A Novel Digestive Proteinase Lipase Member H-A in Bombyx mori Contributes to Digestive Juice Antiviral Activity against B. mori Nucleopolyhedrovirus


Abstract
:1. Introduction
2. Materials and Methods
2.1. B. mori Rearing and Virus Preparation
2.2. BmN Cell Culture and Transfection
2.3. Bioinformatics Analysis
2.4. RNA Isolation and cDNA Synthesis
2.5. Quantitative Reverse Transcription PCR (RT-qPCR)
2.6. Prokaryotic Expression, Antibody Preparation
2.7. Western Blot Analyses
2.8. Analysis of Viral Propagation in B. mori Midgut Infected by BmNPV with BmLHA Treated
2.9. Analysis of Viral Propagation in BmN Cells Infected by BmNPV with BmLHA Treated
2.10. Overexpression of BmLHA in BmN Cells and BmNPV Infection
2.11. Cell Viability Counting Assay
3. Results
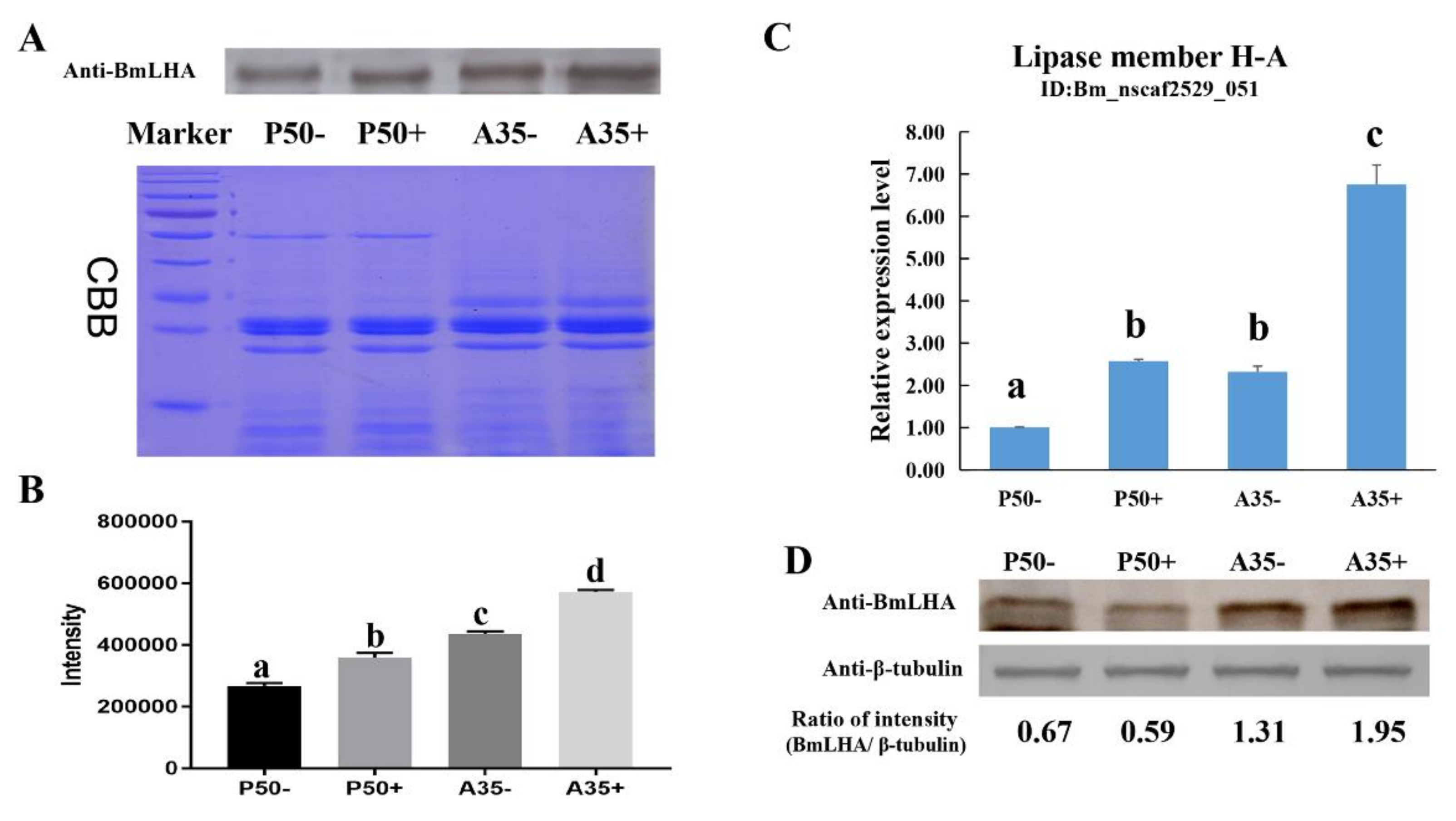
3.1. Analysis and Validation of BmLHA Omics Data
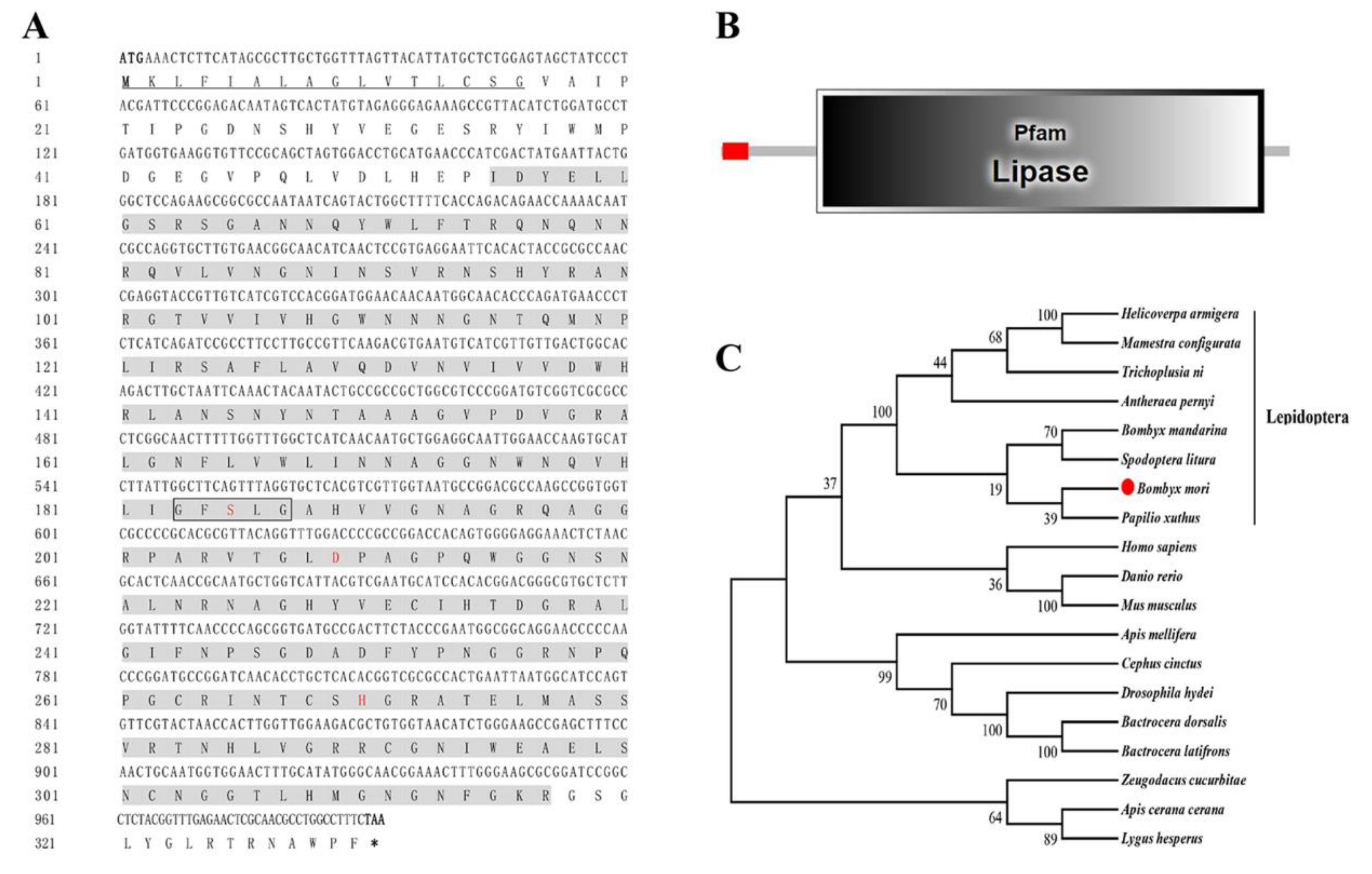
3.2. Characterization of the BmLHA Sequence
3.3. The Analyzed of Expression Pattern of BmLHA
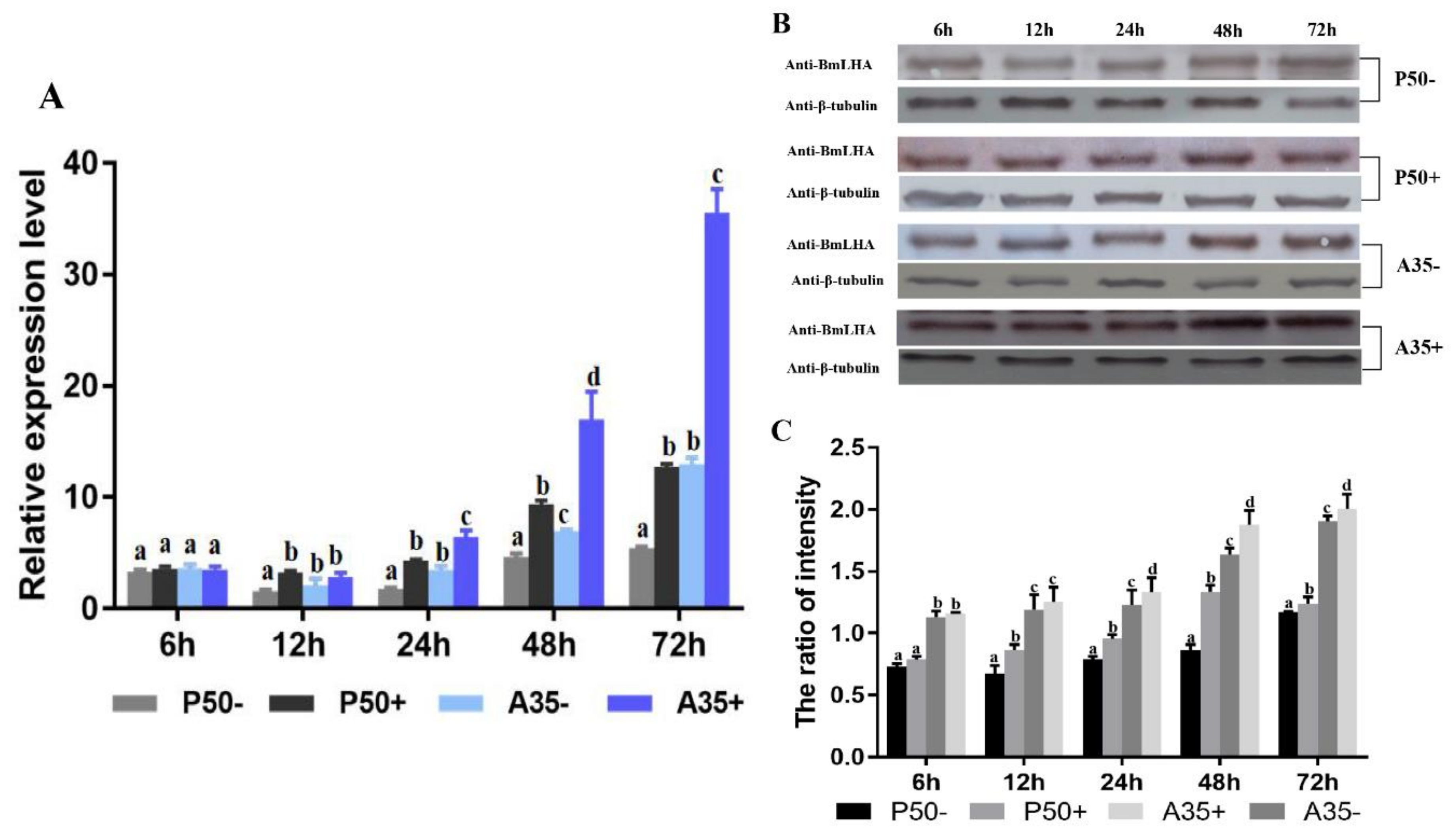
3.4. Variation of BmLHA Expression Level Following BmNPV Infection in Different Resistant Strains
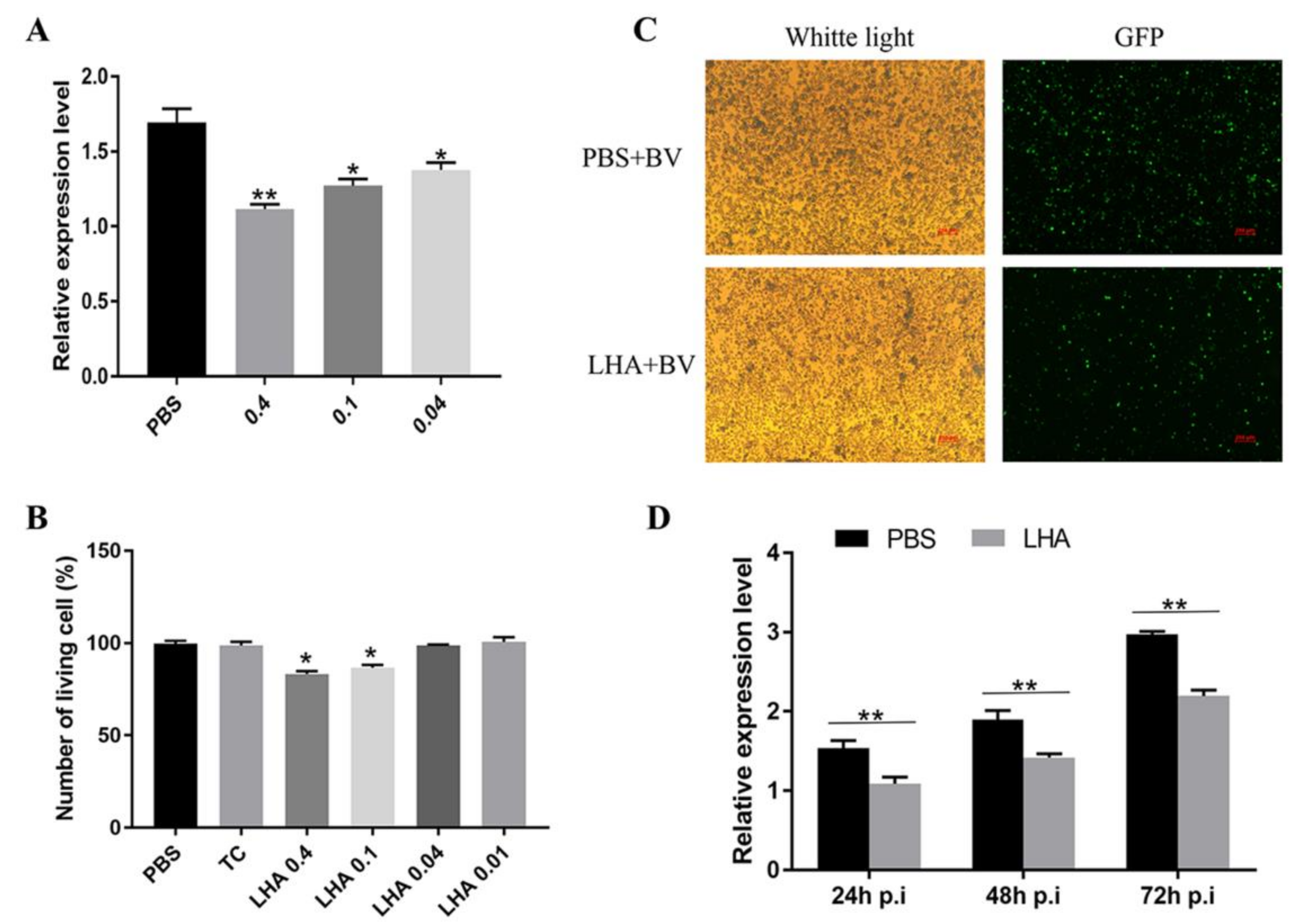
3.5. Antiviral Effects Analysis of BmLHA in B. mori Larvae
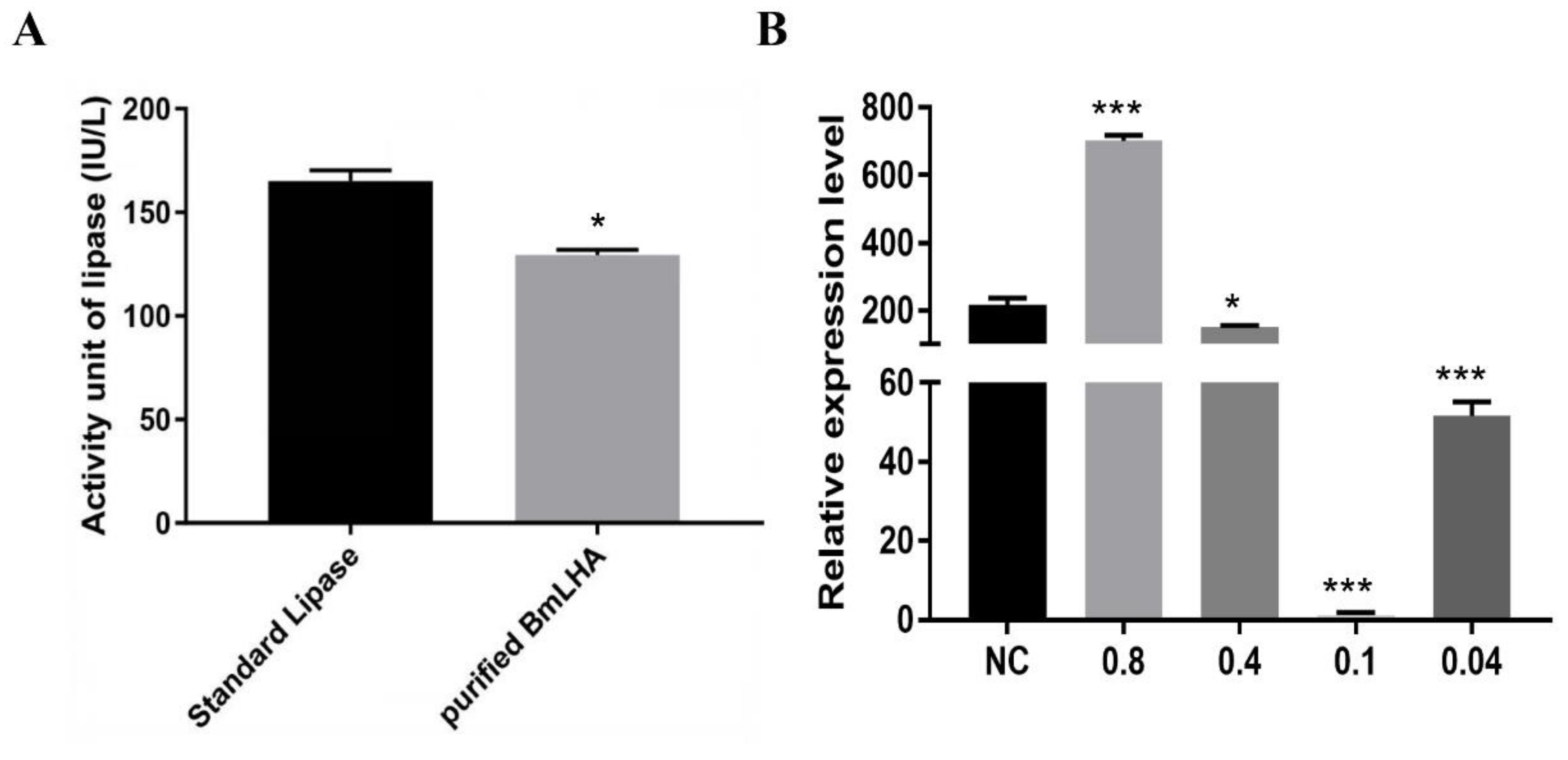
3.6. Antiviral Effects Analysis of BmLHA in BmN Cells
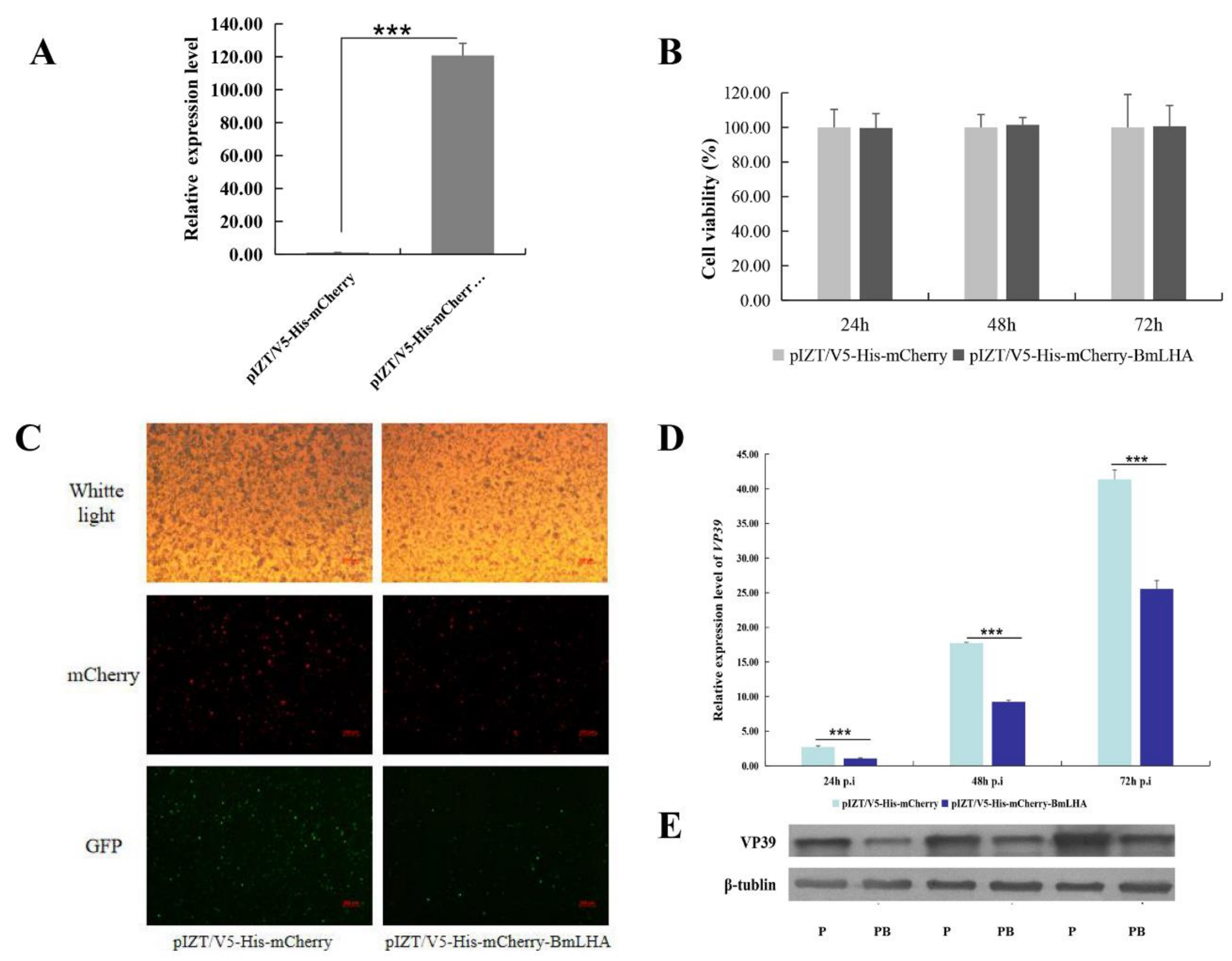
3.7. Antiviral Effects Analysis in BmLHA Overexpressed BmN Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef]
- Shao, Q.M.; Yang, B.; Xu, Q.Y.; Li, X.Q.; Lu, Z.Q.; Wang, C.S.; Huang, Y.P.; Soderhall, K.; Ling, E.J. Hindgut Innate Immunity and Regulation of Fecal Microbiota through Melanization in Insects. J. Biol. Chem. 2012, 287, 14270–14279. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Xia, Q.Y. The progress and future of enhancing antiviral capacity by transgenic technology in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2014, 48, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.Q.; Hou, C.X.; Bi, H.L.; Wang, Y.Q.; Xu, J.; Li, M.W.; James, A.A.; Huang, Y.P.; Tan, A.J. Transgenic Clustered Regularly Interspaced Short Palindromic Repeat/Cas9-Mediated Viral Gene Targeting for Antiviral Therapy of Bombyx mori Nucleopolyhedrovirus. J. Virol. 2017, 91, 13. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.Y.; Tang, X.D.; Lv, Z.Y.; Wang, X.Y.; Tian, C.H.; Xu, Y.P.; Zhang, C.X. Gene expression profiling of resistant and susceptible Bombyx mori strains reveals nucleopolyhedrovirus-associated variations in host gene transcript levels. Genomics 2009, 94, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Yu, H.Z.; Geng, L.; Xu, J.P.; Yu, D.; Zhang, S.Z.; Ma, Y.; Fei, D.Q. Comparative Transcriptome Analysis of Bombyx mori (Lepidoptera) Larval Midgut Response to BmNPV in Susceptible and Near-Isogenic Resistant Strains. PLoS ONE 2016, 11, 22. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yu, H.Z.; Xu, J.P.; Zhang, S.Z.; Yu, D.; Liu, M.H.; Wang, L.L. Comparative Subcellular Proteomics Analysis of Susceptible and Near-isogenic Resistant Bombyx mori (Lepidoptera) Larval Midgut Response to BmNPV infection. Sci. Rep. 2017, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xia, D.G.; Zhao, Q.L.; Zhang, G.Z.; Zhang, Y.S.; Qiu, Z.Y.; Shen, D.X.; Lu, C. Label-free proteomic analysis of silkworm midgut infected by Bombyx mori nuclear polyhedrosis virus. J. Proteom. 2019, 200, 40–50. [Google Scholar] [CrossRef]
- Yu, H.Z.; Wang, X.Y.; Xu, J.P.; Ma, Y.; Zhang, S.Z.; Yu, D.; Fei, D.Q.; Muhammad, A. iTRAQ-based quantitative proteomics analysis of molecular mechanisms associated with Bombyx mori (Lepidoptera) larval midgut response to BmNPV in susceptible and near-isogenic strains. J. Proteom. 2017, 165, 35–50. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, W.Q.; Guo, H.Z.; Dang, Y.H.; Cheng, T.C.; Yang, W.Y.; Sun, Q.; Wang, B.B.; Wang, Y.M.; Xie, E.Y.; et al. Distinct Functions of Bombyx mori Peptidoglycan Recognition Protein 2 in Immune Responses to Bacteria and Viruses. Front. Immunol. 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.H.; Dong, X.L.; Pan, C.X.; Du, G.Y.; Wu, Y.F.; Yang, J.G.; Chen, P.; Lu, C.; Pan, M.H. A newly discovered member of the Atlastin family, BmAtlastin-n, has an antiviral effect against BmNPV in Bombyx mori. Sci. Rep. 2016, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.T.; Tan, L.R.; Hu, N.; Dong, Z.Q.; Hu, Z.G.; Jiang, Y.M.; Chen, P.; Pan, M.H.; Lu, C. C-lysozyme contributes to antiviral immunity in Bombyx mori against nucleopolyhedrovirus infection. J. Insect Physiol. 2018, 108, 54–60. [Google Scholar] [CrossRef]
- Hu, Z.-G.; Dong, Z.-Q.; Dong, F.-F.; Zhu, Y.; Chen, P.; Lu, C.; Pan, M.-H. Identification of a PP2A gene in Bombyx mori with antiviral function against B. mori nucleopolyhedrovirus. Insect Sci. 2019. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, L.; Zhou, X.L.; Zhu, Y.; Dong, Z.Q.; Chen, P.; Lu, C.; Pan, M.H. BmAtg13 promotes the replication and proliferation of Bombyx mori nucleopolyhedrovirus. Pest. Biochem. Physiol. 2019, 157, 143–151. [Google Scholar] [CrossRef]
- Feng, M.; Kong, X.S.; Zhang, J.J.; Xu, W.F.; Wu, X.F. Identification of a novel host protein SINAL10 interacting with GP64 and its role in Bombyx mori nucleopolyhedrovirus infection. Virus Res. 2018, 247, 102–110. [Google Scholar] [CrossRef]
- Wang, X.Y.; Shao, Z.M.; Chen, Q.Y.; Xu, J.P.; Sun, X.; Xu, Z.P.; Li, M.W.; Wu, Y.C. Knockdown of BmTCP-1 beta Delays BmNPV Infection in vitro. Front. Microbiol. 2019, 10, 10. [Google Scholar] [CrossRef]
- Pauchet, Y.; Muck, A.; Svatos, A.; Heckel, D.G.; Preiss, S. Mapping the larval midgut lumen proteorne of Helicoverpa armigera, a generalist herbivorous insect. J. Proteome Res. 2008, 7, 1629–1639. [Google Scholar] [CrossRef]
- Wu, K.; Yang, B.; Huang, W.R.; Dobens, L.; Song, H.S.; Ling, E.J. Gut immunity in Lepidopteran insects. Dev. Comp. Immunol. 2016, 64, 65–74. [Google Scholar] [CrossRef]
- Sunagar, S.G.; Savanurmath, C.J.; Hinchigeri, S.B. The profiles of red fluorescent proteins with antinucleopolyhedrovirus activity in races of the silkworm Bombyx mori. J. Insect Physiol. 2011, 57, 1707–1714. [Google Scholar] [CrossRef]
- Ponnuvel, K.M.; Nakazawa, H.; Furukawa, S.; Asaoka, A.; Ishibashi, J.; Tanaka, H.; Yamakawa, M. A lipase isolated from the silkworm Bombyx mori shows antiviral activity against nucleopolyhedrovirus. J. Virol. 2003, 77, 10725–10729. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, H.; Tsuneishi, E.; Ponnuvel, K.M.; Furukawa, S.; Asaoka, A.; Tanaka, H.; Ishibashi, J.; Yamakawa, M. Antiviral activity of a serine protease from the digestive juice of Bombyx mori larvae against nucleopolyhedrovirus. Virology 2004, 321, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Ponnuvel, K.M.; Nithya, K.; Sirigineedi, S.; Awasthi, A.K.; Yamakawa, M. In vitro antiviral activity of an Alkaline trypsin from the digestive juice of Bombyx mori larvae against nucleopolyherovirus. Arch. Insect Biochem. Physiol. 2012, 81, 90–104. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Reetz, M.T. Microbial lipases form versatile tools for biotechnology. Trends Biotechnol. 1998, 16, 396–403. [Google Scholar] [CrossRef]
- Xu, L.J.; Hou, Q.L.; Zhao, Y.J.; Lu, L.H.; Li, B.Y.; Ni, Z.F.; Liang, R.Q. Silencing of a lipase maturation factor 2-like gene by wheat-mediated RNAi reduces the survivability and reproductive capacity of the grain aphid, Sitobion avenae. Arch. Insect Biochem. Physiol. 2017, 95, 9. [Google Scholar] [CrossRef]
- Xu, L.; Huang, H.J.; Zhou, X.; Liu, C.W.; Bao, Y.Y. Pancreatic lipase-related protein 2 is essential for egg hatching in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2017, 26, 277–285. [Google Scholar] [CrossRef]
- Zhang, S.D.; Li, X.; Bin, Z.; Du, M.F.; Yin, X.M.; An, S.H. Molecular identification of a pancreatic lipase-like gene involved in sex pheromone biosynthesis of Bombyx mori. Insect Sci. 2014, 21, 459–468. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Zhao, X.; Qian, C.; Wei, G.; Zhu, B.; Liu, C. Expression and characterization of a lipase-related protein in the malpighian tubules of the Chinese oak silkworm, Antheraea pernyi. Bull. Entomol. Res. 2016, 106, 615–623. [Google Scholar] [CrossRef]
- Liu, R.H.; Wang, W.H.; Liu, X.Y.; Lu, Y.; Xiang, T.T.; Zhou, W.; Wan, Y.J. Characterization of a Lipase From the Silkworm Intestinal Bacterium Bacillus pumilus With Antiviral Activity Against Bombyx mori (Lepidoptera: Bombycidae) Nucleopolyhedrovirus in vitro. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, G.H.; Cheng, T.C.; Yang, Q.; Jin, S.K.; Lu, G.; Wu, F.Q.; Xiao, Y.; Xu, H.F.; Xia, Q.Y. Resistance to Bombyx mori nucleopolyhedrovirus via overexpression of an endogenous antiviral gene in transgenic silkworms. Arch. Virol. 2012, 157, 1323–1328. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, X.Y.; Du, C.; Gao, J.; Xu, J.P. Expression analysis of several antiviral related genes to BmNPV in different resistant strains of silkworm, Bombyx mori. J. Insect Sci. 2014, 14, 9. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Wang, J.; Zhu, L.B.; Toufeeq, S.; Xu, X.; You, L.L.; Li, B.; Hu, P.; Xu, J.P. Quantitative label-free proteomic analysis reveals differentially expressed proteins in the digestive juice of resistant versus susceptible silkworm strains and their predicted impacts on BmNPV infection. J. Proteom. 2020, 210, 13. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Brevnov, M.G.; Pawar, H.S.; Mundt, J.; Calandro, L.M.; Furtado, M.R.; Shewale, J.G. Developmental Validation of the PrepFiler (TM) Forensic DNA Extraction Kit for Extraction of Genomic DNA from Biological Samples. J. Forensic Sci. 2009, 54, 599–607. [Google Scholar] [CrossRef]
- Selot, R.; Kumar, V.; Shukla, S.; Chandrakuntal, K.; Brahmaraju, M.; Dandin, S.B.; Laloraya, M.; Kumar, P.G. Identification of a soluble NADPH oxidoreductase (BmNOX) with antiviral activites in the gut juice of Bombyx mori. Biosci. Biotechnol. Biochem. 2007, 71, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.J.; Zhan, M.Y.; Ye, C.; Yu, X.Q.; Rao, X.J. Molecular cloning and characterization of a short peptidoglycan recognition protein from silkworm Bombyx mori. Insect Mol. Biol. 2017, 26, 665–676. [Google Scholar] [CrossRef]
- Rivera-Perez, C.; Garcia-Carreno, F.L.; Saborowski, R. Purification and Biochemical Characterization of Digestive Lipase in Whiteleg Shrimp. Mar. Biotechnol. 2011, 13, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Smede, M.; Hussain, M.; Asgari, S. A lipase-like gene from Heliothis virescens ascovirus (HvAV-3e) is essential for virus replication and cell cleavage. Virus Genes 2009, 39, 409–417. [Google Scholar] [CrossRef]
- Gan, L.P.; Liu, R.H.; Li, Y.J. Serine proteinase (SP) and serine protease inhibitor (serpin) gene expression after hormone treatment in the silkworm Bombyx mori (Lepidoptera: Bombycidae). J. Asia-Pac. Entomol. 2016, 19, 1203–1210. [Google Scholar]
- Li, G.H.; Zhou, Q.; Qiu, L.P.; Yao, Q.; Chen, K.P.; Tang, Q.; Hu, Z.Y. Serine protease Bm-SP142 was differentially expressed in resistant and susceptible Bombyx mori strains, involving in the defence response to viral infection. PLoS ONE 2017, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Li, Y.S.; Tang, X.; Guo, P.C.; Wang, D.D.; Zhou, C.Y.; Xia, Q.Y.; Zhao, P. A midgut-specific serine protease, BmSP36, is involved in dietary protein digestion in the silkworm, Bombyx mori. Insect Sci. 2017, 24, 753–767. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, G.H.; Dong, Z.M.; Duan, J.; Xu, P.Z.; Cheng, T.C.; Xiang, Z.H.; Xia, Q.Y. Genome-wide identification and expression analysis of serine proteases and homologs in the silkworm Bombyx mori. BMC Genom. 2010, 11. [Google Scholar] [CrossRef] [Green Version]
- Fei, D.Q.; Yu, H.Z.; Xu, J.P.; Zhang, S.Z.; Wang, J.; Li, B.; Yang, L.A.; Hu, P.; Xu, X.; Zhao, K.; et al. Isolation of ferritin and its interaction with BmNPV in the silkworm, Bombyx mori. Dev. Comp. Immunol. 2018, 86, 130–137. [Google Scholar] [CrossRef]
- Wang, X.Y.; Wu, K.H.; Pang, H.L.; Xu, P.Z.; Li, M.W.; Zhang, G.Z. Study on the Role of Cytc in Response to BmNPV Infection in Silkworm, Bombyx mori (Lepidoptera). Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.X.; Zhang, C.; Ru, W.J.; Wang, D.; Zhang, W.P. Effects of overexpression and inhibited expression of thymosin, an actin-interacting protein from Bombyx mori, on BmNPV proliferation and replication. Arch. Insect Biochem. Physiol. 2018, 98, 10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| BmLHA | GTGATGCCGACTTCTACCCG | ACCACAGCGTCTTCCAACCA |
| VP39 | CAACTTTTTGCGAAACGACTT | GGCTACACCTCCACTTGCTT |
| BmGAPDH | CGATTCAACATTCCAGAGCA | GAACACCATAGCAAGCACGAC |
| Protein ID | Protein Description | Unique Peptides | P50+_vs._ P50− Ratio | A35+_vs._ A35− Ratio | A35−_vs._ P50− Ratio | A35+_vs._ P50+ Ratio |
|---|---|---|---|---|---|---|
| Bm_nscaf 2529_051 | Lipase member H-A |
| 1.296 | 3.025 | 4.164 | 9.716 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-Z.; Zhu, L.-B.; You, L.-L.; Wang, J.; Cao, H.-H.; Liu, Y.-X.; Toufeeq, S.; Wang, Y.-L.; Kong, X.; Xu, J.-P. A Novel Digestive Proteinase Lipase Member H-A in Bombyx mori Contributes to Digestive Juice Antiviral Activity against B. mori Nucleopolyhedrovirus. Insects 2020, 11, 154. https://doi.org/10.3390/insects11030154
Zhang S-Z, Zhu L-B, You L-L, Wang J, Cao H-H, Liu Y-X, Toufeeq S, Wang Y-L, Kong X, Xu J-P. A Novel Digestive Proteinase Lipase Member H-A in Bombyx mori Contributes to Digestive Juice Antiviral Activity against B. mori Nucleopolyhedrovirus. Insects. 2020; 11(3):154. https://doi.org/10.3390/insects11030154
Chicago/Turabian StyleZhang, Shang-Zhi, Lin-Bao Zhu, Ling-Ling You, Jie Wang, Hui-Hua Cao, Ying-Xue Liu, Shahzad Toufeeq, Yu-Ling Wang, Xue Kong, and Jia-Ping Xu. 2020. "A Novel Digestive Proteinase Lipase Member H-A in Bombyx mori Contributes to Digestive Juice Antiviral Activity against B. mori Nucleopolyhedrovirus" Insects 11, no. 3: 154. https://doi.org/10.3390/insects11030154