
Abstract
Although the role of autophagy has been implicated in several forms of chronic hepatitis, it is still not fully understood. Active autophagy eliminates damaged molecules and organelles (such as mitochondria) by lysosomal degradation. In the present study, we aimed to examine and compare autophagy activity in chronic hepatitis C (CHC) and autoimmune hepatitis (AIH) by detecting the expression of autophagy (LC3 and p62) and mitochondrium-related (TOMM20) proteins, as well as the levels of selected microRNAs (miR-101, -155, -204 and − 224) known to be involved in the regulation of autophagy. In addition, the expression levels were related to pathohistological parameters. Liver biopsy samples, including 45 CHC and 18 AIH cases, were immunohistochemically stained for LC3, p62 and TOMM20 and the expression of miRNAs was determined using real-time PCR. We found elevated LC3 and p62 in AIH samples as compared with CHC ones, indicating an activated autophagy that is impaired in AIH as no degradation of p62 seemed to occur. Moreover, p62 showed strong correlation with necroinflammatory grades in the AIH group. The observed elevated levels of TOMM20 and p62 suggest a less efficient elimination of damaged mitochondria in AIH as opposed to CHC, in which autophagy seems to have a more active function. The level of miR-101 was increased in case of CHC as compared with AIH, however, miR-155, -204 and 224 resulted in no expressional. Furthermore, miR-224 level correlated with steatosis and miR-155 expression with fibrosis stage in CHC. In conclusion, dissimilar autophagic activity was observed in CHC and AIH, suggesting a close association between impaired autophagy and severity of necroinflammation. This impairment may not be regulated by the analyzed miRNAs. Nevertheless, miR-224 and − 155 seem to be associated with CHC progression.
Similar content being viewed by others

Introduction
Hepatitis C virus (HCV) infection is one of the main health problems affecting 71 million people worldwide [1]. Despite the remarkable healing effect of the newly introduced direct-acting antivirals (DAAs), several issues are still not resolved or are under dispute [2, 3]. One of these is the role autophagy may play in the pathogenesis of chronic hepatitis C (CHC), which has been in the focus of several recent studies [4,5,6].
Autophagy, this evolutionarily conserved cellular catabolic process, is responsible for the elimination of predominantly damaged molecules and organelles via delivering them to lysosomes for degradation. Cell growth, development and homeostasis are the major processes in which autophagy is involved in order to maintain the balance between synthesis, degradation, and subsequent recycling of cellular products [7]. Several stress factors, including viral infection and autoimmunity, are known to activate autophagy [4, 8, 9]. Selective mitochondrial autophagy, termed mitophagy is particularly important in the elimination of mitochondrial dysfunction arising from aging and damage, thus, mitophagy plays crucial role in the reduction of oxidative stress and maintenance of cellular homeostasis [4, 6, 10,11,12].
An activated autophagy process is described as being beneficial for the replication and life cycle of HCV, since it protects the persistence of the virus in multiple ways. As it is known, intracellular membraneous web-associated virus replication and production of structural and non-structural HCV proteins lead to endoplasmic reticulum (ER)-stress and unfolded protein response (UPR), by which HCV indirectly induces autophagy through the activation of the UPR-induced signaling pathway [8, 13, 14]. ER-stress causes mitochondrial injury and leads to mitophagy [13, 15] as demonstrated in HCV infection [10]. In addition, HCV-induced autophagy is capable of suppressing the innate immune response and thus interferon (IFN) production, too [3, 13]. All these maintain the persistence of HCV infection and the pathogenesis of CHC without causing direct cytopathic effect on hepatocytes [3].
Autoimmune hepatitis (AIH) is another form of chronic hepatitis with an unknown etiology. Activated immune cells, involving cytokines and macrophages, characterize the disease [16], however, overall knowledge about autophagy in AIH is limited. Autophagy is known to be involved in the activation of adaptive immunity, promoting the direct elimination of pathogens and simultaneous antigen processing and presentation to T cells [17]. Autophagy is therefore believed to be linked with the immune response seen in AIH, with involvement of several autophagy-related genes and various pathways via mechanisms that are still unresolved [9].
The process of autophagy involves formation of phagophores, closure of autophagosomes and fusion of lysosomes with autophagosomes, in which the sequestrated cargo is digested by acidic hydrolases [8, 13]. Proteins found to be indicative of autophagy include LC3 (microtubule-associated protein light chain 3α) and p62 (or SQSTM1, sequestosome-1). LC3 becomes associated with the newly formed phagophore and interacts with p62, leading to the closure of the phagophored cargo (autophagosome) [8, 13]. TOMM20 is a protein in the outer mitochondrial membrane, responsible for recognizing and sorting precursor proteins for the biogenesis of mitochondria. Its expression is indicative of the available mitochondria in the cell, either damaged or active, and may correlate with mitophagy [18,19,20].
Several microRNAs (miRNAs) have been implicated in the regulation of autophagy by interfering with gene expression at post-transcriptional level. For example, miR-155 induces the formation of autophagophores via downregulating mTOR signaling but it impairs lysosomal degradation, as shown in alcoholic liver disease (ALD) [21]. miR-101 is thought to be a key regulator of the autophagy process and is a potent inhibitor of rapamycin-induced autophagy [22, 23]. miR-204 attenuates tumor autophagy in hepatocellular carcinoma (HCC) [24]. Additionally, autophagy itself may also regulate the expression of certain miRNAs, for example, the degradation of miR-224 in HCC [25]. Relationship between miRNAs and autophagy may be important in the pathogenesis of liver diseases [22].
In light of the above, our aim was to compare the expression of autophagy and mitochondrium-related proteins, and the levels of selected miRNAs in liver biopsy samples taken from patients with CHC and AIH. Moreover, we related the data to steatosis, fibrosis stage and necroinflammatory grade of the chronic hepatitis samples.
Material and Methods
Patient Characteristics
Liver biopsy samples from patients with CHC (45 cases) and AIH (18 cases) were taken for diagnostic purposes and analysed retrospectively before any treatment had been started. All samples derived from the archives of the 2nd Department of Pathology, Semmelweis University, Budapest with approval from the National Ethics Committee (45727-2/2013/EKU). The study was performed according to the Declaration of Helsinki. Patients were between 18 and 73 years of age at the time of biopsy with average age of 48.8 years for CHC and 43.4 years for AIH cases. The CHC group consisted of 31 males and 14 females, the AIH group comprised 5 males and 13 females. Chronic hepatitis cases of mixed etiology (alcohol consumption, hepatitis B virus (HBV) or other viral infections, metabolic liver diseases, advanced cirrhosis, tumors, and drug treatments) were excluded.
Histology
Liver core biopsy samples taken with Menghini needle were fixed in formalin and embedded in paraffin (FFPE). Hematoxylin-eosin (H&E) stained sections of 3–4 µm thickness were analyzed to establish the diagnosis, necroinflammatory grade (according to the Ishak modification of the Knodell histology activity index [26]) and fibrosis stage (using the METAVIR scoring system [27]). Picrosyrius red staining was applied to highlight connective tissue fibers. Necroinflammatory grades were scored from 0 to 18 and fibrosis stages from F0 to F4. The degree of steatosis was assessed as percentage of lipid droplets-containing hepatocytes and scored 0 (under 5%), 1 (5–33%), 2 (34–66%) and 3 (above 66%) [28].
Immunohistochemistry
Immunohistochemical reactions were carried out on 3-4-µm-thick FFPE sections with Ventana Benchmark XT automated immunohistochemical staining system (Ventana Medical Systems; Tucson, AZ) using HRP multimer based, biotin-free detection technique according to the protocol provided by the manufacturers. Briefly, antigen retrieval for 30 minutes at 95 °C was followed by incubation with specific primary antibodies for 30 minutes at 42 °C. The primary antibodies were directed against LC3 (polyclonal rabbit, NB-100-2331, Novus Biologicals, Abingdon UK, dilution 1:200), p62 (monoclonal mouse, ab56416; AbCam Cambridge UK, dilution 1:1000) and TOMM20 (monoclonal mouse, F-10 sc-17764, Santa Cruz Biotechnology Inc., Oregon USA, dilution 1:200). Indirect biotin streptavidin system (iVIEW DAB Detection Kit, Ventana) was used for visualization of reactions. For negative controls, the specific primary antibodies were omitted and replaced by the antibody diluent. Cervical carcinoma tissues served as positive controls for LC3 and TOMM20, and alcohol-induced HCC samples for p62 as recommended by the manufacturers of the antibodies.
Evaluation of the immunohistochemical reactions was performed using semiquantitative analysis. Each reaction was given a score according to the intensity of staining (0 – no, 1 – minimal, 2 – weak, 3 – moderate, 4 – intensive and 5 – very intensive), as well as the degree of cell staining [(0) < 6%, (1) 6–20%, (2) 21–40%, (3) 41–60%, (4) 61–80% and (5) > 81%]. Finally, the scores for intensity and percentage were summarized (IHC score).
RNA Isolation
Total RNA was isolated from 5 µm thick FFPE sections using RNeasy FFPE Kit (QIAGEN, Venlo, The Netherlands) according to the instructions of the manufacturer with modifications for co-purification of miRNAs [29]. Traces of genomic DNA were eliminated by DNase digestion (Turbo DNA-free kit, Ambion by Life Technologies, Carlsbad, CA, USA). The concentration of RNA was measured using a NanoDrop 1000 spectrophotometer (Life Technologies of Thermo Fischer Scientific, Waltham, MA, USA).
Reverse Transcription (RT) and Quantitative Real Time PCR (qPCR)
The list of the 6 selected miRNAs is presented in Table 1. along with the ID codes for the TaqMan MicroRNA Assays (Life Technologies brand of Thermo Fisher Scientific Inc.) used for measuring the expression of these miRNAs as instructed by the manufacturer. RT reaction was carried out using TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystem brand by Thermo Fisher Scientific Inc.) in a total volume of 7.5 µl containing 20 ng RNA preparation. The qPCR was performed using TaqMan Universal Master Mix II no UNG (Applied Biosystem brand by Thermo Fisher Scientific Inc.) in a final volume of 7.5 µl containing 0.65 µl of the RT product. The amplification reaction was run in triplicates on a LightCycler 480 Instrument II (Roche Diagnostics, Indianapolis, IN). Relative expression was calculated by the 2∆Cq (where ΔCq = CqRef-CqmiR) formula, using the average of miR-140 and RNU6B as the most stable reference determined by NormFinder application [30].
Statistical Analysis
All figures and statistical analyses were created using GraphPad Prism (version 5.01; California, USA) and Statistic v.13 (Stat-Soft Inc.,Tulsa, OK, USA) software. A p-value of 0.05 was set as the threshold for statistical significance. The differences between CHC and AIH in relation to steatosis, fibrosis stage and necroinflammatory grade were examined by non-parametric Kruskal-Wallis analysis of variance and Mann-Whitney U test. Correlation of steatosis, fibrosis stage and necroinflammatory grade with IHC scores and miRNA expression was performed using non-parametric Spearman rank correlation test.
Results
Histology
The established pathohistological parameters for the samples are listed in Table 2. No steatosis was present in 12 (26.7%) samples with CHC and in 14 (77.8%) samples with AIH; whereas liver steatosis could be detected in 33 (73.3%) CHC cases and 4 (22.2%) AIH cases. Fibrosis stage was found to be low (F1-F2) in 31 (68.9%) CHC and 7 (38.9%) AIH cases, whereas high (F3-F4) in 14 (31.1%) CHC and 11 (61.1%) AIH cases. Finally, mild necroinflammation was diagnosed in 25 (55.6%) CHC cases, whereas 11 (61.1%) AIH samples showed severe necroinflammation.
Evaluation of Immunohistochemical Reactions
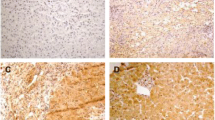
LC3 and p62 stainings were rather diffuse, occasionally showing granular-focal cytoplasmic reaction with varying density both in hepatocytes and cholangiocytes, while TOMM20 exhibited brownish granular cytoplasmic reaction (Fig. 1). Altogether 42 CHC and 18 AIH samples provided evaluable immunohistochemical reaction for LC3, p62 and TOMM20.

Representative images of low and high levels of LC3 (a, b), p62 (c, d) and TOMM20 (e, f) detected by immunohistochemistry (40x magnification).
Overall, AIH cases showed elevated LC3 (p < 0.01), p62 (p < 0.0001) and TOMM20 (p < 0.0001) levels as compared to CHC cases (Fig. 2a). When calculating ratios, the average LC3/TOMM20 ratios for the AIH and CHC groups were 0.76 and 0.80, respectively, while the average p62/TOMM20 ratio turned out to be 0.85 for the AIH and 0.26 for the CHC group.

Semiquantitative evaluation of LC3, p62 and TOMM20 immunohistochemical reactions (a) and their association with steatosis (b) and necroinflammatory grade (c) in CHC and AIH.
No differences in the IHC scores of these proteins were observed when cases with or without steatosis were compared in either CHC or AIH samples (Fig. 2b). Nevertheless, decreased p62 could be found in the steatotic group of CHC samples as compared with the nonsteatotic group, however, the difference did not reach the set threshold for significance. Further, no association could be observed between fibrosis stages and LC3, p62, TOMM20 levels.
In regard to necroinflammatory grade, the level of p62 was significantly elevated (p < 0.0001) in severe grades of AIH as compared to mild grades (Fig. 2c). In addition, p62 showed strong correlation with necroinflammatory grade (r = 0.84; p < 0.0001) in AIH cases (Table 3) but no correlation was detectable in CHC samples (r=-0.03; p = 0.8445). Moreover, no association was observed between the levels of LC3, TOMM20 and necroinflammatory grade in relation to the two etiological groups.
miRNA Expression
The levels of miRNAs could be determined in 24 CHC and 18 AIH samples. Expression of miR-224, -155 and − 204 was not statistically different between the CHC and AIH groups. On the contrary, an elevated level of miR-101 was observed in CHC as compared with AIH samples (p < 0.0001) (Fig. 3a).

Relative miRNA expression (a) and the association of miR-155 and − 101 levels with fibrosis stages (b) in CHC and AIH
No statistically different miRNA levels were found in either the CHC or AIH group when data were analyzed with regard to steatosis and non-steatosis cases. Nevertheless, the miR-224 level correlated with steatosis in CHC samples (r = 0.42; p < 0.05) (Table 3.) but not in AIH ones.
When miRNA expression was compared with the low (F1-F2), high (F3-F4) or separately with each (F1-F4) stage of fibrosis, no differences were found regarding miR-224, -101, -204 in either the CHC or AIH group or in case of miR-155 in the AIH group, but an elevated level of miR-155 (p < 0.05) was observed in the high as compared to the low stage of fibrosis in case of CHC (Fig. 3b). Furthermore, in the CHC group miR-155 level showed statistical difference between stages F1 and F4 (p < 0.05, data not shown) and miR-155 expression exhibited correlation with fibrosis stage as well (r = 0.56; p < 0.01) (Table 3.). Finally, no correlation was observable between miRNA expression and necroinflamatory grades in the CHC and AIH samples.
Discussion
Autophagy acts as a bodyguard to protect cells, which involves concerted action to promote cell survival, for example in a state of stress when the cells are starved of nutrients or in case of a viral infection [5, 10]. The importance of autophagy has been emphasized in case of CHC, since it promotes HCV persistence and suppresses the activated immune response [15]. A possible link between autophagy and AIH has also been suggested [9], in which case however its role is not fully understood.
Upon examining LC3 and p62 as indirect indicators of autophagy function, the present study revealed increased levels of these proteins in AIH as compared with CHC. The level of LC3 per se indicates that autophagy is initiated, but may not reflect upon whether autophagy has proceeded or become impaired [21, 31]. A proceeding autophagy implies a low level of p62 since this protein becomes degraded together with the cargo in the autophago-lysosome [32]. Thus, increased LC3 and p62 in AIH suggest that autophagosome formation has occurred without an increase in lysosomal degradation, as has been demonstrated in case of p62 in a recent study [33]. Like further supported by Babuta and co-workers, increased levels of p62 and LC3 were found as the result of chronic alcohol intake in ALD, leading to disruption of autophagy function at lysosome level (impaired autophagic flux) [21].
Decreased level of p62 was observed in CHC contrary to AIH, suggestive of an autophagic function occurring at a much higher rate. This is supported by the observation that the level of p62 decreased within 3–6 days following HCV infection in Huh7.5 cells due to an increase in the autophagy response, which was indicated by an elevated level of LC3 [34].
Increased TOMM20 was detected in AIH as compared with CHC, which might suggest an increase in the number of active mitochondria and also the accumulation of damaged mitochondria not eliminated by autophago-lysosomes [18]. The latter is reported to imply an increased level of p62 in a nonalcoholic steatohepatitis (NASH) study [33]. Thus, the detected high TOMM20 and p62 might indicate that injured mitochondria are most likely not as effectively eliminated in AIH as in CHC.
According to the above mentioned NASH study, enhanced mitophagic level is characterized by elevated LC3/TOMM20 and decreased p62/TOMM20 ratios as the result of palmitic acid administered to mouse primary hepatocytes [33]. As regards the present study, the average LC3/TOMM20 ratios for AIH and CHC were found to be similar, but the average p62/TOMM20 ratio was three times higher in AIH than in CHC. This further signifies the presence of impaired mitophagy in AIH. The fact that Dynamin-related protein 1 (Drp1, a protein associated with mitochondrial dysfunction) was found to be colocalized with TOMM20 indicates that mitochondrial fission is involved in AIH, suggesting a link to hepatic necrosis [35]. It is intriguing that in the present study not only the expression of p62 increased but the necroinflammatory grades were also higher in AIH as compared with CHC, since we observed a strong correlation between p62 expression and necroinflammatory grade. This suggests that decreasing autophagic activity is linked to severity of inflammation; i.e. the more the cellular function becomes compromised due to an extensive inflammation, the less autophagy is capable of rescueing the cells, which may subsequently undergo necrosis. Dysregulated autophagy is reported to contribute to hepatic inflammatory injury and to enhance pro-inflammatory cytokine expression [36]. Stimulation of mTOR, a central issue in autoimmune disorders, also seems to contribute to inflammation in dysfunctional autophagy [37]. The fact that mTOR per se is an inhibitor of autophagy may explain the use of rapamycin as an mTOR inhibitor to treat AIH in the advanced stage [7, 9].
In CHC, the decreased p62/TOMM20 ratio along with the observed decreased level of p62 (one fourth of the expression observed in AIH) and reduced TOMM20 level (1.4 times less than in AIH) are indicative of a more active and proceeding autophagy process. This is supported by a study in which HCV was found to promote mitochondrial fission within 2 days following infection in Huh7 cells, where the damaged organelles were eliminated by mitophagy [38].
Altered lipid metabolism is linked to the life cycle of HCV considering that virions coated with lipoproteins facilitate hepatocyte entry via lipoprotein receptors, however, the HCV-induced ER-stress leads to steatosis [3]. Intracellular lipid droplets being in excess in the cells are degraded by an autophagy process called lipophagy [3,4,5, 22]. Vescovo et al. reported that autophagy acts against the alterations in lipid metabolism induced by HCV and steatosis may develop when autophagy is disrupted [39]. In the present study, no difference was observed in the expression of LC3 and p62 between steatotic or nonsteatotic chronic hepatitis samples (neither in CHC nor in AIH). Nevertheless, the level of p62 seemed to decrease in steatotic CHC as compared with nonsteatotic CHC, but the level did not reach the threshold for statistical significance.
Analysis of autophagy-associated miRNA expression may provide further knowledge about autophagic activity. Several miRNAs are known to downregulate autophagic activity (miR-101, -155 and 204) [21, 22] or their expression is known to be regulated by autophagy (miR-224) [25]. Autophagy-regulating miRNAs have been studied in HCV infection [40] but, to the best of our knowledge, no data is available on how these miRNAs are expressed in AIH.
In the present study, elevated miR-101 was found in CHC samples as compared with AIH cases, which raises concern as miR-101 is an autophagy inhibitor. Suppressed autophagy on the account of miR-101 is reported in breast cancer (targeting RAB5A, STMN1 and ATG4D), HCC (through EZH2) [22, 23] and in ischemic liver (via activating mTOR signaling pathway) [41]. Nevertheless, the levels of LC3 and p62 observed in the present study support the activation and not the inhibition of autophagy in CHC. Intriguingly, increased serum miR-101 expression has been found in HCV as compared with HCC [42]. Moreover, miR-101 has recently been reported to have an antifibrotic effect, most probably via downregulating the PI3K/Akt/mTOR pathway [43]. In the present study, association between miR-101 and fibrosis stage could not be observed in either CHC or AIH, however, when all of the CHC and AIH samples were combined into one group, miR-101 was found to correlate negatively with fibrosis stage (r=-0.36, p < 0.05) and necroinflammatory grade (r=-0.47, p < 0.01). It is to be taken into consideration, however, that the majority of CHC cases (68.9%) displayed low fibrosis stage, whereas the majority of AIH cases (61.1%) showed higher fibrosis stages. Additionally, higher miR-101 expression was found in CHC cases showing low fibrosis stage (F1, F2). Thus, we hypothesize that the relatively higher level of miR-101 might be enough for cells to avoid advanced fibrosis, but not enough to stop autophagy from happening. Further, miRNAs are known to exert their regulatory effect on a number of target genes, influencing several biological processes simultaneously; on the contrary, miR-101 may not be the only miRNA that simultaneously exerts an inhibitory effect on autophagy.
According to our findings, no association could be observed between miR-224 and autophagy. Nevertheless, the level of miR-224 correlated with steatosis in case of the CHC samples, but not in those of AIH. In a previous study, elevated level of miR-224 was found in steatotic CHC in comparison to nonsteatotic CHC [44]. Another study reported miR-224 to be elevated in the event of HCV recurrence in comparison to normal liver and also after administration of interferon/ribavirin treatment as compared with samples taken before treatment [45]. In addition, miR-224 is a predictive factor in the overall survival of hepatoblastoma [46]. miR-224 is a known oncomiR, described to promote proliferation in HCC. In this regard, the correlation of miR-224 with steatosis can be explained by the fact that steatotic CHC is one of the risk factors of HCC [13, 47].
In case of CHC, we found not only increased miR-155 expression in the higher fibrosis stages as compared with the lower stages, but also a correlation between miR-155 and fibrosis stage. In agreement with this, miR-155 knock-out mice were observed to be protected from early and advanced fibrosis in a CCl4 treatment model [48]. Regarding another regulatory aspect, miR-155 was not only found to initiate autophagy in ALD via downregulating mTOR, but also to impair the fusion of autophagosome with lysosome, leading to increased LC3 and p62 levels [21]. Our investigation is partly in agreement with this since high miR-155 expression (which was highest among the miRNAs) was observed in both CHC and AIH samples, but the levels of LC3 and p62 pointed to an impaired autophagy in case of AIH but not in CHC. Nevertheless, a marked increase in miR-155 expression was reported in HCV-infected patients, which seems to be involved in the regulation of antiviral immune response, promoting replication and persistence of HCV [49]. Further data on miR-155 in AIH is scarce, however, this miRNA may have a different role in inflammatory cell recruitment and liver damage [50].
As a summary, the different levels of LC3, p62, TOMM20 observed in the CHC and AIH cases indicate autophagy initiation in the examined samples of chronic hepatitis of different etiology. The increased p62 expression and its correlation with necroinflammatory grade observed in case of AIH suggest an autophagy process which is inhibited at the level of autophago-lysosomal degradation and is associated with extensive inflammation and overactivated immune response, leading to necrosis [51]. In relation to CHC, however, our findings provide further support for the evidence of a functioning autophagy process which seems beneficial for the persistence of HCV in hepatocytes and the pathogenesis of CHC so as to avoid a direct cytopathic effect on the host cells [3]. In our present study, we found no indication that the examined miRNAs had an inhibitory effect on autophagy in regard to either CHC or AIH cases. Instead, the miRNA levels correlated with steatosis (miR-224) and fibrosis (miR-155) in the CHC samples, which could possibly be associated with CHC progression.
References
Jefferies M, Rauff B et al (2018) Update on global epidemiology of viral hepatitis and preventive strategies. World J Clin Cases 6:589–599. https://doi.org/10.12998/wjcc.v6.i13.589
Ríos-Ocampo WA, Navas MC et al (2019) The cellular stress response in hepatitis C virus infection: a balancing act to promote viral persistence and host cell survival. Virus Res 263:1–8. https://doi.org/10.1016/j.virusres.2018.12.013
Vescovo T, Refolo G et al (2014) Autophagy in HCV infection: keeping fat and inflammation at bay. Biomed Res Int 2014:265353. https://doi.org/10.1155/2014/265353
Ke P-Y (2018) The multifaceted roles of autophagy in flavivirus-host interactions. Int J Mol Sci 19:3940. https://doi.org/10.3390/ijms19123940
Ke PY (2019) Diverse functions of autophagy in liver physiology and liver diseases. Int J Mol Sci 20:300. https://doi.org/10.3390/ijms20020300
Jassey A, Liu C-H et al (2019) Hepatitis C virus non-structural protein 5A (NS5A) disrupts mitochondrial dynamics and induces mitophagy. Cells 8:290. https://doi.org/10.3390/cells8040290
Rautou P-E, Mansouri A et al (2010) Autophagy in liver diseases. J Hepatol 53:1123–1134. https://doi.org/10.1016/j.jhep.2010.07.006
Chan ST, Ou JHJ (2017) Hepatitis C virus-induced autophagy and host innate immune response. Viruses 9:E224. https://doi.org/10.3390/v9080224
Golbabapour S, Bagheri-Lankarani K et al (2019) Autoimmune hepatitis and stellate cells; an insight into the role of autophagy. Curr Med Chem - In Press https://doi.org/10.2174/0929867326666190402120231
Czaja MJ, Ding W et al (2013) Functions of autophagy in normal and diseased liver. Autophagy 9:1131–1158. https://doi.org/10.4161/auto.25063
Galluzzi L, Pietrocola F et al (2015) Autophagy in malignant transformation and cancer progression. EMBO J 34:856–880. https://doi.org/10.15252/embj.201490784
Eid N, Ito Y et al (2016) Triggering of Parkin mitochondrial translocation in mitophagy: implications for liver diseases. Front Pharmacol 7:100. https://doi.org/10.3389/fphar.2016.00100
Wang L, Ou J et al (2015) Hepatitis C virus and autophagy. Biol Chem 396:1215–1222. https://doi.org/10.1515/hsz-2015-0172
Ke P-Y, Chen SS-L (2011) Activation of the unfolded protein response and autophagy after hepatitis C virus infection suppresses innate antiviral immunity in vitro. J Clin Invest 121:37–56. https://doi.org/10.1172/JCI41474DS1
Dash S, Chava S et al (2016) Hepatitis C virus infection induces autophagy as a prosurvival mechanism to alleviate hepatic ER-stress response. Viruses 8:E150. https://doi.org/10.3390/v8050150
Christen U, Hintermann E (2016) Immunopathogenic mechanisms of autoimmune hepatitis: how much do we know from animal models? Int J Mol Sci 17:E2007. https://doi.org/10.3390/ijms17122007
Sridhar S, Botbol Y et al (2012) Autophagy and disease: always two sides to a problem. J Pathol 226:255–273. https://doi.org/10.1038/jid.2014.371
Ott M, Norberg E et al (2007) The mitochondrial TOM complex is required for tBid/Bax-induced cytochrome C release. J Biol Chem 282:27633–27639. https://doi.org/10.1074/jbc.M703155200
Sarnyai F, Szekerczés T et al (2019) BGP-15 protects mitochondria in acute, acetaminophen overdose induced liver injury. Pathol Oncol Res - In Press
Martinez-Outschoorn UE, Curry JM et al (2013) Oncogenes and inflammation rewire host energy metabolism in the tumor microenvironment RAS and NFkB target stromal MCT4. Cell Cycle 12:2580–2597. https://doi.org/10.4161/cc.25510
Babuta M, Furi I et al (2019) Dysregulated autophagy and lysosome function are linked to exosome production by micro-RNA 155 in alcoholic liver disease. Hepatology - In Press. https://doi.org/10.1002/hep.30766
Kim MK, Kim SG (2014) Autophagy and microRNA dysregulation in liver diseases. Arch Pharm Res 37:1097–1116. https://doi.org/10.1007/s12272-014-0439-9
Frankel LB, Wen J et al (2011) MicroRNA-101 is a potent inhibitor of autophagy. EMBO J 30:4628–4641. https://doi.org/10.1038/emboj.2011.331
Li X, Liu J (2019) LncRNA NEAT1 promotes autophagy via regulating miR-204/ATG3 and enhanced cell resistance to sorafenib in hepatocellular carcinoma. J Cell Physiol - In Press. https://doi.org/10.1002/jcp.29230
Lan S, Wu SY et al (2014) Autophagy suppresses tumorigenesis of hepatitis B virus-associated hepatocellular carcinoma through degradation of microRNA-224. Hepatology 59:505–517. https://doi.org/10.1002/hep.26659
Ishak K, Baptista A et al (1995) Histological grading and staging of chronic hepatitis. J Hepatoly 22:696–699. https://doi.org/10.1016/0168-8278(95)80226-6
Poynard T, Bedossa P et al (1997) Natural history of liver fibrosis progression in patients with chronic hepatitis C. Lancet 349:825–832. https://doi.org/10.1016/s0140-6736(96)07642-8
Kleiner DE, Makhlouf HR (2016) Histology of NAFLD and NASH in adults and children. Clin Liver Dis 20:293–312. https://doi.org/10.1016/j.cld.2015.10.011.Histology
Doleshal M, Magotra AA et al (2008) Evaluation and validation of total RNA extraction methods for microRNA expression analyses in formalin-fixed, paraffin-embedded tissues. J Mol Diagnostics 10:203–211. https://doi.org/10.2353/jmoldx.2008.070153
Andersen CL, Jensen JL et al (2004) Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res 64:5245–5250. https://doi.org/10.1158/0008-5472.CAN-04-0496
Klionsky DJ, Abdalla FC et al (2012) Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 8:445–544. https://doi.org/10.4161/auto.19496
Islam A, Sooro MA et al (2018) Autophagic regulation of p62 is critical for cancer therapy. Int J Mol Sci 19:E1405. https://doi.org/10.3390/ijms19051405
Zhang N, Li XL et al (2019) Impaired mitophagy triggers NLRP3 inflammasome activation during the progression from nonalcoholic fatty liver to nonalcoholic steatohepatitis. Lab Investig 99:749–763. https://doi.org/10.1038/s41374-018-0177-6
Panigrahi R, Chandra PK et al (2015) Persistent hepatitis C virus infection impairs ribavirin antiviral activity through clathrin-mediated trafficking of equilibrative nucleoside transporter 1. J Virol 89:626–642. https://doi.org/10.1128/jvi.02492-14
He G, Günther C et al (2017) PGAM5-mediated programmed necrosis of hepatocytes drives acute liver injury. Gut 66:716–723. https://doi.org/10.1136/gutjnl-2015-311247
Zhang X, Wu WKK et al (2017) C-X-C motif chemokine 10 impairs autophagy and autolysosome formation in non-alcoholic steatohepatitis. Theranostics 7:2822–2836. https://doi.org/10.7150/thno.19068
Ni H-M, Chao X et al (2019) Dual roles of mammalian target of rapamycin in regulating liver injury and tumorigenesis in autophagy-defective mouse liver. Hepatology - In Press https://doi.org/10.1002/hep.30770
Kim S, Syed GH et al (2014) Hepatitis C virus triggers mitochondrial fission and attenuates apoptosis to promote viral persistence. Proc Natl Acad Sci USA 111:6413–6418. https://doi.org/10.1073/pnas.1321114111
Vescovo T, Romagnoli A et al (2012) Autophagy protects cells from HCV-induced defects in lipid metabolism. Gastroenterology 142:644–653. https://doi.org/10.1053/j.gastro.2011.11.033
Sasaki R, Sur S et al (2019) Repression of microRNA-30e by hepatitis C virus enhances fatty acid synthesis. Hepatol Commun 3:943–953. https://doi.org/10.1002/hep4.1362
Song HU, Du C et al (2019) MicroRNA-101 inhibits autophagy to alleviate liver ischemia / reperfusion injury via regulating the mTOR signaling pathway. Int J Mol Med 43:1331–1342. https://doi.org/10.3892/ijmm.2019.4077
Shakera O, Alhelfa M et al (2017) miRNA-101-1 and miRNA-221 expressions and their polymorphisms as biomarkers for early diagnosis of hepatocellular carcinoma. Infect Genet Evol 51:173–181. https://doi.org/10.1016/j.meegid.2017.03.030
Lei Y, Wang Q, lan et al (2019) MicroRNA-101 suppresses liver fibrosis by downregulating PI3K/Akt/mTOR signaling pathway. Clin Res Hepatol Gastroenterol 43:575–584. https://doi.org/10.1016/j.clinre.2019.02.003
Lendvai G, Jármay K et al (2014) Elevated miR-33a and miR-224 in steatotic chronic hepatitis C liver biopsies. World J Gastroenterol 20:15343–15350. https://doi.org/10.3748/wjg.v20.i41.15343
Gelley F, Zádori G et al (2014) MicroRNA profile before and after antiviral therapy in liver transplant recipients for hepatitis C virus cirrhosis. J Gastroenterol Hepatol 29:121–127. https://doi.org/10.1111/jgh.12362
Gyugos M, Lendvai G et al (2014) microRNA expression might predict prognosis of epithelial hepatoblastoma. Virhows Arch 464:419–427. https://doi.org/10.1007/s00428-014-1549-y
Smedile A, Bugianesi E (2005) Steatosis and hepatocellular carcinoma risk. Eur Rev Med Pharmacol Sci 9:291–295
Bala S, Csak T et al (2016) The pro-inflammatory effects of miR-155 promote liver fibrosis and alcohol-induced steatohepatitis. J Hepatol 64:1378–1387. https://doi.org/10.1016/j.jhep.2016.01.035
Kałużna EM (2014) MicroRNA-155 and microRNA-196b: promising biomarkers in hepatitis C virus infection? Rev Med Virol 24:169–185. https://doi.org/10.1002/rmv.1785
Blaya D, Aguilar-Bravo B et al (2018) Expression of microRNA-155 in inflammatory cells modulates liver injury. Hepatology 68:691–706. https://doi.org/10.1002/hep.29833
Tiniakos DG, Brain JG et al (2015) Role of histopathology in autoimmune hepatitis. Dig Dis 33:53–64. https://doi.org/10.1159/000440747
Acknowledgements
The authors thank Mrs. Erzsébet Kovács for the technical assistance and Mrs. Elvira Kálé Rigóné for the English proofreading.
Funding
Open access funding provided by Semmelweis University (SE). This work was supported by grants OTKA 108548; OTKA 128881 from the Hungarian National Research Foundation, NVKP_16_1-2016-0004 from the Hungarian National Research, Development and Innovation Office and EFOP-3.6.3-VEKOP-16-2017-00009 from Semmelweis University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest with respect to the research, authorship, and/or publication of this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Szekerczés, T., Gógl, A., Illyés, I. et al. Autophagy, Mitophagy and MicroRNA Expression in Chronic Hepatitis C and Autoimmune Hepatitis. Pathol. Oncol. Res. 26, 2143–2151 (2020). https://doi.org/10.1007/s12253-020-00799-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12253-020-00799-y