
Abstract
Gastric cancer (GC) is one of the most common malignancies and its prognosis is extremely poor. This study identifies a novel oncogene, microfibrillar-associated protein 2 (MFAP2) in GC. With integrative reanalysis of transcriptomic data, we found MFAP2 as a GC prognosis-related gene. And the aberrant expression of MFAP2 was explored in GC samples. Subsequent experiments indicated that silencing and exogenous MFAP2 could affect motility of cancer cells. The inhibition of silencing MFAP2 could be rescued by another FAK activator, fibronectin. This process is probably through affecting the activation of focal adhesion process via modulating ITGB1 and ITGA5. MFAP2 regulated integrin expression through ERK1/2 activation. Silencing MFAP2 by shRNA inhibited tumorigenicity and metastasis in nude mice. We also revealed that MFAP2 is a novel target of microRNA-29, and miR-29/MFAP2/integrin α5β1/FAK/ERK1/2 could be an important oncogenic pathway in GC progression. In conclusion, our data identified MFAP2 as a novel oncogene in GC and revealed that miR-29/MFAP2/integrin α5β1/FAK/ERK1/2 could be an important oncogenic pathway in GC progression.
Similar content being viewed by others

Introduction
Gastric cancer (GC) is one of the most common and lethal malignant cancer throughout the world, particularly in Eastern Asian and South American countries1. Surgery is the optimal strategy of treating patients with GC; unfortunately, the application of surgical resection in patients with GC is limited, as most patients are diagnosed at an advanced stage of the disease2. What is more, many cases of GC are also not sensitive to chemotherapy and radiotherapy, making the situation more severe2. Recent years have witnessed the great progress of targeted cancer therapies; however, for GC patients, only trastuzumab, a monoclonal antibody against human epidermal growth factor receptor 2, and ramucirumab, a monoclonal antibody against vascular endothelial growth factor receptor 2, proved to have certain therapeutic effects and are widely applied in clinic3,4. Current treatment regimens for GC are still not adequate.
Researchers are trying to clarify the biological mechanisms underlying tumorigenesis and progression of GC, aiming to provide novel clues to fight against this fatal disease. With the rapid development of high-throughput detection techniques, gene expression data are accumulating rapidly in public repositories and a massive amount of differentially expressed genes (DEGs) between GC and normal tissue has been identified in several studies5,6,7,8,9. Many DEGs have been validated as oncogenes or tumor suppressors, which effect different malignant phenotypes of GC including proliferation, angiogenesis, metastasis, and chemoresistance via activating or inactivating multiple downstream signaling pathways5,6,7,8,9. But owing to different sample resources, experimental techniques, and bioinformatics algorithms, the results among these studies are greatly divergent, and there is still no widely accepted factor dominating the malignant transformation and progression of GC.
Integrative reanalysis of independent transcriptomic data may indicate common and remarkable changes during GC progression. In this study, by integrative analysis of datasets from either Gene Expression Omnibus (GEO) or The Cancer Genome Atlas (TCGA) databases, we successfully unveiled a set of DEGs that were invariably dysregulated in each cohort. Intriguingly, the functions of intersecting DEGs were found to significantly focus on the biological processes, such as extracellular space, extracellular matrix (ECM) organization, extracellular exosome, collagen catabolic process, and ECM–receptor interaction. ECM provides both the structure and signals that modulate biological behavior of cells, and recent studies have established the importance of the remodeling of ECM in cancer progression10,11. Our results implied that matrix remodeling was a hallmark of GC, which was probably underestimated in the past.
To further verify the crucial role of matrix remodeling in GC progression, we conducted survival analysis and obtained 14 genes associated with prognosis of GC patients, including SPARC, MFAP2, SERPINE1, LOX, PDGFRB, OLFML2B, VCAN, COLA18A1, SPON2, COL4A2, CHD11, NRP1, NREP, and COL4A5. Consistent with our expectations, most of them were important components of ECM or important modulators of matrix remodeling. This provided further evidence implying the crucial role of ECM in GC progression.
Among the 14 genes, we were particularly interested in MFAP2 (the microfibrillar-associated protein 2), which is also named microfibril-associated glycoprotein 1 (MAGP1). It is a 183-amino acid protein composed of two domains: a proline- and glutamine-enriched residues in amino terminal half and a 54-amino acid region in carboxy terminal half that targets itself to ECM12,13. Its extracellular form binds to fibrillin, collagen VI, tropoelastin, decorin, and biglycan14, and the intracellular form of MFAP2 upregulated the expression of downstream genes linked to cell adhesion, motility, and matrix remodeling13. Recently, the function of MFAP2 in metabolic disease has attracted a lot of attention. Previous studies demonstrated that, in adipose tissue, MFAP2 had high affinity for members of the transforming growth factor (TGF)-β superfamily, and in the absence of MFAP2, there was an increase in basal TGF-β activity15,16. However, its role in cancer biology is still obscure. In this study, we validated that MFAP2 was upregulated in GC tissue, and it was implicated in the malignant behavior of GC cells, such as proliferation, migration, and invasion. We also demonstrated that it activated focal adhesion kinase (FAK), paxillin, and extracellular signal-regulated kinase 1/2 (ERK1/2) through the MFAP2/integrin α5β1/FAK/ERK1/2 pathway. Furthermore, we explored the mechanisms of its expression dysregulation in GC. Loss of microRNA29 (miR-29) is known to be a mechanism of fibrosis and we found that MFAP2 was a target of miRNA-29 family, and its aberrant high expression was probably due to the absence or inhibition of miR-29 family.
In general, we reveal a set of GC-related genes that are potential diagnostic biomarkers and therapy targets. We also demonstrate that the novel oncogene MFAP2 endows cancer cells by activating integrin signaling. Finally, we provide evidence that miR-29 family members have potential to inhibit MFAP2 and at least partly reverse the aberrant matrix status of GC.
Materials and methods
Study strategy
The workflow of data mining and the number of candidate genes remaining at each step are shown in Fig. 1.

Gastric cancer (GC)-related RNA sequence data were used to screen differentially expressed genes (DEGs) between GC and normal gastric tissues. After taking intersection from different cohorts, DEGs were further screened to identify prognosis-associated genes. Number of candidate genes remaining at each step is shown.
Patients and gene expression data
In this study, five cohorts of patients with GC (GSE29272, GSE79973, GSE62254 and GSE15459 and TCGA) were used for identifying and validating prognostic biomarkers. Description of these cohorts is presented in Supporting Information.
Identification of DEGs
DEGs between matched GC and adjacent normal gastric tissues were identified using TwoClassDif17,18. Briefly, we first filtered DEGs with a fold-change (Tumor/Normal) of >1.5 or <0.67. Next, we confirmed the DEGs with the random variance model-modified t test to reduce statistical errors. Venn diagrams were drawn by online BioVenn website (http://www.biovenn.nl/index.php).
Functional annotation
DAVID database and Gene Ontology (GO) functional and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were used to explore the potential biological function of intersecting DEGs from different cohorts19. P value < 0.01 and false discovery rate (FDR) <0.25 were set as the cutoff criteria.
Identification of prognostic genes
Patients were classified as either high-expression (more than the median expression level of DEGs) or low-expression (less than the median expression level of DEGs) groups according to the expression of intersecting DEGs one by one. Univariate analyses of overall survival (OS) were performed with two-sided log-rank test to compare the differences between the two groups.
Kaplan–Meier plots were made using an online dataset (http://www.kmplot.com)20 with the data of GSE15459 and GSE62254. The analysis was performed using both disease-free survival (DFS) and OS information of patients. The patients were split by median.
Lentivirus transfection
To knockdown the expression of MFAP2, we infected AGS and HGC-27 cells with the MFAP2–short hairpin RNA (shRNA) recombinant lentivirus (Genepharma, Suzhou, China). Detailed protocol of lentivirus transfection is presented in Supporting Information.
Cell proliferation assay
MTT assay was performed using Thiazolyl Blue Tetrazolium Bromide (MTT, M2128, Sigma) following the manufacturer’s recommendations. The cell viability was detected using the multifunctional microplate reader at 490 nm with cells incubated for 2 h at 37 °C. The relative absorbance value was normalized and compared to the control group.
In vitro migration and invasion assays
In the migration assay, cells were plated into the upper chamber of 8-mm-pore-size Transwell chambers (Corning, Corning, NY). Dulbecco's modified Eagle’s medium containing 10% fetal bovine serum was added into the lower chamber. Then the chambers were incubated at 37 °C for 48 h. Cells in the upper chamber were then removed, and the bottom surface of the membranes was counted using 0.1% crystal violet dye. In the invasion assay, matrigel (Clontech, Madison, WI) was used in the Transwell chambers (Corning). Cell migration and invasion were qualified by counting six random fields under a microscope.
Immunofluorescence
AGS cells were grown to confluency on glass coverslips. Cells were fixed with 3.7% paraformaldehyde in phosphate-buffered saline for 20 min. Cells were permeabilized with 0.1% Triton X-100 for 5 min at 4 °C and then blocked with 5% bovine serum albumin in TBST for 1 h. Samples were incubated with primary antibodies overnight for 4 °C and then with appropriate secondary antibodies. Samples were mounted onto slides with mounting medium, and images were acquired using a fluorescence microscope. Images were processed using the Photoshop software (Adobe).
Luciferase assay
Cells were plated into 24-well plates and cotransfected with 200 ng of psiCHECK-2 plasmids and 50 nmol/l of miR-29a (or NC microRNA) for 48 h. Luciferase activities were then measured using Dual-Luciferase Reporter Assay system (Promega, Madison, WI). Renilla luciferase activity was normalized to firefly luciferase activity.
In vivo assays
Animal protocols were approved by the Institutional Animal Care and Use Committee of the Renmin Hospital of Wuhan University. Nude mice (4–5-week old) were raised in an specific pathogen-free environment at the experimental animal center of the Renmin Hospital of Wuhan University. Xenograft tumor growth models were established by subcutaneous injection of MFAP2 knockdown cells and NC cells (2 × 106 cells) into the right dorsal flank. Tumor growth in the nude mice was observed for 28 days. Tumor volume (V, cm3) was evaluated based on tumor length (L) and width (W) with the following formula: V = 1/2 × L × W2. In order to test how MFAP2 affect tumor metastasis, we established metastatic tumor model by giving intravenous tail vein injections of 1 × 105 MFAP2 knockdown cells to two groups of mice. After 7 weeks, the mice were sacrificed, and the tumor nodules formed on the lung and liver surfaces were counted. The tumors were embedded in paraffin for further study. All animal studies were conducted with the approval of the Renmin Hospital of Wuhan University and Use Committee.
Statistical analysis
The correlation between gene expression and the clinicopathologic features was analyzed by Chi-square test using SPSS 20.0 (International Business Machines, Armonk, NY, USA). Three independent experiments were conducted in cellular studies, and results were analyzed using the two-tailed, unpaired Student’s t test. The mean standard deviation (SD) of three independent experiments was determined. Results were expressed as mean ± S.E.M. P < 0.05 was considered statistically significant.
Results
Identification of DEGs in GC
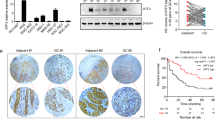
From the expression profile datasets GSE29272 (n = 134), GSE79973 (n = 10), and TCGA (n = 374), we extracted 1352, 2845, and 3453 DEGs, respectively. Two-dimensional hierarchical clustering showed a marked difference of expression modules of the DEGs (Fig. 2a–c). Taking the intersection of DEGs from the three datasets, we extracted 279 genes differently expressed in the GC tissues compared to normal tissues, including 171 upregulated and 108 downregulated genes (Fig. 2d, Table S1).

a–c Using two-dimensional hierarchical clustering, 1352, 2845, and 3453 DEGs were identified from the expression profile datasets GSE29272 (n = 134), GSE79973 (n = 10), and TCGA (n = 374), respectively. d Taking the intersection of DEGs from the three datasets, 279 DEGs were extracted between GC and normal gastric tissues. e DEGs in intersection were mapped onto the DAVID database and subjected to Gene Ontology (GO) functional and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses. GO function and KEGG pathway analysis show that DEGs in intersection are significantly associated with matrix remodeling process. f Using data with clinical information from GSE62254 (n = 300) and TCGA (n = 374), log-rank test was performed to explore the prognostic value of the intersecting DEGs. Ninety-two and 29 genes were closely related to patients’ overall survival in GSE62254 and TCGA, respectively. Taking the intersection of the two datasets, 14 prognostic biomarkers were obtained. g Among the 14 prognostic biomarkers, we are most interested in microfibrillar-associated protein 2 (MFAP2). Kaplan–Meier survival for overall survival (OS) and disease-free survival (DFS) of GC patients was performed. OS (P = 0.009) and DFS (P = 0.008) of GC patients in GSE15459 was significantly negatively associated with the expression of MFAP2. h OS (P = 0.027) and DFS (P = 0.019) of GC patients in GSE62254 was significantly negatively associated with the expression of MFAP2.
The DEGs in intersection are significantly associated with matrix remodeling process
As shown in Fig. 2e, in GO functional analysis, biological processes of the 279 DEGs were found to focus on the extracellular space (P = 1.12 × 10−16), ECM organization (P = 3.20 × 10−15), extracellular exosome (P = 7.84 × 10−15), ECM (P = 5.12 × 10−14), etc. In KEGG pathway analysis, ECM–receptor interaction (P = 3.56 × 10−7), protein digestion and absorption (P = 3.03 × 10−6), cell cycle (P = 1.59 × 10−5), and focal adhesion (P = 1.63 × 10−4) were identified as significant pathways. Collectively, these results implied that the dysregulation of ECM-related proteins are common features in different cohorts.
Identification of prognostic genes among the DEGs
Using data with clinical information from GSE62254 (n = 300) and TCGA (n = 374), we further explored the prognostic value of the 279 DEGs. As shown in Fig. 2f, 92 and 29 genes were closely related to patients’ OS in GSE62254 and TCGA, respectively. Taking the intersection of the two datasets, 14 prognostic biomarkers were obtained (Fig. 2f, Tables 1, 2). Most of the 14 genes were closely related to matrix remodeling, which further supported that matrix remodeling is a crucial character of GC progression. Among the 14 genes, most of them have been reported in GC such as the well-known oncogenes PDGFRB21, VCAN22, and COL18A123, while there were also four genes, MFAP2, OLFML2B, NREP, and COL4A5, that have never been studied in GC. We are especially interested in MFAP2. Kaplan–Meier analysis in cohorts GSE15459 and GSE62254 showed that increased MFAP2 expression revealed poor OS and DFS in GC patients (Fig. 2g, h). Clinical pathology analysis showed that the expression level of MFAP2 was positively correlated with venous invasion and local invasion (Table S2).
MFAP2 is upregulated in GC tissues and cell lines
To validate the aberrant expression of MFAP2 in GC, we first observed the mRNA and protein levels of MFAP2 in 28 and 14 paired GC and adjacent tissues, respectively. As expected, most of the GC tissues exhibited significant upregulation of MFAP2 (Fig. 3a, b). Then we examined the expression level of MFAP2 in different GC cell lines and a gastric epithelial-derived cell line, GES-1. As shown in Fig. 3c, GC cell lines expressed more MFAP2 than GES-1. We also found that a previous study had verified that MFAP2 was upregulated in GC among 168 paired samples by immunohistochemistry24. These findings demonstrated that MFAP2 was upregulated in GC, implying the importance of them in GC pathogenesis.

a Lysates from paired GC and adjacent normal tissues were analyzed by qPCR for the detection of MFAP2. GAPDH was used as a loading control. Each value presents the mean ± S.E.M. of three independent triplicate experiments. b Western blotting analysis of MFAP2 expression in 14 pairs of GC and adjacent tissues. c Western blotting analysis of MFAP2 expression in 4 GC cell lines, namely, HGC-27, SGC-7901, MGC-803, and AGS, and the normal gastric cell line GSE-1. GAPDH was used as a loading control. d The knockdown of MFAP2 in cells was affirmed by western blot. e The knockdown of MFAP2 in cells was affirmed by real-time RT-PCR. RNA sequencing using Illumina HumanHT-12 V4.0 expression beadchip was applied to assess the change of gene expression profile after MFAP2 knockdown in AGS cell line. **P < 0.01 vs. NC. f Cluster analysis of gene expression profile after MFAP2 knockdown. Downregulated (green) and upregulated genes (red) were identified. g Significantly changed pathways were identified based on Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database using the Gene Cloud of Biotechnology Information.
MFAP2 modulates integrin-stimulated FAK activation in GC
In order to elucidate the molecular mechanisms by which MFAP2 influences GC pathogenesis, RNA sequencing was applied to assess the change of gene expression profile after MFAP2 knockdown in AGS cell line. First, we built a stably transfected MFAP2 knockdown cell line and validated it by western and quantitative PCR (qPCR; Fig. 3d, e). As shown by the sequencing, the control groups and MFAP2 knockdown groups were clearly distinguished by hierarchical cluster analysis (Fig. 3f). Five hundred and eighty-one DEGs were screened between sh-MFAP2 and control groups. Pathway analysis indicated that genes regulating focal adhesion were most significantly disrupted after MFAP2 knockdown (enrichment score = 6.82, P = 3.28 × 10−5, FDR = 0.005, Fig. 3g). Together with lamellipodia formation and cell polarization, adhesion to ECM via specific focal adhesion points has long been regarded as an essential step in cancer cell migration and invasion, which is mediated by integrin signal25,26. We noticed that both of ITGB1 and ITGA5 were downregulated after MFAP2 knockdown in RNA sequencing (Fig. 3e). This result was also validated by qPCR (Fig. 4e). Furthermore, western blotting was used to examine the expression of the integrin-stimulated FAK pathway in the MFAP2 Knockdown HGC-27 and AGS cells. As we expected, the expression of ITGB1, ITGA5, FAK, PXN, ERK1/2, PFAK (Tyr397), PPXN (Tyr118), and PERK1/2 (T202/Y204) were all prominently downregulated in the MFAP2 knockdown cells compared with the negative control (Fig. 4f). Based on the above study, we further explored FAK-associated phenotype on MFAP2 knockdown GC cell lines. As shown, HGC-27 and AGS cells displayed a lower cell proliferation rate than control cells after MFAP2 knockdown (Fig. 4b). What is more important, knockdown of MFAP2 suppressed the wound healing (Fig. 4c, d), migration, and invasion rates of AGS and HGC-27 cells (Fig. 4a). FAK is known as a crucial oncogene and promotes cell motility, survival, and proliferation, while in our current results, silencing of MFAP2 markedly inhibited FAK activation27. To explore whether the re-activation of FAK could rescue the effect of MFAP2 silencing, a widely used FAK activator fibronectin was applied28. As shown by immunostaining, the focal adhesion formation was significantly activated or re-activated by fibronectin (Fig. 4g). In the subsequent wound healing assays, AGS cells treated with fibronectin (10 μg/ml) showed stronger motility than the control group even after MFAP2 knockdown (Fig. 4h). Based on these results, we concluded that MFAP2 could possibly modulate integrin-stimulated focal adhesion formation and then work on motility and proliferation. Moreover, the effect of silencing MFAP2 could be rescued by activating FAK and paxillin.

a Cell migration was evaluated by cell wounding assay. b Wound width was quantified in eight fields per dish. c Cell migration and invasion were evaluated by migration and invasion assay. d The effect of MFAP2 knockdown on GC cell proliferation was determined by MTT assay. e Lysates from MFAP2 knockdown and scramble control cells were analyzed by qPCR to validate the downregulation of ITGB1 and ITGA5 after MFAP2 knockdown. GAPDH was used as a loading control. Each value presents the mean ± S.E.M. of three independent triplicate experiments. *P < 0.05 vs. blank, **P < 0.01 vs. blank, ^P < 0.05 vs. control, ^^P < 0.01 vs. control. f Western blotting analysis of MFAP2 knockdown and scramble control cells was performed to validate the downregulation of ITGB1 and ITGA5 and to explore the inhibition effect on FAK and paxillin after MFAP2 knockdown. g Fibronectin as a FAK activator was pre-added into Control and shMFAP2 cells. Immunostaining showed focal adhesion formation (×100, Green: F-actin, Red: pY118paxillin or pY397FAK, Blue: hoechst). h Wound healing assays showed that fibronectin significantly rescue AGS cells motility after MFAP2 knockdown (wound width was quantified in eight fields per dish).
Altering MFAP2 expression has profound effects on tumor growth and metastasis in vivo
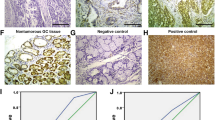
To better demonstrate influence of MFAP2 on proliferation and motility of GC cells, we performed nude mice xenograft assay and tail vein injection assay. For the xenograft assay, nude mice were subcutaneously injected with HGC-27 cells that had stably knocked down MFAP2 or empty vector as control. Tumor volumes were measured every 3 days after inoculation. Hematoxylin and eosin (H&E) staining was used to confirm that the nodules developed in mice were tumors (Table S3). Remarkably, we observed that the tumors formed by MFAP2 knockdown HGC-27 cells grew clearly slower than those formed by control cells (Fig. 5a, b). To evaluate the effects of MFAP2 on tumor metastasis in vivo, two groups of eight mice each were injected intravenously into the tail vein with MFAP2 knockdown or control cells, respectively. After 6 weeks, the mice were sacrificed, and the metastatic nodules in the lung and liver surfaces were counted. A significantly fewer number of metastatic nodules were induced at the surface of the lungs and livers of mice injected with the MFAP2 knocked down cells than in those with the control cells (Fig. 5f, g). H&E staining confirmed that the nodules on the surfaces of mice lungs and livers were metastatic tumors (Fig. 5d, e).

a Image of the xenograft tumors formed in nude mice injected with shRNA silencing of MFAP2 and scrambled shRNA control cells (NC). *P < 0.05. b Volume of xenograft tumors are summarized. c Representative images of metastatic tumor nodules in the lung and liver section of nude mice intravenously injected with MFAP2 knockdown and scrambled shRNA control cells (NC). d, e Representative images of H&E in the lung and liver section of nude mice (original magnification: ×40, calibration bar 125 μm). f, g Number of metastatic tumor nodules in the lung and liver are compared between nude mice injected with MFAP2 knockdown and scrambled shRNA control cells. **P < 0.01, *P < 0.05.
Extracellular MFAP2 also modulates GC cells’ FAK activation
According to previous researches and literatures, MFAP2 was an ECM protein whose function is to help structure elastic fibers16. So we explored whether the extracellular MFAP2 could influence focal adhesion formation. MFAP2 recombinant protein was added to the AGS and HGC-27 cell lines. Notably, treated with MFAP2 recombinant protein greatly enhanced migration and invasion rates of AGS and HGC-27 cells in Transwell assays (Fig. 6a). Then we added MFAP2 into AGS cell line at different concentrations and time gradients. As shown in picture, MFAP2 rapidly activated paxillin, FAK, and ERK1/2 in a time- and concentration-dependent manner (Fig. 6b, c). Immunostaining with pY118paxillin and F-actin showed that AGS cells rapidly made new focal adhesions after treating with MFAP2 recombinant protein (Fig. 6d, e). The expression of integrin α5β1 was reported to be correlated with ERK activity, and as an important downstream kinase of FAK, the activity of ERK1/2 was also closely related to MFAP2 knockdown or exogenous MFAP2 treatment29,30. Therefore, we interrogated whether the integrin expression was modulated in an ERK1/2 activation-dependent way. The AGS cell line was treated with an ERK1/2 inhibitor LY3214996 (200 nM) 2 h before exposure to MFAP2 recombinant protein. As shown, the expression of ITGA5 and ITGB1 significantly increased after MFAP2 treatment, while the expression remained unchanged when ERK1/2 inhibitor was pretreated (Fig. 6f). In conclusion, our results revealed that the extracellular MFAP2 was also strongly correlated with focal adhesion formation and mainly function through integrin α5β1 receptor. Moreover, MFAP2-modulated integrin expression was partially through ERK1/2 activation.

MFAP2 recombinant protein was added into AGS and SGC-7901 cells. a Cell migration and invasion were evaluated by Transwell assay. b The effect of MFAP2 on FAK/paxillin/ERK tyrosine phosphorylation at different time gradients (390 ng/ml for 0, 5, 10, 15, 20, 25 min). *P < 0.05 vs. control. c The effect of MFAP2 on FAK/paxillin/ERK tyrosine phosphorylation at different concentration gradients (0, 130, 260, 390, 520, 650 ng/ml for 25 min). d, e Western blot and immunostaining showed the effect of MFAP2 on control or si ITGA5/ITGB1 AGS cell focal adhesion formation (×100, Green: F-actin, Red: pY118paxillin or pY397FAK, Blue: hoechst). f Western blot showed the effect of MFAP2 on DMSO or LY3214996-treated AGS.
Identification of MFAP2 as a novel target for miR-29
The mechanism of MFAP2 upregulation in GC tissues remained to be solved. We first investigated whether MFAP2 locus is amplified in human cancers using TCGA data on cBioPortal. However, the incidence of homozygous gain of MFAP2 locus was quite low (Fig. S1). We then explored whether the promoter region of MFAP2 was methylated low in human cancers using TCGA data available on MethHC. As shown, the methylation level of MFAP2 promoter was also not significantly decreased in cancer tissue (Fig. S2). Therefore, we focused on miRNA, another key factor contributing to gene expression regulation. Three databases, including TargetScan, miRanda, and miRWalk, were used to search potential microRNAs that are complementary to the 3’-untranslated region (UTR) of MFAP2. Only those miRNAs predicted by all of these methods were considered. One candidate identified was miR-29 family, whose family members miR-29a, miR-29b, and miR-29c all have two complementary sites to 3’-UTR of MFAP2 (located at 4–12 and 51–68, respectively, Fig. 7a, Fig. S3). miR-29 family is downregulated in GC31,32. After transfection with miR-29a, b and c, we observed that MFAP2 was downregulated on both mRNA and protein levels in AGS and HGC-27 cells (Fig. 7b, c). Among the miR-29 family, miR-29a transfection induced the most significant decrease of MFAP2 expression. Cells co-transfected with a miR-29a and wild-type MFAP2 3’-UTR presented a significant decrease in luciferase activity; however, in the mutant groups, much less changes in luciferase activity was observed (Fig. 7d). The luciferase reporter assay suggested that miR-29 family suppressed the transcription of the MFAP2 by targeting MFAP2. Further analysis of GC samples revealed the significant reciprocal association of expression levels between MFAP2 with miR-29a (Fig. 7e). FAK and paxillin are crucial integrin interactors and focal adhesion markers25,33. In AGS and HGC-27 cells, we observed that ITGB1, ITGA5, FAK, pY397FAK, paxillin, and pY118paxillin were downregulated after transfection with miR-29a mimics and upregulated after transfection with miR-29a inhibitors, which mimic the effects of MFAP2 knockdown or overexpression (Fig. 7f). To better visualize the correlation between MFAP2 and miR29a, we transfected AGS with miR-29a or scrambled control, and after 48 h, MFAP2 recombinant protein was added to activate the focal adhesion formation. As shown, treatment with MFAP2 could rescue the inhibition of miR29a on focal adhesion formation (Fig. 7g, h). Collectively, our results indicate that MFAP2 is a direct target of miR-29 family, and its dysregulation may have resulted from the loss of miR-29 family in GC.

a The target sites of miR-29a in 3’-UTR of MFAP2 are shown as a schematic representation. b Real-time RT-PCR showed the expression of MFAP2 mRNA in AGS and HGC-27 cells transfected with miR-29a mimics or inhibitors. **P < 0.01 vs. blank, ^^P < 0.01 vs. control. c Western blotting assays showed the expression of MFAP2 protein in AGS and HGC-27 cells transfected with miR-29a mimics or inhibitors. d Wild-type or mutant 3’-UTR constructs of MFAP2 were cloned into a psi-CHECK2 vector, respectively, and cotransfected with miR-29a mimics in HEK293 cells. Renilla luciferase activities were normalized to firefly luciferase activities. All assays were performed in triplicates and repeated at least three times. ***P < 0.001, **P < 0.01, *P < 0.05. e An inverse correlation was found between miR-29a expression and MFAP2 in GC samples (Spearman’s correlation, P < 0.001, R = −0.758). Each value presents the mean ± S.E.M. of three independent triplicate experiments. ***P < 0.001, **P < 0.01. f Western blotting analysis was performed to examine the expression change of ITGB1, ITGA5, FAK, and paxillin in AGS and HGC-27 cells transfected with miR-29a mimics or inhibitors. g, h Western blot and immunostaining showed that mi29a inhibited activation of FAK and paxillin while this inhibition could be rescued by MFAP2 (Green: F-actin, Red: pY118paxillin or pY397FAK, Blue: hoechst).
Discussion
Gene expression profile data were typically produced on a small scale in most studies, and the list of DEGs from different studies showed distressing inconsistency5,6,7,8. Even though some important “driver genes” in GC have been obtained from DEGs, excessive attention to single gene function may miss important changes of biological characters, which are often determined by a set of genes acting in concert34. Many hallmarks of cancer have been presented such as sustaining proliferation, resistance to cell death, enhancing angiogenesis, and activating metastasis35. In this study, via integrative reanalysis of several expression profile datasets, we identified a novel, generalizable hallmark of GC: matrix remodeling.
In this study, we obtained 14 genes associated with prognosis of GC patients. Most of them participate in matrix remodeling during cancer progression, including MFAP2. MFAP2 plays a vital role in the regulation of integrin signal pathway in cancer cell–ECM interaction. The intracellular form of MFAP2 can induce the transcription of integrin α4 in human osteosarcoma cell line SAOS-213; in vascular development, MFAP2 defect will result in the reduction of integrin–matrix interaction36; our findings highlight the capability of MFAP2 to modify the phenotype of GC cells, partly by upregulating integrin α5β1. In this study, we concluded that MFAP2 might be an activator of integrin/FAK/ERK1/2 signaling in GC progression; however, it may also function importantly in GC carcinogenesis. In this work, there is only preliminary research on the role of MFAP2 in GC and many issues remain to be addressed. MFAP2 may exert oncogenic function by alternative mechanism. MFAP2 is associated with microfibrils in ECM and can also induce the expression of other matrix remodeling genes, such as VCAN13. Predictably, MFAP2 dysregulation will greatly change the status of ECM in cancer microenvironment and further modulate the phenotypes of cancer cells. Intriguingly, MFAP2 was also found to be an unfavorable indicator in multiple other cancers such as liver cancer, pancreatic cancer, renal cancer, and cervical cancer in our unpublished data. The mechanism of how MFAP2 exhibits functions in cancers remains to be studied in detail. In this study, we made a hypothesis that extracellular form of MFAP2 is produced by GC cells and promotes disease progression via autocrine secretion. Protein carriers between cancer cells and ECM such as exosome were potential mediators to transport MFAP2 to cancer microenvironment, and this need to be further explored and validated. Cancer-associated fibroblast is also an important regulator in matrix remodeling and whether it can secrete MFAP2 and participate in the deposition of MFAP2 are also worth studying. What is more, except for MFAP2, among these 14 genes, OLFML2B, NREP, and COL4A5 have not been investigated in GC previously, and they are also candidates for further functional studies.
MiRNA consists of short noncoding sequences that combine with target genes and inhibit gene expression by mRNA translational repression or degradation37. It has been demonstrated that miR-29 is downregulated in most cancers including GC31,38,39,40,41,42. Many downstream genes of miR-29 are matrix remodeling-related genes. For example, miR-29 mediates TGF-β1-induced ECM synthesis through activating Wnt/β-catenin pathway in human pulmonary fibroblasts43; in breast cancer tissues, miR-29 inhibits ECM network genes44; in pancreatic cancer, loss of miR-29 is correlated with a significant increase in ECM deposition45. In this study, we identify that MFAP2 is a direct target gene of miR-29, not only explaining the possible mechanisms of MFAP2 high expression in GC but also helping make the fact more explicit that miR-29 family is the crucial miRNA in matrix remodeling. A single miRNA can regulate multiple genes involved in a biological process. Some researchers think that, compared with conventional pharmacological approaches, the use of miRNA as a therapeutic agent may be more effective in some diseases45. Our results suggest that MFAP2 dysregulation partially resulted from loss of miR-29 family in GC, and miR-29-based therapy is promising in reversing aberrant ECM status of GC.
Conclusion
In conclusion, our findings indicated that matrix remodeling is crucial in the development of GC. This study also provided potential biomarkers and therapy targets for GC and identified miR-29/MFAP2/integrin α5β1/FAK/ERK1/2 as an important oncogenic pathway in GC progression.
Data availability
The datasets analyzed during the current study are available in the GEO repository with accession numbers GSE29272, GSE79973, GSE62254 and GSE15459 [http://www.ncbi.nlm.nih.gov/geo/] and the TCGA repository [https://cancergenome.nih.gov/]. The datasets generated during the current study are available in the GEO repository and assigned accession number GSE114796.
References
Luo, G. et al. Global patterns and trends in stomach cancer incidence: age, period and birth cohort analysis. Int. J. Cancer 141, 1333–1344 (2017).
Sitarz, R. et al. Gastric cancer: epidemiology, prevention, classification, and treatment. Cancer Manag. Res. 10, 239–248 (2018).
Yuan, D. D. et al. Targeted therapy for gastric cancer: current status and future directions (Review). Oncol. Rep. 35, 1245–1254 (2016).
Dreanic, J. et al. Gastric carcinoma at the era of targeted therapies. Curr. Drug Targets 17, 1818–1826 (2016).
Wang, G. et al. Comparison of global gene expression of gastric cardia and noncardia cancers from a high-risk population in china. PLoS ONE 8, e63826 (2013).
Li, W. Q. et al. PLCE1 mRNA and protein expression and survival of patients with esophageal squamous cell carcinoma and gastric adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 23, 1579–1588 (2014).
He, J. et al. Downregulation of ALDOB is associated with poor prognosis of patients with gastric cancer. Onco Targets Ther. 9, 6099–6109 (2016).
Cristescu, R. et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 21, 449–456 (2015).
Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 513, 202–209 (2014).
Kim, S. H., Turnbull, J. & Guimond, S. Extracellular matrix and cell signalling: the dynamic cooperation of integrin, proteoglycan and growth factor receptor. J. Endocrinol. 209, 139–151 (2011).
Wight, T. N. Provisional matrix: a role for versican and hyaluronan. Matrix Biol. 60–61, 38–56 (2017).
Segade, F. 1 et al. Identification of a matrix-binding domain in MAGP1 and MAGP2 and intracellular localization of alternative splice forms. J. Biol. Chem. 277, 11050–11057 (2002).
Segade, F. et al. The intracellular form of human MAGP1 elicits a complex and specific transcriptional response. Int. J. Biochem. Cell Biol. 39, 2303–2313 (2007).
Craft, C. S. MAGP1, the extracellular matrix, and metabolism. Adipocyte 4, 60–64 (2014).
Turecamo, S. E. et al. Contribution of metabolic disease to bone fragility in MAGP1-deficient mice. Matrix Biol. 67, 1–14 (2018).
Craft, C. S. et al. The extracellular matrix protein MAGP1 supports thermogenesis and protects against obesity and diabetes through regulation of TGF-β. Diabetes 63, 1920–1932 (2014).
Clarke, R. et al. The properties of high-dimensional data spaces: implications for exploring gene and protein expression data. Nat. Rev. Cancer 8, 37–49 (2008).
Wright, G. W. & Simon, R. M. A random variance model for detection of differential gene expression in small microarray experiments. Bioinformatics 19, 2448–2455 (2003).
Huang da, W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57 (2009).
Gyorffy, B. et al. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res. Treat. 123, 725–731 (2010).
Guo, Y. et al. Clinicopathological significance of platelet-derived growth factor B, platelet-derived growth factor receptor-β, and E-cadherin expression in gastric carcinoma. Contemp. Oncol. (Pozn.) 17, 150–155 (2013).
Theocharis, A. D. et al. Altered content composition and structure of glycosaminoglycans and proteoglycans in gastric carcinoma. Int. J. Biochem. Cell Biol. 35, 376–390 (2003).
Lee, K. S. et al. Prognostic relevance of collagen XVIII expression in metastatic gastric carcinoma. Tumour Biol. 31, 165–170 (2010).
Wang, J. K. et al. MFAP2 promotes epithelial-mesenchymal transition in gastric cancer cells by activating TGF-β/SMAD2/3 signaling pathway. Onco Targets Ther. 11, 4001–4017 (2018).
Ringer, P. et al. Sensing the mechano-chemical properties of the extracellular matrix. Matrix Biol. 64, 6–16 (2017).
Paluch, E. K., Aspalter, I. M. & Sixt, M. Focal adhesion-independent cell migration. Annu. Rev. Cell Dev. Biol. 32, 469–490 (2016).
Sulzmaier, F. J., Jean, C. & Schlaepfer, D. D. FAK in cancer: mechanistic findings and clinical applications. Nat. Rev. Cancer 14, 598–610 (2014).
Bernard-Trifilo, J. A. et al. Analyzing FAK and Pyk2 in early integrin signaling events. Curr. Protoc. Cell Biol. Chapter 14, Unit 14.7 (2006).
Hu, H. et al. Osteoactivin inhibits dexamethasone-induced osteoporosis through up-regulating integrin β1 and activate ERK pathway. Biomed. Pharmacother. 105, 66–72 (2018).
Xia, M. & Zhu, Y. Fibronectin fragment activation of ERK increasing integrin α5 and β1 subunit expression to degenerate nucleus pulposus cells. J. Orthop. Res. 29, 556–561 (2011).
Gong, J. et al. Characterization of microRNA-29 family expression and investigation of their mechanistic roles in gastric cancer. Carcinogenesis 35, 497–506 (2014).
Wang, Y. et al. Chemotherapy-induced miRNA-29c/Catenin-δ signaling suppresses metastasis in gastric cancer. Cancer Res. 75, 1332–1344 (2015).
Yu, H. G. et al. p190RhoGEF (Rgnef) promotes colon carcinoma tumor progression via interaction with focal adhesion kinase. Cancer Res. 71, 360–370 (2011).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Hanahan, D. & Weinberg, R. A. Hallmarks of cancer: the next generation. Cell 144, 646–674 (2011).
Chen, E., Larson, J. D. & Ekker, S. C. Functional analysis of zebrafish microfibril-associated glycoprotein-1 (Magp1) in vivo reveals roles for microfibrils in vascular development and function. Blood 107, 4364–4374 (2006).
Hu, W. & Coller, J. What comes first: translational repression or mRNA degradation? The deepening mystery of microRNA function. Cell Res. 22, 1322–1324 (2012).
Wang, D. et al. Hsa-miR-21 and Hsa-miR-29 in tissue as potential diagnostic and prognostic biomarkers for gastric cancer. Cell. Physiol. Biochem. 37, 1454–1462 (2015).
Koshizuka, K. et al. Inhibition of integrin β1-mediated oncogenic signalling by the antitumor microRNA-29 family in head and neck squamous cell carcinoma. Oncotarget 9, 3663–3676 (2017).
Jiang, H. et al. Diverse roles of miR-29 in cancer (review). Oncol. Rep. 31, 1509–1516 (2014).
Ru, P. et al. Feedback loop regulation of SCAP/SREBP-1 by miR-29 modulates EGFR signaling-driven glioblastoma growth. Cell Rep. 16, 1527–1535 (2016).
Wu, D. W. et al. c-Myc suppresses microRNA-29b to promote tumor aggressiveness and poor outcomes in non-small cell lung cancer by targeting FHIT. Oncogene 34, 2072–2082 (2015).
Wang, Y. et al. MiR-29 mediates TGFβ 1-induced extracellular matrix synthesis through activation of Wnt/β-catenin pathway in human pulmonary fibroblasts. Technol. Health Care 23, S119–S125 (2015).
Zhu, J. et al. Chaperone Hsp47 drives malignant growth and invasion by modulating an ECM gene network. Cancer Res. 75, 1580–1591 (2015).
Kwon, J. J. et al. Pathophysiological role of microRNA-29 in pancreatic cancer stroma. Sci. Rep. 5, 11450 (2015).
Amy, D. Bradshaw, Diverse biological functions of the SPARC family of proteins. Int. J. Biochem. Cell. Biol. 44, 480–488 (2012).
Chlenski, A. & Cohn, S. L. Modulation of matrix remodeling by SPARC in neoplastic progression. Semin. Cell. Dev. Biol. 21, 55–65 (2010).
Nagaraju, G. P. et al. Molecular mechanisms underlying the divergent roles of SPARC in human carcinogenesis. Carcinogenesis 35, 967–973 (2014).
Thomas, S. L. et al. PTEN augments SPARC suppression of proliferation and inhibits SPARC-induced migration by suppressing SHC-RAF-ERK and AKT signaling. Neuro Oncol. 12, 941–955 (2010).
Seux, M. et al. TP53INP1 decreases pancreatic cancer cell migration by regulating SPARC expression. Oncogene 30, 3049–3061 (2011).
Zhao, Z.-S. et al. SPARC Is Associated with Gastric Cancer Progression and Poor Survival of Patients. Clin. Cancer Res. 16, 260–268 (2010).
Zhang, J. et al. SPARC expression is negatively correlated with clinicopathological factors of gastric cancer and inhibits malignancy of gastric cancer cells. Oncol. Rep. 31, 2312–2320 (2014).
Silveira, N. J. F. et al. Searching for molecular markers in head and neck squamous cell carcinomas (HNSCC) by statistical and bioinformatic analysis of larynx-derived SAGE libraries. BMC Med. Genomics 1, 56 (2008).
Pavón, M. A. et al. uPA/uPAR and SERPINE1 in head and neck cancer: role in tumor resistance, metastasis, prognosis and therapy. Oncotarget 7, 57351–57366 (2016).
Vial, D. & McKeown-Longo, P. J. PAI1 stimulates assembly of the fibronectin matrix in osteosarcoma cells through crosstalk between the αvβ5 and α5β1 integrins. J. Cell Sci. 121, 1661–1670 (2008).
Sato, M. et al. Decreased expression of the plasminogen activator inhibitor type 1 is involved in degradation of extracellular matrix surrounding cervical cancer stem cells. Int. J. Oncol. 48, 829–835 (2016).
Placencio, V. R. & DeClerck, Y. A. Plasminogen Activator Inhibitor-1 in Cancer: Rationale and Insight for Future Therapeutic Testing. Cancer Res. 75, 2969–2974 (2015).
Ju, H. et al. SERPINE1 intron polymorphisms affecting gene expression are associated with diffuse-type gastric cancer susceptibility. Cancer 116, 4248–4255 (2010).
Harris, E. D. Copper-induced activation of aortic lysyl oxidase in vivo. Proc. Natl. Acad. Sci. 73, 371–374 (1976).
Siegel, R. C., Pinnell, S. R. & Martin, G. R. Cross-linking of collagen and elastin. Properties of lysyl oxidase. Biochemistry 9, 4486–4492 (2002).
Xiao, Q. & Ge, G. Lysyl Oxidase, Extracellular Matrix Remodeling and Cancer Metastasis. Cancer Microenviron. 5, 261–273 (2012).
Wang, T.-H., Hsia, S.-N. & Shieh, T.-M. Lysyl Oxidase and the Tumor Microenvironment. Int. J. Mol. Sci. 18, 62 (2017).
Kaneda, A. et al. Lysyl oxidase is a tumor suppressor gene inactivated by methylation and loss of heterozygosity in human gastric cancers. Cancer Res. 64, 6410–6415 (2004).
Kasashima, H. et al. Lysyl oxidase is associated with the epithelial–mesenchymal transition of gastric cancer cells in hypoxia. Gastric Cancer 19, 431–442 (2016).
Jiang, Z. et al. The Role of Platelet-Derived Growth Factor-B/Platelet-Derived Growth Factor Receptor-β Signaling in Chronic Atrial Fibrillation. Cardiology 133, 242–256 (2016).
Paulsson, J., Ehnman, M. & Östman, A. PDGF receptors in tumor biology: prognostic and predictive potential. Future Oncol. 10, 1695–1708 (2014).
Roskoski, R. The role of small molecule platelet-derived growth factor receptor (PDGFR) inhibitors in the treatment of neoplastic disorders. Pharmacol. Res. 129, 65–83 (2018).
Wang, J. X. et al. GLI2 induces PDGFRB expression and modulates cancer stem cell properties of gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 21, 3857–3865 (2017).
Furutani, Y. et al. Identification and characterization of photomedins: novel olfactomedin-domain-containing proteins with chondroitin sulphate-E-binding activity. Biochem. J. 389, 675–684 (2005).
Mukhopadhyay, A. et al. Bioinformatic approaches for identification and characterization of olfactomedin related genes with a potential role in pathogenesis of ocular disorders. Mol Vis. 10, 304–314 (2004).
Wight, T. N. Versican: a versatile extracellular matrix proteoglycan in cell biology. Curr. Opin. Cell Biol. 14, 617–623 (2002).
Carthy, J. M. et al. Versican V1 Overexpression Induces a Myofibroblast-Like Phenotype in Cultured Fibroblasts. PLoS ONE 10, e0133056 (2015).
Ricciardelli, C. et al. The biological role and regulation of versican levels in cancer. Cancer Metastasis Rev. 28(1–2), 233–245 (2009).
Saarela, J. et al. The Short and Long Forms of Type XVIII Collagen Show Clear Tissue Specificities in Their Expression and Location in Basement Membrane Zones in Humans. Am. J. Pathol. 153, 611–626 (1998).
Sánchez-Arévalo Lobo, V. J. et al. Enhanced antiangiogenic therapy with antibody-collagen XVIII NC1 domain fusion proteins engineered to exploit matrix remodeling events. Int. J. Cancer 119, 455–462 (2006).
Heljasvaara, R. et al. Collagen XVIII in tissue homeostasis and dysregulation — Lessons learned from model organisms and human patients. Matrix Biol. 57–58, 55–75 (2017).
He, Y.-W. et al. The extracellular matrix protein mindin is a pattern-recognition molecule for microbial pathogens. Nat. Immunol. 5, 88–97 (2004).
Zhang, Y.-L. et al. SPON2 Promotes M1-like Macrophage Recruitment and Inhibits Hepatocellular Carcinoma Metastasis by Distinct Integrin–Rho GTPase–Hippo Pathways. Cancer Res. 78, 2305–2317 (2018).
Xu, Z. et al. Structure-based design of competitive ligands to target Spon2 in gastric cancer: An integration of molecular modeling and in vitro assay. Bioorg. Chem. 74, 115–121 (2017).
Li, Y. et al. Structure of the F-spondin domain of mindin, an integrin ligand and pattern recognition molecule. EMBO J. 28, 286–297 (2009).
Jeanne, M. et al. COL4A2 Mutations Impair COL4A1 and COL4A2 Secretion and Cause Hemorrhagic Stroke. Am. J. Hum. Genet. 90, 91–101 (2012).
Verbeek, E. et al. COL4A2 mutation associated with familial porencephaly and small-vessel disease. Eur. J. Hum. Genet. 20, 844–851 (2012).
Yoneda, Y. et al. De Novo and Inherited Mutations in COL4A2, Encoding the Type IV Collagen α2 Chain Cause Porencephaly. Am. J. Hum. Genet. 90, 86–90 (2012).
Sugiyama, A., Okada, M., Yamawaki, H. Pathophysiological roles of canstatin on myofibroblasts after myocardial infarction in rats. Eur. J. Pharmacol. 807, 32–43 (2017).
Zhu, J. et al. Six stroma-based RNA markers diagnostic for prostate cancer in European-Americans validated at the RNA and protein levels in patients in China. Oncotarget 6, 16757–16765 (2015).
Xing, Y.-N., Deng, P. & Xu, H.-M. Canstatin induces apoptosis in gastric cancer xenograft growth in mice through the mitochondrial apoptotic pathway. Biosci. Rep. 34, e00106 (2014).
Marchong, M. N. et al. Cdh11 Acts as a Tumor Suppressor in a Murine Retinoblastoma Model by Facilitating Tumor Cell Death. PLoS Genet. 6, e1000923 (2010).
Piao, S. et al. CDH11 inhibits proliferation and invasion in head and neck cancer. J. Oral Pathol. Med. 46, 89–97 (2017).
Zhu, J. et al. The human cadherin 11 is a pro-apoptotic tumor suppressor modulating cell stemness through Wnt/β-catenin signaling and silenced in common carcinomas. Oncogene 31, 3901–3912 (2012).
Raimondi, C. Neuropilin-1 enforces extracellular matrix signalling via ABL1 to promote angiogenesis. Biochem. Soc. Trans. 42, 1429–1434 (2014).
Prud’homme, G. J. & Glinka, Y. Neuropilins are multifunctional coreceptors involved in tumor initiation, growth, metastasis and immunity. Oncotarget 3, 921–939 (2012).
Tomida, C. et al. VEGF pathway-targeting drugs induce evasive adaptation by activation of neuropilin-1/cMet in colon cancer cells. Int. J. Oncol. 52, 1350–1362 (2018).
Li, L. et al. Neuropilin-1 is associated with clinicopathology of gastric cancer and contributes to cell proliferation and migration as multifunctional co-receptors. J. Exp. Clin. Cancer Res. 35, 16 (2016).
Peng, Y. et al. MicroRNA-338 inhibits growth, invasion and metastasis of gastric cancer by targeting NRP1 expression. PLoS ONE 9, e94422 (2014).
Fujitani, M. et al. P311 accelerates nerve regeneration of the axotomized facial nerve. J. Neurochem 91, 737–744 (2004).
McDonough, W. S., Tran, N. L. & Berens, M. E. Regulation of glioma cell migration by serine-phosphorylated P311. Neoplasia 7, 862–872 (2005).
Renieri, A. et al. Major COL4A5 gene rearrangements in patients with juvenile type Alport syndrome. Am. J. Med. Genet. 59, 380–385 (1995).
Gil-Cayuela, C. et al. New Altered Non-Fibrillar Collagens in Human Dilated Cardiomyopathy, Role in the Remodeling Process. PLoS ONE 11, e0168130 (2016).
Petitclerc, E. et al. New functions for non-collagenous domains of human collagen type IV. Novel integrin ligands inhibiting angiogenesis and tumor growth in vivo. J. Biol. Chem. 275, 8051–8061 (2000).
Maeshima, Y. et al. Distinct antitumor properties of a type IV collagen derived from basement membrane. J. Biol. Chem. 275, 21340–21348 (2000).
Acknowledgements
This study was supported by the Natural Science Foundation of China No. 81672387 (granted to Y.-h.G.), No. 81703030 (granted to D.-q.S.), No. 81770899 (granted to Y.-n.Y.) and The guided Fund of Renmin Hospital of Wuhan University, RMYD2018M65 (granted to Z.W.).
Author information
Authors and Affiliations
Contributions
H.-g.Y., Q.-s.D., W.Z. and Y.-n.Y. conceived and designed the experiments; H.-g.Y., Q.-s.D., Y.-n.Y. and W.Z. provided the funds and materials; L.-l.W., L.W., L.-w.Y., K.H., Y.-c.D., D.-m.Y., J.-c.R., G.-g.M., M.X., J.Z. and G.-a.X. performed the experiments; L.-l.W. and Q.-s.D. conducted the bioinformatics and statistical analysis; Q.-s.D. and L.-l.W. wrote this manuscript; L.-w.Y. and L.-h.Z. conducted the revision.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval and consent to participate
This study was approved by the Ethics Committee of Renmin Hospital of Wuhan University and all patients provided informed consent. All procedures were followed in accordance with the ethical standards of the responsible committee on human and experimentation (institutional and national) and with Helsinki Declaration of 1964 and later versions.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
41389_2020_198_MOESM3_ESM.jpg
Supplementary Figure 1. Cross-cancer summary of homozygous mutations and copy number variations of MFAP2 in all cancers available on cBioPortal.
41389_2020_198_MOESM7_ESM.doc
Supplementary Table 2. Correlation between MFAP2 expression and clinicopathological features in 300 patients with GC (GSE62254).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yao, Lw., Wu, Ll., Zhang, Lh. et al. MFAP2 is overexpressed in gastric cancer and promotes motility via the MFAP2/integrin α5β1/FAK/ERK pathway. Oncogenesis 9, 17 (2020). https://doi.org/10.1038/s41389-020-0198-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41389-020-0198-z
This article is cited by
-
Tumor-derived small extracellular vesicles facilitate omental metastasis of ovarian cancer by triggering activation of mesenchymal stem cells
Cell Communication and Signaling (2024)
-
Integrinβ-1 in disorders and cancers: molecular mechanisms and therapeutic targets
Cell Communication and Signaling (2024)
-
Immunosuppressive MFAP2+ cancer associated fibroblasts conferred unfavorable prognosis and therapeutic resistance in gastric cancer
Cellular Oncology (2024)
-
Hippo pathway dysregulation in gastric cancer: from Helicobacter pylori infection to tumor promotion and progression
Cell Death & Disease (2023)
-
Calreticulin enhances gastric cancer metastasis by dimethylating H3K9 in the E-cadherin promoter region mediating by G9a
Oncogenesis (2022)

{kind=link}
{kind=link}
{kind=link}