
Abstract
Our ability to predict invasions has been hindered by the seemingly idiosyncratic context-dependency of individual invasions. However, we argue that robust and useful generalisations in invasion science can be made by considering “invasion syndromes” which we define as “a combination of pathways, alien species traits, and characteristics of the recipient ecosystem which collectively result in predictable dynamics and impacts, and that can be managed effectively using specific policy and management actions”. We describe this approach and outline examples that highlight its utility, including: cacti with clonal fragmentation in arid ecosystems; small aquatic organisms introduced through ballast water in harbours; large ranid frogs with frequent secondary transfers; piscivorous freshwater fishes in connected aquatic ecosystems; plant invasions in high-elevation areas; tall-statured grasses; and tree-feeding insects in forests with suitable hosts. We propose a systematic method for identifying and delimiting invasion syndromes. We argue that invasion syndromes can account for the context-dependency of biological invasions while incorporating insights from comparative studies. Adopting this approach will help to structure thinking, identify transferrable risk assessment and management lessons, and highlight similarities among events that were previously considered disparate invasion phenomena.
Similar content being viewed by others
Introduction
A major challenge in invasion science is to identify general patterns that help us to predict, prevent and manage biological invasions. To this end, recent research has focused on identifying pathways by which alien taxa are likely to be introduced and disseminated (Reichard and White 2001; Hulme 2009; Essl et al. 2015; Pergl et al. 2017; Saul et al. 2017), alien taxa most likely to become invasive and cause impact (Hayes and Barry 2008; Tingley et al. 2010; van Kleunen et al. 2010; Pyšek et al. 2012b; Hawkins et al. 2015; Kumschick et al. 2015; Bacher et al. 2018), and environments that are particularly susceptible to invasion and impacts from alien taxa (Chytrý et al. 2008a, b; Catford et al. 2012; Guo et al. 2015). Some of the more robust and broadly applicable invasion patterns (Pyšek and Richardson 2006) include: the probability that invasion increases with propagule pressure (Cassey et al. 2018); alien taxa with large native ranges are more likely to become invasive and cause impact than those with smaller ranges (Rejmánek and Richardson 1996; Goodwin et al. 1999; Pyšek et al. 2009; Shah et al. 2011); and isolated oceanic islands are more susceptible to the establishment of alien taxa than continental regions (van Kleunen et al. 2015; Dawson et al. 2017; Pyšek et al. 2017; Delavaux et al. 2019). There are, however, many exceptions to such generalizations (Kueffer et al. 2013). The probability of an invasion can be insensitive to propagule pressures across a wide range of values (e.g. if invasions are simply not possible due to incompatible environmental conditions, or if an invasion is likely to result from the introduction of a single propagule; Bacon et al. 2014; Duncan et al. 2014). Similarly, while the positive relationship between native range size and the likelihood of an alien species becoming established and/or causing an undesirable impact has been demonstrated for some aquatic (Bates et al. 2013), bird (Duncan et al. 2001), mammal (Forsyth et al. 2004) and plant (Pyšek et al. 2009; Hui et al. 2011; Moodley et al. 2013; Potgieter et al. 2014; Moodley et al. 2016; Novoa et al. 2016b) species, such correlations are not always clear-cut (Jeschke and Strayer 2006; Swart et al. 2018). New formulations and fine-tuning of generalizations are thus needed (Jeschke et al. 2012; Kueffer et al. 2013).
Two main approaches have been proposed to deal with context-dependencies in ecology. Some authors suggest focusing research efforts on finding relatively simple general patterns at large scales (i.e. large numbers of species, large spatial scales, or over long time periods) (Lawton 1999; Hui et al. 2013; Prins and Gordon 2014). However, such general patterns informed by ‘big’ data have many exceptions and are often heuristic—useful but with limited predictive value. For example, they are not particularly helpful when deciding whether a specific alien species of potential commercial value can be imported or whether it poses an unacceptable risk (Keller and Kumschick 2017). Others have argued against seeking generalizations and instead propose focusing on small scales to collate and catalogue comprehensive case studies containing more rigorous evidence (Crawley 1987; Sun et al. 2013; Simberloff 2014). The second approach helps us understand and manage particular invasions. However, studying each ongoing invasion separately is incredibly costly (Dawson et al. 2017). It is also unclear how insights gained from the increasing numbers of case studies can be distilled to provide general lessons for management.
In response to this challenge, Kueffer et al. (2013) proposed that invasion scientists should focus on identifying “typical recurrent associations of species biology and invasion dynamics with particular invasion contexts such as an invasion stage, invaded habitat and/or socioeconomic context” (p. 616), which they termed “invasion syndromes” (see Box 1 for a list of definitions). The premise of “invasion syndromes” is that no single combination of factors is applicable to all invasions and determines which management options are appropriate for all alien taxa in the same way, but that it is still possible to find useful general patterns (Perkins and Nowak 2013) that characterize groups of invasion events. The key challenge is to determine the shared context under which generalizations are possible, robust, and useful (Kueffer 2012).
The concept of “invasion syndromes” is often confused with the traditional concept of “model systems”, defined by Kueffer et al. (2013) as “in-depth research of particular invasions of particular species or in a particular site” (p. 616). Model systems are, however, groups of taxa/ecosystems that contain many species/sites, have a long history of introduction/invasion, contain many species at different stages of the continuum and/or a large literature exist on their invasion, allowing for in-depth studies (e.g. Moodley et al. 2013; Richardson et al. 2011). The results from model system research may therefore allow us to identify recurrent patterns of species-ecosystem (pathways) interactions, i.e. ‘invasion syndromes’.
A revised definition
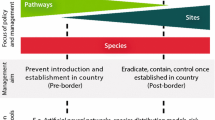
Here we revise the definition of an invasion syndrome as “a combination of pathways, alien species traits, and characteristics of the recipient ecosystem which collectively result in predictable dynamics and impacts, and that can be managed effectively using specific policy and management actions” (Fig. 1). Our definition builds on Kueffer et al. (2013), as well as some more recent studies (McGeoch et al. 2016; Wilson et al. 2018; Latombe et al. 2019a), in several key respects. To improve our understanding of biological invasions and how to best manage them, the context of any invasion event must explicitly consider human actions or pathways, the traits of the introduced taxa (which determine their invasiveness), and the characteristics of the recipient ecosystems (which determine their invasibility), as well as any interactions between these factors (Wilson et al. 2018). We also specify the outcomes (i.e. invasion dynamics and impacts) of invasion events in the definition, with the intention that the invasion syndrome approach is explicitly designed to improve management efficacy.

An invasion syndrome is defined as a combination of pathways, alien species traits, and characteristics of the recipient ecosystem which collectively result in predictable dynamics and impacts, and that can be managed effectively using specific policy and management actions. For it to be coherent, the shared characteristics (pathways, alien species traits, and characteristics of the recipient ecosystem) must result in predictable outcomes (regarding invasion dynamics and impacts) which in turn can be best managed using similar management or policy responses. This means an invasion syndrome should be created from generalizations that are as broad as possible, but which are still robust and useful. The invasion context is displayed here on three vertical axes (i.e. vertical black bars) that range from general (at the top) to specific (at the bottom). For example, the alien species traits axis could vary (top/general to bottom/specific) from all aquatic species, to aquatic species within a specific genus, to congeneric freshwater species within a specific body size range. The positions along the axes (i.e. black boxes) are adjusted so that all invasion events within the selected context result in similar outcomes and response options. A change in any one of the axes, or a change in the outcomes or response options that are to be encompassed by the invasion syndrome, will likely affect all other aspects of the framework, which means that circumscribing a syndrome is an iterative process
Pathways
Pathways are defined as “a combination of processes and opportunities resulting in the movement of propagules from one area to another” (Richardson et al. 2011, p. 412). The different pathways by which alien taxa are intentionally or unintentionally introduced to areas outside their native range and/or spread within their introduced range (Hulme et al. 2008) can influence the dynamics of their invasion or impacts (Lambdon and Hulme 2006; Westphal et al. 2008; Wilson et al. 2009; Kueffer 2017; Pergl et al. 2017). For example, since the number of individuals moved and the frequency of these movements depend on the characteristics of the pathway used, pathways are intrinsically linked to colonization and propagule pressure, which in turn influence invasion outcomes (Lockwood et al. 2009, 2013). Some species might also lack the opportunity to invade because no pathway currently exists to move them beyond their native range. Therefore, assessing particular pathways of introduction and spread is crucial for enhancing prevention and, more generally, for guiding policies and management responses to invasions (Padilla and Williams 2004; Hulme 2009; Kikillus et al. 2012; Essl et al. 2015).
Pathways can be characterized in general terms (e.g. “disseminated as ornamental plants”), or more specifically by identifying vectors involved in the introduction of particular alien taxa from specific donor regions (Hulme 2009; Novoa et al. 2017). Whatever the level of pathway specificity, the goal should be to provide insights of direct relevance for management and policy. For example, identifying the stakeholder groups associated with particular pathways can provide valuable data about the characteristics of each pathway (e.g. the areas where the species are moved to, and the identity and number of species moved), and promote responsible behaviour (Cole et al. 2019).
The Convention on Biological Diversity (CBD) recently adopted a hierarchical system of classifying introduction pathways in particular (Hulme et al. 2008; CBD 2014). This is being applied in the implementation of the European Union Regulation 1143/2014 on invasive alien species and is being used in South Africa for its national-level reporting on biological invasions (van Wilgen and Wilson 2018). While these are important and increasingly used classification schemes, it is often also important to explicitly consider key characteristics of the introduction pathways, for example, the frequency of introduction, the vectors involved, and the likelihood of co-introduction of different alien species (Wilson et al. 2009).
Alien species traits
Species traits are attributes that relate to how species interact with the abiotic environment and with other species (Díaz and Cabido 2001). Possessing certain traits conveys advantages for alien species [e.g. alien plants with longer flowering durations tend to be more likely pollinated (Cadotte et al. 2006), and therefore more invasive]. Research on how traits differ among species has been a topic of particular interest in invasion science because it is thought that identifying and comparing species traits associated with invasion dynamics and impact can improve the prediction and management of invasions. Such research has sought to link species traits with invasion outcomes (Pyšek and Richardson 2007; Pyšek et al. 2009; Capellini et al. 2015; Mahoney et al. 2015; Gallien and Carboni 2017), impacts (Nentwig et al. 2010; Pyšek et al. 2012b; Elleouet et al. 2014; Novoa et al. 2016b; Measey et al. 2016), and policy and management actions (Murray et al. 2011; Novoa et al. 2015a). Within the invasion syndrome approach, a wide variety of alien species traits (from life-history traits to behavioural traits or ecological preferences; see Supplementary information for examples) can be selected to adjust the context.
Characteristics of the recipient ecosystem
The biotic and abiotic characteristics of the recipient ecosystem, including its anthropogenic modifications (Kueffer 2017), influence alien species’ invasion dynamics and impact (Hood and Naiman 2000; Denslow 2003; Riley et al. 2005; Johnson et al. 2008; Catford et al. 2009; Vermonden et al. 2010; Pyšek et al. 2012b, 2017). A wide range of characteristics of the recipient ecosystem can be important: from broad classifications such as biome types (Campbell 1996), ecoregions (Olson and Dinerstein 2002; Abell et al. 2008) or habitats (Chytrý et al. 2008a, b; Latombe et al. 2019b), to more specific abiotic (e.g. altitude, precipitation or nutrient availability; Chytrý et al. 2008a, b), biotic (e.g. native biodiversity or abundance of mutualists or natural enemies; Le Roux et al. 2017; Latombe et al. 2018; Hui and Richardson 2019) or socioeconomic characteristics and processes (e.g. national wealth or human population density; Pyšek et al. 2010). Biotic and abiotic matches between the donor and the recipient ecosystems can also influence the outcomes of invasions (Thuiller et al. 2005; Gallien et al. 2015; Hui et al. 2016), their impact (Ricciardi and Atkinson 2004), and management (Sun et al. 2017).
Outcomes
To be considered as an invasion syndrome, the invasion events defined by the context (i.e. the pathways, alien species traits, and characteristics of the recipient ecosystem) must result in similar outcomes, i.e. they must share similar invasion dynamics or cause similar impacts. In other words, the outcomes of an invasion event defined by the context need to be predictable. Invasion dynamics refer both to the dynamics from introduction to invasion [e.g. a long lag phase, rapid long-distance dispersal, and more generally the path to commonness (McGeoch and Latombe 2016)]; to general properties like extent, local abundance, dispersal patterns; and potentially, biotic interactions (Hui and Richardson 2017). Impacts refer to a wide range of both positive and negative environmental and socioeconomic changes that invasive alien species can cause in the social-ecological systems to which they are introduced (Shackleton et al. 2007; Binimelis et al. 2008; Kull et al. 2011; Vilà et al. 2011; Pyšek et al. 2012b; Gallardo et al. 2016; Gallien et al. 2017; Zengeya et al. 2017). For example, depending on the context, invasive alien species can cause changes in the biodiversity or the ecosystem properties of the invaded areas (Pyšek et al. 2012a; Blackburn et al. 2014), or affect human well-being (Vilà and Hulme 2017; Bacher et al. 2018, Shackleton et al. 2019). These impacts result, in part, from the invasion dynamics themselves (e.g. extent, abundance, dispersal, and biotic interactions).
Response options
The context and outcomes associated with a particular syndrome will affect the set of suitable response options for managing invasions. For an invasion syndrome to be of practical value, there have to be some general rules as to which management responses are effective to deal with invasion events included in the syndrome. This might include steps taken to prevent invasive alien species from entering a new area; if introduced, efforts to remove species before they become widely established; and if species are widely established, and it is no longer possible to remove them, actions to limit negative impacts while retaining any benefits (van Wilgen et al. 2011; Shackleton et al. 2017; Novoa et al. 2019).
Examples of invasion syndromes
Invasion syndromes occur across a broad range of taxonomic groups and environments (see Fig. 2 for a schematic summary), and in the following section we discuss seven examples to highlight this diversity.


Examples of seven invasion syndromes proposed in this paper. See text for explanation.
A. Cacti with clonal fragmentation in arid ecosystems
The Cactaceae family contains 1919 species of succulent plants (Novoa et al. 2015b), almost all of which are endemic to the Americas (Novoa et al. 2016a). Fifty-seven cactus species are currently listed as invasive around the world (Novoa et al. 2015b). While all cacti have thick, fleshy and swollen stems and/or leaves, and are adapted to dry environments [and so primarily invade arid recipient ecosystems (Anderson 2001)], invasive cacti share several species traits that include large native range size, spread by clonal fragmentation, and spines (Novoa et al. 2016b). They were introduced and spread along similar pathways—historically a few taxa were used for food, fodder, as barrier plants, or for cochineal production; and more recently a wide range of species have been introduced and spread for horticulture (Novoa et al. 2016a). Once introduced, they often have similar invasion dynamics and impacts. Due to the presence of spines and fragments, invasive cacti can attach and disperse via animals, clothes or equipment. The resulting small clumps expand rapidly through vegetative growth, and coalesce to form monocultures, resulting in impenetrable thickets with negative impacts on biodiversity, ecosystem functioning, resource availability, pastoralism, and human health (Novoa et al. 2016b). Moreover, similar management actions are highly effective in controlling invasions of different cactus species (Novoa et al. 2019). Classical biocontrol agents have been used to manage 28 invasive cactus species, achieving complete control in many situations (i.e. no other management interventions are required to reduce impacts to an acceptable level; Zimmermann et al. 2009). The cacti syndrome is potentially extendable for all succulent plants that reproduce vegetatively, have large native ranges and spines, such as Agave species (Badano and Pugnaire 2004).
B. Small aquatic organisms introduced through ballast water in harbors
Ship ballast water is a specific introduction sub-pathway as per the CBD’s scheme, categorized under the broader transport-stowaway pathway category. This pathway explicitly selects for particular species traits, i.e. aquatic organisms with pelagic life stages that are small enough to be taken up into ballast water tanks, and that can survive the journey to a new destination (Briski et al. 2014). Survival during transport often correlates with high levels of phenotypic plasticity. The recipient ecosystems are inevitably boat harbours and can be marine, estuarine or freshwater. As such, the invasion dynamics initially have some similarities, whereby entire pelagic communities are taken up in one or more locations and transferred to a new location. Alien species establish in these artificial habitats and expand from these points of entry. Thus far, there is not enough information on the common impacts of these invasion events. Various response options have been proposed but managing invasions reactively has been found to be mostly infeasible in marine systems. Therefore, the focus has been on preventing introductions through monitoring and pathway management (Ojaveer et al. 2015). Although the Ballast Water Convention came into force in 2017 with the aim of minimizing the biosecurity risk associated with ballast water, it is yet to be fully implemented. Ultimately, it is envisaged that vessels will carry an international ballast water management certificate, demonstrating compliance with ballast management standards, including the use of on-board ballast water treatment units.
C. Large ranid frogs with frequent secondary transfers
Ranoidea is a superfamily of frogs that contain seventeen different families. Ten ranid species within the genera Lithobates and Hoplobatrachus have been recorded as invasive in many climatic zones (including arid regions). These invaders share the same pathways of introduction—intentional introductions for consumption and the pet trade (Tingley et al. 2010), or as contaminants in aquaculture (Mohanty and Measey 2019). Once introduced, they often disperse through natural spread between lentic water bodies, although intentional human-mediated transfers often also occur. In terms of species traits they tend to have large body sizes (> 100 mm snout-vent length). They are environmentally constrained to breeding in static water bodies, and so recipient ecosystems with aquatic impoundments, e.g. agricultural impoundments, can be a prerequisite for an invasion. Invasive large ranid frogs impact native biodiversity by predating on invertebrates and small vertebrates, serving as reservoirs of diseases, and competing with other anurans during the larval stage (Measey et al. 2016). Removal of metamorphs and juveniles was identified to be the best management intervention (Govindarajulu et al. 2005).
D. Piscivorous freshwater fishes in connected aquatic ecosystems
Fishes are among the most widely introduced alien vertebrates and their invasions are a global problem because of their importance in fisheries, aquaculture, recreational fishing and the global pet trade (Cucherousset and Olden 2011; Dawson et al. 2017). Piscivorous freshwater fishes (e.g. rainbow trout Onchorhynchus mykiss, largemouth bass Micropterus salmoides), are mainly introduced through intentional pathways (e.g. for enhancing fisheries) and are mostly released outside of captivity, with high propagule pressure, to provide opportunities for recreational fishing (Cucherousset and Olden 2011). Establishment depends on the interaction between the recipient ecosystem and species traits, including reproductive strategy and physiological tolerance (Marchetti et al. 2004). They primarily invade connected aquatic ecosystems (Marchetti et al. 2004; Ruesink 2005) and inter-catchment movement is dependent on human activity resulting in direct releases, escape or dispersal via infrastructural opportunities, e.g. inter-basin water transfers (Ellender and Weyl 2014).
The impacts of piscivorous freshwater alien fish invasions often include hybridization with native species, introduction of disease, and extirpations of native taxa by direct predation (Cucherousset and Olden 2011). Fishes are extremely difficult to eradicate once established. Methods such as dewatering, manual removal and the use of piscicides are only practical in small and relatively isolated habitats (Britton et al. 2011). Moreover, management of established piscivorous freshwater fishes can be contentious because of conflicting values of stakeholders (Zengeya et al. 2017). For this reason, the management of invasive fishes focusses on preventing further introductions and limiting their spread. To guide this process, considerable research has focussed on developing the widely applied Fish Invasiveness Screening Kit (FISK) which evaluates invasion risk (Copp 2013). Retrospective assessments of the FISK have found the tool relatively robust in predicting successful invaders (Vilizzi et al. 2019).
E. Plant invasions in high elevation areas
Most human-mediated introductions of alien plants are to low- or mid-elevation areas (Alexander et al. 2011; McDougall et al. 2011). As a result, invasive alien plants are rarely mountain specialists, and high elevation areas are generally less invaded than other ecosystems (Chytrý et al. 2008b, 2009; Pauchard et al. 2009). Most invasive plant species in high elevation areas share the samepathways of introduction, since they are species that were initially introduced to low or mid-elevation areas and were then able to spread to higher elevations along roads or other transport corridors, either through their own dispersal or aided by human disturbance, construction, or livestock movements. They also share the species trait of a broad climatic tolerance, which allows them to establish in high elevation recipient ecosystems (Leger et al. 2009; Monty and Mahy 2009; Alexander et al. 2011; Haider et al. 2011; McDougall et al. 2011; Gallien et al. 2016). The outcomes of such invasions are typified by the spread from lower to higher altitudes, with impacts on soil properties and native communities along the way (Alexander et al. 2016). Therefore, when planning the management of plant invasions in high elevations at regional scales, the major goal should be to monitor the presence of alien plants along roadsides and limit their spread (Pauchard and Alaback 2004). This syndrome of “plant invasions in high elevation areas” might be extended to other areas with harsh climates, low propagule pressure and low human populations, such as polar ecosystems.
It is important to note that those species that are specialists in high elevation areas might be intentionally introduced directly to other high elevations (instead of low- or mid-elevations) outside their native range in the future, e.g. through the intensification of agriculture, as ornamental or forestry plants, for the restoration of ski runs, or for managed relocation (McDougall et al. 2011). If this happens and some of the intentionally introduced species become invasive, then the syndrome of “plant invasions in high elevation areas” will become outdated.
F. Tall-statured grasses
Tall-statured grasses include grasses that reach heights of at least 2 m (~8.6% of grasses; 929 species scattered among 21 tribes in seven subfamilies; Canavan et al. 2019). Tall-statured grasses share similar pathways of introduction outside their native range (e.g. for use as biomass feedstock and for bioenergy crops). Moreover, typical species traits that confer tall-statured grasses their invasion success include high biomass production and accumulation, dual reproductive modes, and a generally great anthropogenic interest (Canavan et al. 2017, 2019). Although they can invade different ecosystems (e.g. grasslands, wetlands and forests), invasions by tall-statured grasses often have similar outcomes (e.g. high rates of resource acquisition, competitive exclusion of understory plants, and light reduction) related to their production of biomass. It remains to be assessed whether this group of grasses can be addressed with similar policies or management actions. However, since the traits of invasive tall-statured grasses are very specific, targeted risk assessments should probably be developed. It seems likely that the tall-statured grasses syndrome can be extended to include some other tall species within the order Poales, such as species from the Juncaceae or Cyperaceae families, but this is still to be explicitly tested. Additionally, Canavan et al. (2019) found that species in the subfamily Bambusoideae (woody bamboos; tribes Arundinarieae and Bambuseae) have lower rates of naturalization compared to other tall-statured grasses, seem to invade predominately forest ecosystems post-disturbance, and species within the group typically receive lower risk scores in risk assessments, suggesting that bamboos might be a distinct invasion syndrome.
G. Tree-feeding insects in forests with suitable hosts
Tree-feeding insects are prominent as invasive species. For example, 455 and 400 non-native tree-feeding species are recorded in the USA (Aukema et al. 2010) and Europe (Roques et al. 2016) respectively. They are mainly introduced through unintentional pathways, associated with live plant imports, machinery, roundwood, sawn timber, sea containers, ships or wood packaging materials (Roques 2010). Once introduced, their secondary spread is generally facilitated by anthropogenic dispersal, such as the movement of wood (e.g. wood for campfires or home heating) or attached to live ornamental trees (Brockerhoff and Liebhold 2017). Tree-feeding insect invasions share similar species traits. Asexual reproduction or inbreeding strategies help to avoid mate-finding failure (Liebhold et al. 2016), and most tree-feeding insects are host-specific, i.e. they only feed on their natural hosts or closely related trees. Therefore, introduced species are only able to establish when their recipient ecosystems present suitable hosts (Brockerhoff and Liebhold 2017), and phylogeographic patterns can help to predict host suitability (Gilbert et al. 2012), i.e. the most likely donors of invasive tree-feeding insects are from related biogeographic regions. Once established, they often have similar invasion dynamics. For example, they often exhibit long lag phases between establishment and impact (Roques et al. 2016). These can be due to an initial low habitat suitability, or to the need to reach high densities before their presence and impacts are detected.
Tree-feeding insect invasions are among the greatest threats to forests worldwide (Liebhold et al. 1995), causing highly visible impacts, such as severe defoliation, mortality or reduced growth (Morin and Liebhold 2016). These impacts can, in turn, facilitate non-host tree species, causing changes in tree compositions or indirect effects on other species in the food web. They can also affect other characteristics of the invaded forests, such as carbon and nitrogen flows, carbon sequestration and storage or light penetration. These impacts pose an existential threat to forestry in some areas (Wingfield et al. 2015; Brockerhoff and Liebhold 2017). Finally, by killing street trees and those in gardens, tree-feeding insects can affect both property values and people’s sense of place (Shackleton et al. 2019).
The exchanges of tree-feeding insect species among world regions is increasing, leading to an increasing number of established non-native tree-feeding insect species. For example, in the USA, on average, two new species are detected each year (Aukema et al. 2010). Several specific tools are available to assist with managing such invasions. For example, phylogenetic relationships can help to predict host suitability (Gilbert et al. 2012), i.e. are most likely to come from regions that are biogeographically and climatically similar to the introduced regions. DNA barcoding can help detect immature stages, such as eggs and larvae, at the ports of entry (Ball and Armstrong 2006), and pheromones can help in detecting and monitoring post-border (Myers and Hosking 2002, Suckling et al. 2005).
A method to identify further invasion syndromes
To facilitate the identification of additional invasion syndromes, we propose a systematic method for their circumscription and confirmation (Latombe et al. 2019a), based on the premise that an invasion syndrome should be formed of generalizations that are as broad as possible, but which are still robust and useful (Fig. 3). Often the starting point is to identify invasion events with similar contexts, i.e. pathways, species traits, and characteristics of the recipient ecosystem. However identifying similar outcomes and responses might also be a useful starting point. If the invasion events classified into a putative syndrome vary in the context, outcomes or appropriate responses, then the syndrome needs to be adjusted until it encompasses the invasion events. Putative invasion syndromes should be made progressively more general (i.e. by including more invasion events), and if the context, outcomes, and responses still fit the expanded set of invasion events, then the more general invasion syndrome should be preferred. In other words, the more specific the invasion syndrome, the less useful it is.

Schematic diagram and description of the five steps proposed for identifying invasion syndromes
The invasion syndrome approach thus helps to determine under which situations it is meaningful to generalize, and so make predictions about biological invasions—this is crucial if management lessons are to be shared. We argue that jointly considering groups of invasion events presenting similar management requirements is the only practical way of dealing with the growing numbers of alien species (OEPP/EPPO 2008; van Wilgen et al. 2011). The invasion syndrome approach can facilitate the transfer of lessons between invasion events; for example, transferring insights from Australian Acacia spp. (van Wilgen et al. 2011), Pinus spp. (Nuñez et al. 2017) and Prosopis spp. (Shackleton et al. 2017) between regions of introduction. Moreover, grouping invasion events according to their context and outcomes can identify not only common management goals but also shared stakeholders, thereby potentially simplifying decision-making processes (Novoa et al. 2016a, 2018). We believe that the approach outlined here will provide new insights into the drivers of invasion dynamics; help establish management priorities; and identify more accurate, efficient and transferable management responses. Applying invasion syndromes paves the way for easier sharing of information among stakeholder groups to reveal and identify solutions for new or extant invasions. Incorporating invasion syndromes into decision-making may also help to link practitioners and managers working on different taxa or in regions or ecosystems that, perhaps unknowingly, are actually dealing with similar problems.
Testing the approach
The concept of invasion syndromes remains to be tested empirically. Although data on the characteristics that define the context for an invasion syndrome are becoming more accessible, challenges remain. Information on pathways of introduction and secondary spread is not always available, in particular on their quantitative aspects such as when, how often, or how many individuals are introduced or dispersed. On the other hand, data on species traits are becoming more accessible due to the development of trait databases that encompass a large number of taxa (Supplementary information). For example, the global TraitBank database contains data on more than 330 different traits for 1.7 million species (Parr et al. 2016). However, data quality remains an issue, especially: if the methodology used to measure the traits is not indicated; if traits are not comparable because they are measured differently or in different contexts; if it is unclear whether traits were measured in the native or the alien range; or if trait databases ignore geographic variations in trait values (Yesson et al. 2007; Robertson 2008; Moravcová et al. 2010). Moreover, trait databases often have data for a limited number of species or have many missing values. Such data gaps make it difficult to define a syndrome for a large group of species or invasion events.
Information on the outcomes of invasion events is also becoming increasingly available for a large number of taxa (Zenetos et al. 2005; van Kleunen et al. 2015; Pyšek et al. 2017). For example, the recently released Global Naturalized Flora (GloNAF) database contains information on the distribution of more than 13,000 naturalized alien species in more than 1000 regions of the world (van Kleunen et al. 2015, 2019; Pyšek et al. 2017). The Global Register of Introduced and Invasive Species (GRIIS), supported by the Secretariat of the Convention on Biological Diversity, currently provides checklists of naturalized and invasive species for 20 countries, and aims to soon provide checklists for most countries globally (Pagad et al. 2018). This is, however, a very coarse measure (i.e. naturalized vs. invasive) at relatively large scales (i.e. country level) and only represents a “snapshot in time” since many species are still progressing along the introduction—naturalization—invasion continuum (Pyšek et al. 2012a). GRIIS also conflates the concepts of spread and impact when defining a species at a site as invasive (whereas in reality and practice spread and impact are mediated by different factors). A coordinated monitoring and reporting scheme with standard metrics is clearly needed at the global level (Latombe et al. 2017). More precisely defined categorizations (e.g. the 11 categories proposed by Blackburn et al. 2011 to identify the invasion stage of any invasion event) or population dynamics metrics (Leung et al. 2012), at finer scales, are preferable for characterizing invasion syndromes.
A number of analytical approaches could be used for quantitatively identifying invasion syndromes. As one example, machine learning techniques (see Kelleher et al. 2015 for a detailed review) are possibly the most powerful approaches for identifying invasion syndromes (even with the current limitations). Unsupervised clustering techniques (e.g. hierarchical clustering, k-means clustering, etc.) could be applied to data describing the context, outcomes and response options of invasions (Fig. 1). While this approach ignores the links between the three facets of an invasion syndrome, as shown in Fig. 1, such links could be specified via numerous regressions between multiple response variables (i.e. outcomes and response options) and multiple predictor variables (context) and a clustering method applied to these regressions (Qin and Self 2006). Alternatively, supervised classification techniques could be used (e.g. Random Forest, Support Vector Machines, and Artificial Neural Networks). Data representing the context, outcomes and response options would be collected for a training set of invasion events, which researchers would have already assigned to a predefined invasion syndrome (e.g. cacti with clonal fragmentation in arid ecosystems). Then, a test dataset for a collection of invasion events not yet assigned to any invasion syndrome (e.g. succulents with clonal fragmentation in arid ecosystems) would be fed into the model to determine their possible affiliation to this invasion syndrome. Machine learning techniques are already widely used in invasion science; for example to predict the invasion stage of alien plants using trait and biogeographical data (Chen et al. 2015), to predict eradication success (Xiao et al. 2018), and to identify the source of ballast water using bacterial species composition (Gerhard and Gunsch 2019).
Conclusion
We believe the invasion syndrome approach is a dynamic, composite, and repeatable way of accounting for context-dependencies within invasion science. Its application will facilitate a more mechanistic and predictive understanding of biological invasions, thereby offering better guidelines for management. We suggest that developing and refining invasion syndromes should be a key activity of the “global networks for invasion science” proposed by Packer et al. (2017).
References
Abell R, Thieme ML, Revenga C et al (2008) Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience 58:403–414
Alexander JM, Kueffer C, Daehler CC, Edwards PJ, Pauchard A, Seipel T, MIREN Consortium (2011) Assembly of nonnative floras along elevational gradients explained by directional ecological filtering. Proc Natl Acad Sci USA 108:656–661
Alexander JM, Lembrechts JJ, Cavieres LA et al (2016) Plant invasions into mountains and alpine ecosystems: current status and future challenges. Alpine Bot 126:89–103
Anderson EF (2001) The cactus family. Timber Press, Portland
Aukema JE, McCullough DG, Von Holle B, Liebhold AM, Britton K, Frankel SJ (2010) Historical accumulation of non-indigenous forest pests in the continental US. Biol Sci 60:886–897
Bacher S, Blackburn TM, Essl F et al (2018) Socio-economic impact classification of alien taxa (SEICAT). Meth Ecol Evol 9:159–168
Bacon SJ, Aebi A, Calanca P, Bacher S (2014) Quarantine arthropod invasions in Europe: the role of climate, hosts and propagule pressure. Divers Distrib 20:84–94
Badano EI, Pugnaire FI (2004) Invasion of Agave species (Agavaceae) in south-east Spain: invader demographic parameters and impacts on native species. Divers Distrib 10:493–500
Ball SL, Armstrong KF (2006) A universal DNA-based identification system for insect pests: a test case with the Lymantriidae (Lepidoptera). Can J For Res 36:337–350
Bates AE, McKelvie CM, Sorte CJB, Morley SA, Jones NAR, Mondon JA, Bird TJ, Quinn G (2013) Geographical range, heat tolerance and invasion success in aquatic species. Proc Biol Sci 280:20131958
Binimelis R, Born W, Monterroso I, Rodríguez-Labajos B (2008) Socio-economic impact and assessment of biological invasions. In: Nentwig W (ed) biological invasions. Springer, Berlin, pp 331–347
Blackburn TM, Pyšek P, Bacher S, Carlton JT, Duncan RP, Jarošík V, Wilson JRU, Richardson DM (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26:333–339
Blackburn TM, Essl F, Evans T et al (2014) A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol 12:e1001850
Briski E, Chan FT, MacIsaac HJ, Bailey SA (2014) A conceptual model of community dynamics during the transport stage of the invasion process: a case study of ships’ ballast. Divers Distrib 20:236–244
Britton JR, Gozlan RE, Copp GH (2011) Managing non-native fish in the environment. Fish Fish 12:256–274
Brockerhoff EG, Liebhold AM (2017) Ecology of forest insect invasions. Biol Invasions 19:3141–3159
Cadotte MW, Murray BR, Lovett-Doust J (2006) Evolutionary and ecological influences of plant invader success in the flora of Ontario. Écoscience 13:388–395
Campbell NA (1996) Biology, 4th edn. The Benjamin/Cummings Publishing Company, Riverside
Canavan S, Richardson DM, Visser V, Le Roux JJ, Vorontsova MS, Wilson JRU (2017) The global distribution of bamboos: assessing correlates of introduction and invasion. AoB Plants 9:plw078
Canavan S, Pyšek P, Packer JG et al (2019) Tall-statured grasses: a useful functional group for invasion science? Biol Invasions 21:37–58
Capellini I, Baker J, Allen WL, Street SE, Venditti C (2015) The role of life history traits in mammalian invasion success. Ecol Lett 18:1099–1107
Cassey P, Delean S, Lockwood JL, Sadowski J, Blackburn TM (2018) Dissecting the null model for biological invasions: a meta-analysis of the propagule pressure effect. PLoS Biol 16:e2005987
Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 15:22–40
Catford JA, Vesk PA, Richardson DM, Pyšek P (2012) Quantifying levels of biological invasion: towards the objective classification of invaded and invasible ecosystems. Glob Change Biol 18:44–62
CBD (2014) Pathways of introduction of invasive species, their prioritization and management. https://www.cbd.int/doc/meetings/sbstta/sbstta-18/official/sbstta-18-09-add1-en.pdf
Chen L, Peng S, Yang B (2015) Predicting alien herb invasion with machine learning models: biogeographical and life-history traits both matter. Biol Invas 17:2187–2198
Chytrý M, Jarošík V, Pyšek P, Hájek O, Knollová I, Tichý L, Danihelka J (2008a) Separating habitat invasibility by alien plants from the actual level of invasion. Ecology 89:1541–1553
Chytrý M, Maskell LC, Pino J, Pyšek P, Vilà M, Font X, Smart SM (2008b) Habitat invasions by alien plants: a quantitative comparison among Mediterranean, subcontinental and oceanic regions of Europe. J Appl Ecol 45:448–458
Chytrý M, Pyšek P, Wild J, Pino J, Maskell LC, Vilà M (2009) European map of alien plant invasions, based on the quantitative assessment across habitats. Divers Distrib 15:98–107
Cole E, Keller RP, Garbach K (2019) Risk of invasive species spread by recreational boaters remains high despite widespread adoption of conservation behaviors. J Environ Manag 229:112–119
Copp GH (2013) The Fish Invasiveness Screening Kit (FISK) for non-native freshwater fishes—A summary of current applications. Risk Anal 33:1394–1396
Crawley MJ (1987) What makes a community invasible? In: Gray AJ, Crawley MJ, Edwards PJ (eds) Colonization. Succession and stability. Blackwell, Oxford, pp 429–453
Cucherousset J, Olden JD (2011) Ecological impacts of non-native freshwater fishes. Fisheries 36:215–230
Dawson W, Moser D, van Kleunen M et al (2017) Global hotspots and correlates of alien species richness across taxonomic groups. Nat Ecol Evol 1:0186
Delavaux CS, Weigelt P, Dawson W et al (2019) Mycorrhizal fungi influence global plant biogeography. Nat Ecol Evol 3:424–429
Denslow JS (2003) Weeds in paradise: thoughts on the invasibility of tropical islands. Ann Miss Bot Gard 90:119–127
Díaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Duncan RP, Bomford M, Forsyth DM, Conibear L (2001) High predictability in introduction outcomes and the geographical range size of introduced Australian birds: a role for climate. J Anim Ecol 70:621–632
Duncan RP, Blackburn TM, Rossinelli S, Bacher S (2014) Quantifying invasion risk: the relationship between establishment probability and founding population size. Meth Ecol Evol 5:1255–1263
Ellender BR, Weyl OLF (2014) A review of current knowledge, risk and ecological impacts associated with non-native freshwater fish introductions in South Africa. Aquat Invasions 9:117–132
Elleouet J, Albouy C, Ben Rais Lasram F, Mouillot D, Leprieur F (2014) A trait-based approach for assessing and mapping niche overlap between native and exotic species: the Mediterranean coastal fish fauna as a case study. Divers Distrib 20:1333–1344
Essl F, Bacher S, Blackburn TM et al (2015) Crossing frontiers in tackling pathways of biological invasions. Bioscience 65:769–782
Forsyth DM, Duncan RP, Bomford M, Moore G (2004) Climatic suitability, life-history traits, introduction effort, and the establishment and spread of introduced mammals in Australia. Conserv Biol 18:557–569
Gallardo B, Clavero M, Sánchez MI, Vilà M (2016) Global ecological impacts of invasive species in aquatic ecosystems. Glob Change Biol 22:151–163
Gallien L, Carboni M (2017) The community ecology of invasive species: where are we and what’s next? Ecography 40:335–352
Gallien L, Mazel F, Lavergne S, Renaud J, Douzet R, Thuiller W (2015) Contrasting the effects of environment, dispersal and biotic interactions to explain the distribution of invasive plants in alpine communities. Biol Invasions 17:1407–1423
Gallien L, Thuiller W, Fort N, Boleda M, Alberto FJ, Rioux D, Lainé J, Lavergne S (2016) Is there any evidence for rapid, genetically-based, climatic niche expansion in the invasive common ragweed? PLoS ONE 11:e0152867
Gallien L, Altermatt F, Wiemers M, Schweiger O, Zimmermann NE (2017) Invasive plants threaten the least mobile butterflies in Switzerland. Divers Distrib 23:185–195
Gerhard WA, Gunsch CK (2019) Metabarcoding and machine learning analysis of environmental DNA in ballast water arriving to hub ports. Environ Int 124:312–319
Gilbert GS, Magarey R, Suiter K, Webb CO (2012) Evolutionary tools for phytosanitary risk analysis: phylogenetic signal as a predictor of host range of plant pests and pathogens. Evol Appl 5:869–878
Goodwin BJ, McAllister AJ, Fahrig L (1999) Predicting invasiveness of plant species based on biological information. Conserv Biol 13:422–426
Govindarajulu P, Altwegg R, Anholt BR (2005) Matrix model investigation of invasive species control: bullfrogs on Vancouver Island. Ecol Appl 15:2161–2170
Guo Q, Fei S, Dukes JS, Oswalt CM, Iannone BV III, Potter KM (2015) A unified approach for quantifying invasibility and degree of invasion. Ecology 96:2613–2621
Haider S, Alexander JM, Kueffer C (2011) Elevational distribution limits of non-native species: combining observational and experimental evidence. Plant Ecol Divers 4:363–371
Hawkins CL, Bacher S, Essl F et al (2015) Framework and guidelines for implementing the proposed IUCN Environmental Impact Classification for Alien Taxa (EICAT). Divers Distrib 21:1360–1363
Hayes KR, Barry SC (2008) Are there any consistent predictors of invasion success? Biol Invasions 10:483–506
Hood WG, Naiman RJ (2000) Vulnerability of riparian zones to invasion by exotic vascular plants. Plant Ecol 148:105–114
Hui C, Richardson DM (2017) Invasion dynamics. Oxford University Press, Oxford
Hui C, Richardson DM (2019) How to invade an ecological network. Trends Ecol Evol 34:121–131
Hui C, Richardson DM, Robertson MP, Wilson JRU, Yates CJ (2011) Macroecology meets invasion ecology: linking the native distributions of Australian acacias to invasiveness. Divers Distrib 17:872–883
Hui C, Richardson DM, Pyšek P, Le Roux JJ, Kučera T, Jarošík V (2013) Increasing functional modularity with residence time in the co-distribution of native and introduced vascular plants. Nat Commun 4:2454
Hui C, Richardson DM, Landi P, Minoarivelo HO, Garnas J, Roy HE (2016) Defining invasiveness and invasibility in ecological networks. Biol Invasions 18:971–983
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46:10–18
Hulme PE, Bacher S, Kenis M et al (2008) Grasping at the routes of biological invasions: a framework for integrating pathways into policy. J Appl Ecol 45:403–414
Jeschke JM, Strayer DL (2006) Determinants of vertebrate invasion success in Europe and North America. Glob Change Biol 12:1608–1619
Jeschke JM, Aparicio LG, Haider S, Heger T, Lortie CJ, Pyšek P, Strayer DL (2012) Taxonomic bias and lack of cross-taxonomic studies in invasion biology. Front Ecol Environ 10:349–350
Johnson PT, Olden JD, Vander Zanden MJ (2008) Dam invaders: impoundments facilitate biological invasions into freshwaters. Front Ecol Environ 6:357–363
Kelleher JD, Mac Namee B, D’arcy A (2015) Fundamentals of machine learning for predictive data analytics: algorithms, worked examples, and case studies. MIT Press, Cambridge
Keller RP, Kumschick S (2017) Promise and challenges of risk assessment as an approach for preventing the arrival of harmful alien species. Bothalia 47:1–8
Kikillus KH, Hare KM, Hartley S (2012) Online trading tools as a method of estimating propagule pressure via the pet-release pathway. Biol Invasions 14:2657–2664
Kueffer C (2012) The importance of collaborative learning and research among conservationists from different oceanic islands. Rev Ecol Terre Vie 11:125–135
Kueffer C (2017) Plant invasions in the Anthropocene. Science 358:724–725
Kueffer C, Pyšek P, Richardson DM (2013) Integrative invasion science: model systems, multi-site studies, focused meta-analysis and invasion syndromes. New Phytol 200:615–633
Kull CA, Shackleton CM, Cunningham PJ et al (2011) Adoption, use and perception of Australian acacias around the world. Divers Distrib 17:822–836
Kumschick S, Bacher S, Evans T, Marková Z, Pergl J, Pyšek P, Vaes-Petignat S, van der Veer G, Vilà M, Nentwig W (2015) Comparing impacts of alien plants and animals using a standard scoring system. J Appl Ecol 52:552–561
Lambdon PW, Hulme PE (2006) Predicting the invasion success of Mediterranean alien plants from their introduction characteristics. Ecography 29:853–865
Latombe G, Pyšek P, Jeschke JM, Blackburn TM, Bacher S et al (2017) A vision for global monitoring of biological invasions. Biol Conserv 213:295–308
Latombe G, Richardson DM, Pyšek P, Kučera T, Hui C (2018) Drivers of species turnover vary with species commonness for native and alien plants with different residence times. Ecology 99:2763–2775
Latombe G, Canavan S, Hirsch H et al (2019a) A four-component classification of uncertainties in biological invasions: implications for management. Ecosphere 10:e02669
Latombe G, Roura-Pascual N, Hui C (2019b) Similar compositional turnover but distinct insular biogeographical drivers of native and exotic ants in two oceans. J Biogeogr 46:2299–2310
Lawton JH (1999) Are there general laws in ecology? Oikos 2:177–192
Le Roux JJ, Hui C, Keet JH, Ellis A (2017) Co-introduction versus ecological fitting pathways to the establishment of effective mutualisms during biological invasions. New Phytol 215:1354–1360
Leger EA, Espeland EK, Merrill KR, Meyer SE (2009) Genetic variation and local adaptation at a cheatgrass (Bromus tectorum) invasion edge in western Nevada. Mol Ecol 18:4366–4379
Leung B, Roura-Pascual N, Bacher S et al (2012) TEASIng apart alien species risk assessments: a framework for best practices. Ecol Lett 15:1475–1493
Liebhold AM, MacDonald WL, Bergdahl D, Mastro VC (1995) Invasion by exotic forest pests: a threat to forest ecosystems. For Sci Monogr 30:1–49
Liebhold AM, Yamanaka T, Roques A, Augustin S, Chown SL, Brockerhoff EG, Pyšek P (2016) Global compositional variation among native and non-native regional insect assemblages emphasizes the importance of pathways. Biol Invasions 18:893–905
Lockwood JL, Cassey P, Blackburn TM (2009) The more you introduce the more you get: the role of colonization pressure and propagule pressure in invasion ecology. Divers Distrib 15:904–910
Lockwood JL, Hoopes MF, Marchetti MP (2013) Invasion ecology. Wiley-Blackwell, Chichester
Mahoney PJ, Beard KH, Durso AM, Tallian AG, Long AL, Kindermann RJ, Nolan NE, Kinka D, Mohn HE (2015) Introduction effort, climate matching and species traits as predictors of global establishment success in non-native reptiles. Divers Distrib 21:64–74
Marchetti MP, Moyle PB, Levine R (2004) Invasive species profiling? Exploring the characteristics of non-native fishes across invasion stages in California. Freshw Biol 49:646–661
McDougall KL, Alexander JM, Haider S, Pauchard A, Walsh NG, Kueffer C (2011) Alien flora of mountains: global comparisons for the development of local preventive measures against plant invasions. Divers Distrib 17:103–111
McGeoch MA, Latombe G (2016) Characterizing common and range expanding species. J Biogeogr 43:217–228
McGeoch MA, Genovesi P, Bellingham PJ, Costello MJ, McGrannachan C, Sheppard A (2016) Prioritizing species, pathways, and sites to achieve conservation targets for biological invasion. Biol Invasions 18:299–314
Measey GJ, Vimercati G, de Villiers FA, Mokhatla M, Davies SJ, Thorp CJ, Rebelo AD, Kumschick S (2016) A global assessment of alien amphibian impacts in a formal framework. Divers Distrib 22:970–981
Meyerson LA, Cronin JT, Pyšek P (2016) Phragmites australis as a model organism for studying plant invasions. Biol Invasions 18:2421–2431
Mohanty NP, Measey J (2019) Reconstructing biological invasions using public surveys: a new approach to retrospectively assess spatio-temporal changes in invasive spread. Biol Invasions 21:467–480
Monty A, Mahy G (2009) Clinal differentiation during invasion: Senecio inaequidens (Asteraceae) along altitudinal gradients in Europe. Oecologia 159:305–315
Moodley D, Geerts S, Richardson DM, Wilson JRU (2013) Different traits determine introduction, naturalization and invasion success in woody plants: Proteaceae as a test case. PLoS ONE 8:e75078
Moodley D, Procheş Ş, Wilson JRU (2016) A global assessment of a large monocot family highlights the need for group-specific analyses of invasiveness. AoB Plants 8:plw009
Moravcová L, Pyšek P, Jarošík V, Havlíčková V, Zákravský P (2010) Reproductive characteristics of neophytes in the Czech Republic: traits of invasive and non-invasive species. Preslia 82:365–390
Morin RS, Liebhold AM (2016) Invasive forest defoliator contributes to the impending downward trend of oak dominance in eastern North America. Forestry 89:284–289
Murray KA, Rosauer D, McCallum H, Skerratt LF (2011) Integrating species traits with extrinsic threats: closing the gap between predicting and preventing species declines. Proc R Soc B 278:1515–1523
Myers JH, Hosking G (2002) Eradication. In: Hallman GJ, Schwalbe CP (eds) lnvasive arthropods in agriculture: problems and solutions. Science Publishers, Enfield, pp 293–307
Nentwig W, Kühnel E, Bacher S (2010) A generic impact-scoring system applied to alien mammals in Europe. Conserv Biol 24:302–311
Novoa A, Kaplan H, Kumschick S, Wilson JRU, Richardson DM (2015a) Soft touch or heavy hand? Legislative approaches for preventing invasions: insights from cacti in South Africa. Invas Plant Sci Manag 8:307–316
Novoa A, Le Roux JJ, Robertson MP, Wilson JRU, Richardson DM (2015b) Introduced and invasive cactus species: a global review. AoB Plants 7:plu078
Novoa A, Kaplan H, Wilson JRU, Richardson DM (2016a) Resolving a prickly situation: involving stakeholders in invasive cactus management in South Africa. Environ Manag 57:998–1008
Novoa A, Kumschick S, Richardson DM, Rouget M, Wilson JRU (2016b) Native range size and growth form in Cactaceae predict invasiveness and impact. NeoBiota 30:75–90
Novoa A, Le Roux JJ, Richardson DM, Wilson JRU (2017) Level of environmental threat posed by horticultural trade in Cactaceae. Conserv Biol 31:1066–1075
Novoa A, Shackleton R, Canavan S et al (2018) A framework for engaging stakeholders on the management of alien species. J Environ Manage 205:286–297
Novoa A, Brundu G, Day MD et al (2019) Global actions for managing Cactus invasions. Plants 8:421
Nuñez MA, Chiuffo MC, Torres A et al (2017) Ecology and management of invasive Pinaceae around the world: progress and challenges. Biol Invasions 19:3099–3120
OEPP/EPPO (2008) Recommendations made by EPPO Council in 2007. OEPP/EPPO Bull 38:521–522
Ojaveer H, Galil BS, Campbell ML et al (2015) Classification of non-indigenous species based on their impacts: considerations for application in marine management. PLoS Biol 13:e1002130
Olson DM, Dinerstein E (2002) The Global 200: priority ecoregions for global conservation. Ann Miss Bot Gard 89:199–224
Packer JG, Meyerson LA, Richardson DM et al (2017) Global network for invasion science: benefits, challenges and guidelines. Biol Invasions 19:1081–1096
Padilla DK, Williams SL (2004) Beyond ballast water: aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front Ecol Environ 2:131–138
Pagad S, Genovesi P, Carnevali L, Schiger D, McGeoch M (2018) Introducing the global register of introduced and invasive species. Sci Data 5:170202
Parr CS, Wilson N, Schulz KS, Leary P, Hammock J, Rice J, Corrigan RJ Jr (2016) TraitBank: practical semantics for organism attribute data. Semantic Web 7:577–588
Pauchard A, Alaback PB (2004) Influence of elevation, land use, and landscape context on patterns of alien plant invasions along roadsides in protected areas of south-central Chile. Conserv Biol 18:238–248
Pauchard A, Kueffer C, Dietz H et al (2009) Ain’t no mountain high enough: plant invasions reaching new elevations. Front Ecol Environ 7:479–486
Pergl J, Pyšek P, Bacher S et al (2017) Troubling travellers: are ecologically harmful alien species associated with particular introduction pathways? NeoBiota 32:1–20
Perkins LB, Nowak RS (2013) Invasion syndromes: hypotheses on relationships among invasive species attributes and characteristics of invaded sites. J Arid Land 5:275–283
Potgieter LJ, Richardson DM, Wilson JRU (2014) Casuarina: biogeography and ecology of an important tree genus in a changing world. Biol Invasions 16:609–633
Prins HH, Gordon IJ (2014) Invasion biology and ecological theory: insights from a continent in transformation. Cambridge University Press, Cambridge
Pyšek P, Richardson DM (2006) The biogeography of naturalization in alien plants. J Biogeogr 33:2040–2050
Pyšek P, Richardson DM (2007) Traits associated with invasiveness in alien plants: where do we stand? In: Nentwig W (ed) Biological invasions. Springer, Berlin, pp 97–125
Pyšek P, Cock MJW, Nentwig W, Ravn HP (2007) Ecology and management of giant hogweed (Heracleum mantegazzianum). CAB International, Wallingford
Pyšek P, Jarošík V, Pergl J, Randall R, Chytrý M, Kühn I, Tichý L, Danihelka J, Jun Chrtek J, Sádlo J (2009) The global invasion success of Central European plants is related to distribution characteristics in their native range and species traits. Divers Distrib 15:891–903
Pyšek P, Jarošík V, Hulme PE et al (2010) Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc Natl Acad Sci USA 107:12157–12162
Pyšek P, Danihelka J, Sádlo J et al (2012a) Catalogue of alien plants of the Czech Republic (2nd edition): checklist update, taxonomic diversity and invasion patterns. Preslia 84:155–255
Pyšek P, Jarošík V, Hulme PE, Pergl J, Hejda M, Schaffner U, Vilà M (2012b) A global assessment of invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Glob Change Biol 18:1725–1737
Pyšek P, Pergl J, Essl F et al (2017) Naturalized alien flora of the world: species diversity, taxonomic and phylogenetic patterns, geographic distribution and global hotspots of plant invasion. Preslia 89:203–274
Qin LX, Self SG (2006) The clustering of regression models method with applications in gene expression data. Biometrics 62:526–533
Reichard SH, White P (2001) Horticulture as a pathway of invasive plant introductions in the United States: most invasive plants have been introduced for horticultural use by nurseries, botanical gardens, and individuals. Bioscience 51:103–113
Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1661
Ricciardi A, Atkinson SK (2004) Distinctiveness magnifies the impact of biological invaders in aquatic ecosystems. Ecol Lett 7:781–981
Richardson DM, Pyšek P, Carlton JT (2011) A compendium of essential concepts and terminology in invasion ecology. In: Richardson DM (ed) Fifty years of invasion ecology. The legacy of Charles Elton. Wiley-Blackwell, Chichester, pp 409–420
Riley SPD, Busteed GT, Kats LB, Vandergon TL, Lee LFS, Dagit RG, Kerby JL, Fisher RN, Sauvajot RM (2005) Effects of urbanization on the distribution and abundance of amphibians and invasive species in southern California streams. Conserv Biol 19:1894–1907
Robertson DR (2008) Global biogeographical data bases on marine fishes: caveat emptor. Divers Distrib 14:891–892
Roques A (2010) Alien forest insects in a warmer world and a globalised economy: impacts of changes in trade, tourism and climate on forest biosecurity. N Z J For Sci 40:S77–S94
Roques A, Auger-Rozenberg M-A, Blackburn TM, Garnas JR, Pysˇek P, Rabitsch W, Richardson DM, Wingfield MJ, Liebhold AM, Duncan RP (2016) Temporal and inter- specific variation in rates of spread for insect species invading Europe during the last 200 years. Biol Invasions 18:907–920
Ruesink JI (2005) Global analysis of the factors affecting the outcomes of freshwater fish introductions. Conserv Biol 19:1883–1893
Saul W-C, Roy HE, Booy O, Carnevali L, Chen H-J, Genovesi P, Harrower CA, Hulme PE, Pagad S, Pergl J, Jeschke JM (2017) Assessing patterns in introduction pathways of alien species by linking major invasion databases. J Appl Ecol 54:657–669
Shackleton CM, McGarry D, Fourie S, Gambiza J, Shackleton SE, Fabricius C (2007) Assessing the effects of invasive alien species on rural livelihoods: case examples and a framework from South Africa. Hum Ecol 35:113–127
Shackleton RT, Le Maitre DC, van Wilgen BW, Richardson DM (2017) Towards a national strategy to optimise the management of a widespread invasive tree (Prosopis species; mesquite) in South Africa. Ecosyst Serv 27:242–252
Shackleton RT, Shackleton CM, Kull CA (2019) The role of invasive alien species in shaping local livelihoods and human well-being: a review. J Environ Manag 229:145–157
Shah MA, Reshi ZA, Lavoie C (2011) Predicting plant invasiveness from native range size: clues from the Kashmir Himalaya. J Plant Ecol 5:167–173
Simberloff D (2014) Herbert H. T. Prins and Iain J. Gordon (eds): Invasion biology and ecological theory. Insights from a continent in transformation. Biol Invasions 16:2757–2759
Snow N, Witmer G (2010) American bullfrogs as invasive species: a review of the introduction, subsequent problems, management options, and future directions. In: Timm RM, Fagerstone KA (eds) Proceedings of the 24th vertebrate pest conference. University of California, Davis, pp 86–89
Suckling DM, Gibb AR, Dentener PR, Seldon DS, Clare GK, Jamieson L, Baird D, Kriticos DJ, El-Sayed AM (2005) Urabu Iugens (Lepidoptera: Nolidae) in New Zealand: Pheromone trapping for delimitation and phenology. J Econ Entomol 98:1157–1192
Sun Y, Collins AR, Schaffner U, Müller-Schärer H (2013) Dissecting impact of plant invaders: do invaders behave differently in the new range? Ecology 94:2124–2130
Sun Y, Brönnimann O, Roderick GK, Poltavsky A, Lommen STE, Müller-Schärer H (2017) Climatic suitability ranking of biological control candidates: a biogeographic approach for ragweed management in Europe. Ecosphere 8:e01731
Swart C, Visser V, Robinson TB (2018) Patterns and traits associated with invasions by predatory marine crabs. NeoBiota 39:79–102
Thuiller W, Richardson DM, Pyšek P, Midgley GF, Hughes GO, Rouget M (2005) Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob Change Biol 11:2234–2250
Tingley R, Romagosa CM, Kraus F, Bickford D, Phillips BL, Shine R (2010) The frog filter: amphibian introduction bias driven by taxonomy, body size and biogeography. Glob Ecol Biogeogr 19:496–503
van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13:235–245
van Kleunen M, Dawson W, Essl F et al (2015) Global exchange and accumulation of non-native plants. Nature 525:100–103
van Kleunen M, Pyšek P, Dawson W et al (2019) The Global Naturalized Alien Flora (GloNAF) database. Ecology 100:e02542
van Wilgen BW, Wilson JR (eds) (2018) The status of biological invasions and their management in South Africa in 2017. South African National Biodiversity Institute, Kirstenbosch and DST-NRF Centre of Excellence for Invasion Biology, Stellenbosch
van Wilgen BW, Dyer C, Hoffmann JH, Ivey P, Le Maitre DC, Moore JL, Richardson DM, Rouget M, Wannenburgh A, Wilson JRU (2011) National-scale strategic approaches for managing introduced plants: insights from Australian acacias in South Africa. Divers Distrib 17:1060–1075
Vermonden K, Leuven RS, Van Der Velde G (2010) Environmental factors determining invasibility of urban waters for exotic macroinvertebrates. Divers Distrib 16:1009–1021
Vilà M, Hulme P (2017) Impact of biological invasions on ecosystem services. Springer, Cham
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708
Vilizzi L, Copp GH, Adamovich B et al (2019) A global review and meta-analysis of applications of the freshwater Fish Invasiveness Screening Kit. Rev Fish Biol Fisher. https://doi.org/10.1007/s11160-019-09562-2
Westphal MI, Browne M, MacKinnon K, Noble I (2008) The link between international trade and the global distribution of invasive alien species. Biol Invasions 10:391–398
Wilson JRU, Dormontt EE, Prentis PJ, Lowe AJ, Richardson DM (2009) Something in the way you move: dispersal pathways affect invasion success. Trends Ecol Evol 24:136–144
Wilson JRU, Faulkner KT, Rahlao SJ, Richardson DM, Zengeya TA, van Wilgen BW (2018) Indicators for monitoring biological invasions at a national level. J Appl Ecol 55:2612–2620
Wingfield MJ, Brockerhoff EG, Wingfield BD, Slippers B (2015) Planted forest health: the need for a global strategy. Science 349:832–836
Winston RL, Schwarzländer M, Hinz HL, Day MD, Cock MJW, Julien MH (eds) (2014) Biological control of weeds: a world catalogue of agents and their target weeds, 5th edn. USDA Forest Service, Forest Health Technology Enterprise Team, Morgantown
Xiao Y, Greiner R, Lewis MA (2018) Evaluation of machine learning methods for predicting eradication of aquatic invasive species. Biol Invasions 20:2485–2503
Yesson C, Brewer PW, Sutton T et al (2007) How global is the global biodiversity information facility? PLoS ONE 2:e1124
Zenetos A, Çinar ME, Pancucci-Papadopoulou MA, Harmelin JG, Furnari G, Andaloro F, Bellou N, Streftaris N, Zibrowius H (2005) Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr Mar Sci 6:63–118
Zengeya T, Ivey P, Woodford DJ, Weyl O, Novoa A, Shackleton R, Richardson D, van Wilgen B (2017) Managing conflict-generating invasive species in South Africa: challenges and trade-offs. Bothalia 47:a2160
Zimmermann HG, Moran VC, Hoffmann JH (2009) Invasive cactus species (Cactaceae). In: Muniappan R, Reddy GVP, Raman A (eds) Biological control of tropical weeds using arthropods. Cambridge University Press, Cambridge, pp 108–129
Acknowledgements
This paper originated at a workshop arranged, hosted and funded by the DSI-NRF Centre of Excellence for Invasion Biology (CIB) in Stellenbosch, South Africa, in November 2017. Additional support was provided by the office of the Vice Rector: Research and Postgraduate Studies at Stellenbosch University. AN, DMR, WCS and FAY acknowledge funding from the DSI-NRF Centre of Excellence for Invasion Biology. AN, PP, JČ and JP were supported by Project No. 14-36079G Centre of Excellence PLADIAS (Czech Science Foundation), EXPRO Grant No. 19-28807X (Czech Science Foundation) and long-term research development project RVO 67985939 (The Czech Academy of Sciences). AN, SC, DM and JRUW thank the South African Department of Environment, Forestry, and Fisheries (DEFF) for funding noting that this publication does not necessarily represent the views or opinions of DEFF or its employees. CH is supported by the National Research Foundation of South Africa (Grants 89967 and 109244) and Australian Research Council (Grant DP150103017). DMR, OLFW and JRUW acknowledge support from the National Research Foundation (Grants 85417, 110507 and 85412). FAY acknowledges funding from the Working for Water Programme through their collaborative research project: “Research for Integrated Management of Invasive Alien Species”. FE acknowledges funding by the Austrian Science Foundation FWF (Grant I2086-B16). The attendance of JP at the workshop was supported by Project 17-19025S (Czech Science Foundation). We thank Tim Blackburn, Laure Gallien and Jonathan Jeschke for their participation in the workshop and comments on the manuscript.
Author information
Authors and Affiliations
Contributions
AN, DMR, and JRUW conceived the initial idea based on Kueffer et al. (2013) and organized a workshop where the idea was further developed. AN coordinated the study and wrote the first version of the manuscript with contributions from DMR, JM, JRUW, NPM, OLFW, PP, SC and TBR. All authors participated at the workshop, refined the initial idea, commented on the manuscript and contributed to the final version.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare no competing interests.
Data availability
All data generated or analysed during this study are included in this article (and its Supplementary Information files).
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Novoa, A., Richardson, D.M., Pyšek, P. et al. Invasion syndromes: a systematic approach for predicting biological invasions and facilitating effective management. Biol Invasions 22, 1801–1820 (2020). https://doi.org/10.1007/s10530-020-02220-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-020-02220-w