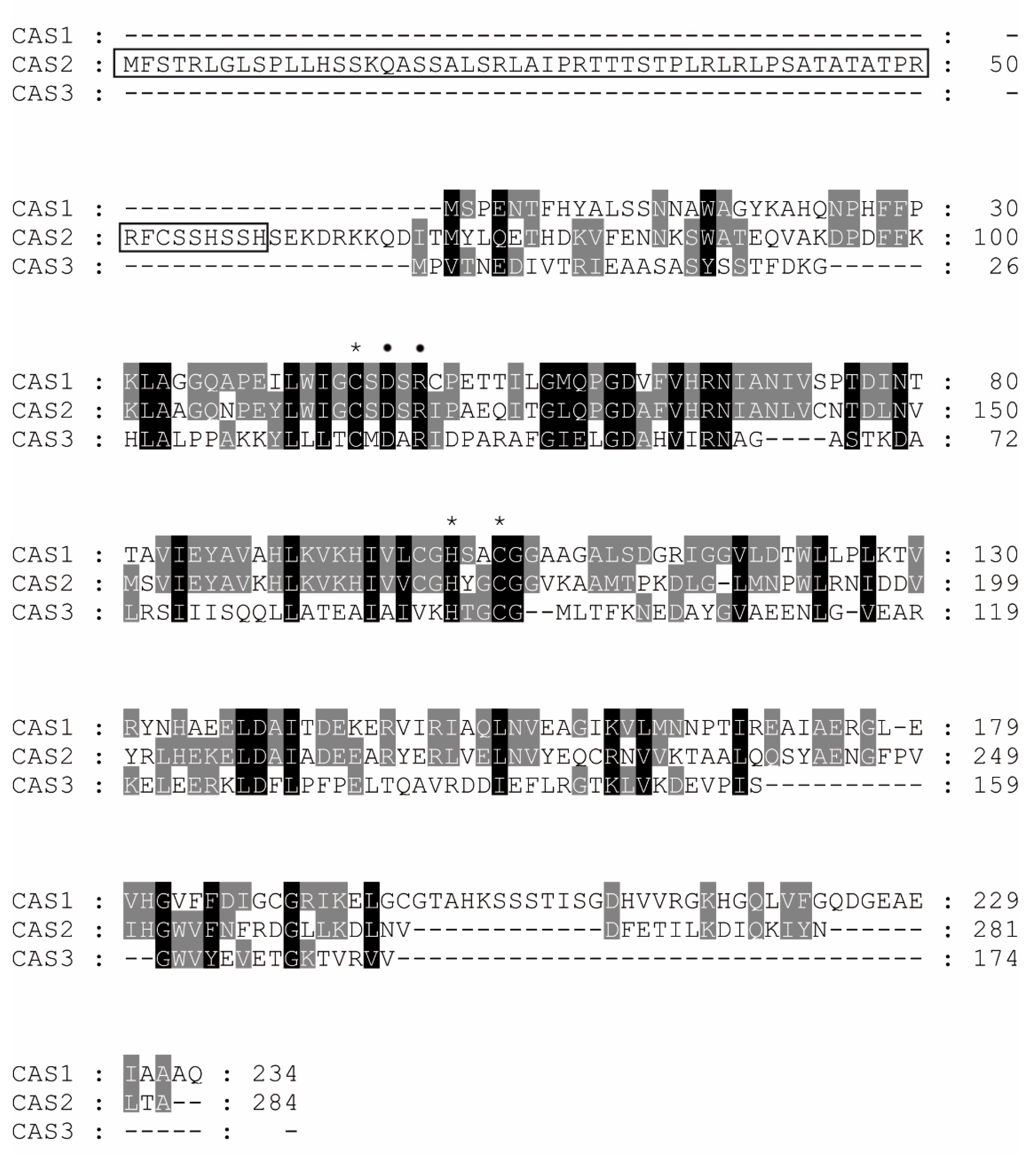
- Lehneck, R.; Neumann, P.; Vullo, D.; Elleuche, S.; Supuran, C.T.; Ficner, R.; Pöggeler, S. Crystal structures of two tetrameric β-carbonic anhydrases from the filamentous ascomycete Sordaria macrospora. FEBS J. 2014, 281, 1759–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innocenti, A.; Leewattanapasuk, W.; Mühlschlegel, F.A.; Mastrolorenzo, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzyme from the pathogenic yeast Candida glabrata with anions. Bioorg. Med. Chem. Lett. 2009, 19, 4802–4805. [Google Scholar] [CrossRef]
- Isik, S.; Kockar, F.; Arslan, O.; Guler, O.O.; Innocenti, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzyme from the yeast Saccharomyces cerevisiae with anions. Bioorg. Med. Chem. Lett. 2008, 18, 6327–6331. [Google Scholar] [CrossRef]
- Innocenti, A.; Mühlschlegel, F.A.; Hall, R.A.; Steegborn, C.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors: Inhibition of the beta-class enzymes from the fungal pathogens Candida albicans and Cryptococcus neoformans with simple anions. Bioorg. Med. Chem. Lett. 2008, 18, 5066–5070. [Google Scholar] [CrossRef]
- Innocenti, A.; Zimmerman, S.; Ferry, J.G.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzyme from the methanoarchaeon Methanobacterium thermoautotrophicum (cab) with anions. Bioorg. Med. Chem. Lett. 2004, 14, 4563–4567. [Google Scholar] [CrossRef]
- Nocentini, A.; Supuran, C.T. Advances in the structural annotation of human carbonic anhydrases and impact on future drug discovery. Expert Opin. Drug Discov. 2019, 14, 1175–1197. [Google Scholar] [CrossRef]
- Lehneck, R.; Pöggeler, S. Fungal carbonic anhydrases and their inhibition. In Zinc Enzyme Inhibitors—Volume 1: Enzymes from Microorganisms; Supuran, C.T., Capasso, C., Eds.; Topics in Medicinal Chemistry; Springer: Cham, Switzerland, 2017; Volume 22, pp. 95–110. [Google Scholar]
- Supuran, C.T.; Capasso, C. Biomedical applications of prokaryotic carbonic anhydrases. Expert Opin. Ther. Pat. 2018, 28, 745–754. [Google Scholar] [CrossRef]
- Annunziato, G.; Giovati, L.; Angeli, A.; Pavone, M.; Del Prete, S.; Pieroni, M.; Capasso, C.; Bruno, A.; Conti, S.; Magliani, W.; et al. Discovering a new class of antifungal agents that selectively inhibits microbial carbonic anhydrases. J. Enzyme Inhib. Med. Chem. 2018, 33, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Sulfonamide Inhibition Profile of the β-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions. Metabolites 2020, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vullo, D.; Lehneck, R.; Pöggeler, S.; Supuran, C.T. Sulfonamide inhibition studies of two β-carbonic anhydrases from the ascomycete fungus Sordaria macrospora, CAS1 and CAS2. J. Enzyme Inhib. Med. Chem. 2018, 33, 390–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitson, K.S.; Vullo, D.; Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Molecular cloning, characterization, and inhibition studies of a β-carbonic anhydrase from Malassezia globosa, a potential antidandruff target. J. Med. Chem. 2012, 55, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Teichert, I.; Nowrousian, M.; Pöggeler, S.; Kück, U. The filamentous fungus Sordaria macrospora as a genetic model to study fruiting body development. Adv. Genet. 2014, 87, 201–246. [Google Scholar]
- Elleuche, S.; Pöggeler, S. Evolution of carbonic anhydrases in fungi. Curr. Genet. 2009, 55, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Elleuche, S.; Pöggeler, S. β-Carbonic anhydrases play a role in fruiting body development and ascospore germination in the filamentous fungus Sordaria macrospora. PLoS ONE 2009, 4, e5177. [Google Scholar] [CrossRef] [Green Version]
- Elleuche, S.; Pöggeler, S. Carbonic anhydrases in fungi. Microbiology 2010, 156, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Lehneck, R.; Elleuche, S.; Pöggeler, S. The filamentous ascomycete Sordaria macrospora can survive in ambient air without carbonic anhydrases. Mol. Microbiol. 2014, 95, 931–944. [Google Scholar] [CrossRef]
- Schlicker, C.; Hall, R.A.; Vullo, D.; Middelhaufe, S.; Gertz, M.; Supuran, C.T.; Muhlschlegel, F.A.; Steegborn, C. Structure and inhibition of the CO2-sensing carbonic anhydrase Can2 from the pathogenic fungus Cryptococcus neoformans. J. Mol. Biol. 2009, 385, 1207–1220. [Google Scholar] [CrossRef]
- Diaz, J.R.; Fernández Baldo, M.; Echeverría, G.; Baldoni, H.; Vullo, D.; Soria, D.B.; Supuran, C.T.; Camí, G.E. A substituted sulfonamide and its Co (II), Cu (II), and Zn (II) complexes as potential antifungal agents. J. Enzyme Inhib. Med. Chem. 2016, 31, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [PubMed]
- Ferraroni, M.; Del Prete, S.; Vullo, D.; Capasso, C.; Supuran, C.T. Crystal structure and kinetic studies of a tetrameric type II β-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Acta Crystallogr. D Biol. Crystallogr. 2015, 71, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Suarez Covarrubias, A.; Larsson, A.M.; Hogbom, M.; Lindberg, J.; Bergfors, T.; Bjorkelid, C.; Mowbray, S.L.; Unge, T.; Jones, T.A. Structure and function of carbonic anhydrases from Mycobacterium tuberculosis. J. Biol. Chem. 2005, 280, 18782–18789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez Covarrubias, A.; Brgfors, T.; Jones, T.A.; Hogbom, M. Structural mechanics of the pH-dependent activity of the β-carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.S.; Ingram-Smith, C.; Ferry, J.G. Roles of the conserved aspartate and arginine in the catalytic mechanism of an archaeal beta-class carbonic anhydrase. J. Bacteriol. 2002, 184, 4240–4245. [Google Scholar] [CrossRef] [Green Version]
- Tufts, B.L.; Esbaugh, A.; Lund, S.G. Comparative physiology and molecular evolution of carbonic anhydrase in the erythrocytes of early vertebrates. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 136, 259–269. [Google Scholar] [CrossRef]
- Vullo, D.; Lehneck, R.; Donald, W.A.; Pöggeler, S.; Supuran, C.T. Sulfonamide Inhibition Studies of the β-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora. Molecules 2020, 25, 1036. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isozyme | Activity Level | kcat (s−1) | kcat/Km (M−1 × s−1) | KI (Acetazolamide) (nM) |
|---|---|---|---|---|
| hCA I a | moderate | 2.0 × 105 | 5.0 × 107 | 250 |
| hCA II a | very high | 1.4 × 106 | 1.5 × 108 | 12 |
| Can2 b | moderate | 3.9 × 105 | 4.3 × 107 | 10.5 |
| CalCAc | high | 8.0 × 105 | 9.7 × 107 | 132 |
| SceCA d | high | 9.4 × 105 | 9.8 × 107 | 82 |
| Cab e | low | 3.1 × 104 | 1.82 × 106 | 12,100 |
| CAS1 f | low | 1.2 × 104 | 1.30 × 106 | 445 |
| CAS2 f | low | 1.3 × 104 | 1.21 × 106 | 816 |
| CAS3 g | high | (7.9 ± 0.2) × 105 | (9.5 ± 0.12) × 107 | 94 ± 3 |
| Inhibitor § | hCA II a | KI [mM] # CAS1 b | CAS2 b | CAS3 c |
|---|---|---|---|---|
| F- | >300 | >100 | >100 | >100 |
| Cl- | 200 | 9.2 | >100 | >100 |
| Br- | 63 | 9.3 | >100 | >100 |
| I- | 26 | 8.6 | 7.7 | 9.9 |
| CNO- | 0.03 | 0.9 | 0.82 | 3.2 |
| SCN- | 1.6 | 5.4 | 5.6 | 7.3 |
| CN- | 0.02 | 0.94 | 0.75 | 8.7 |
| N3- | 1.51 | >100 | 6.1 | 7.2 |
| HCO3- | 85 | 6.5 | 5.5 | 3.4 |
| CO32- | 73 | >100 | 8.8 | 8 |
| NO3- | 35 | >100 | >100 | 8.5 |
| NO2- | 63 | >100 | >100 | 8.3 |
| HS- | 0.04 | 0.89 | 8.5 | 8.3 |
| HSO3- | 89 | 3.3 | 7.3 | >100 |
| SO42- | >200 | >100 | 4.8 | >100 |
| SnO32- | 0.83 | 4.3 | 0.92 | 7.9 |
| SeO42- | 112 | 2.4 | 9.2 | 3.4 |
| TeO42- | 0.92 | 2.5 | 6.3 | 8.1 |
| P2O74- | 48.5 | 3.1 | 0.96 | >100 |
| V2O74- | 0.57 | >100 | 1.4 | >100 |
| B4O72- | 0.95 | 6.7 | 6.9 | 5.9 |
| ReO4- | 0.75 | 8.2 | >100 | 8.8 |
| RuO4- | 0.69 | 3.9 | >100 | 9.2 |
| S2O82- | 0.084 | 5 | >100 | >100 |
| SeCN- | 0.086 | 2.9 | 9.3 | 7.1 |
| CS32- | 0.0088 | 0.79 | >100 | 8.6 |
| Et2NCS2- | 3.1 | 0.38 | 0.93 | 0.89 |
| ClO4- | >200 | > 100 | > 100 | >100 |
| BF4- | >200 | > 100 | > 100 | >100 |
| FSO3- | 0.46 | 0.93 | 8.4 | >100 |
| NH(SO3)22- | 0.76 | 0.88 | 9.2 | >100 |
| H2NSO2NH2 | 1.13 | 0.084 | 0.048 | 0.094 |
| H2NSO3H | 0.39 | 0.069 | 0.072 | 0.095 |
| Ph-B(OH)2 | 23.1 | 0.009 | 0.056 | 0.097 |
| Ph-AsO3H2 | 49.2 | 0.035 | 0.054 | 0.091 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vullo, D.; Lehneck, R.; Donald, W.A.; Pöggeler, S.; Supuran, C.T. Anion Inhibition Studies of the β-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora. Metabolites 2020, 10, 93. https://doi.org/10.3390/metabo10030093
Vullo D, Lehneck R, Donald WA, Pöggeler S, Supuran CT. Anion Inhibition Studies of the β-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora. Metabolites. 2020; 10(3):93. https://doi.org/10.3390/metabo10030093
Chicago/Turabian StyleVullo, Daniela, Ronny Lehneck, William A. Donald, Stefanie Pöggeler, and Claudiu T. Supuran. 2020. "Anion Inhibition Studies of the β-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora" Metabolites 10, no. 3: 93. https://doi.org/10.3390/metabo10030093