Impact of Mutations in Arabidopsis thaliana Metabolic Pathways on Polerovirus Accumulation, Aphid Performance, and Feeding Behavior

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Aphid Rearing
2.2. Plant Inoculation with Viruliferous Aphids
2.3. Virus Detection by DAS-ELISA and Quantitative RT-PCR (qRT-PCR)
2.4. Plant to Plant Virus Transmission by M. Persicae
2.5. Myzus Persicae Feeding Behavior Analysis on A. Thaliana Plants
2.6. Myzus Persicae Performance on A. Thaliana Mutants
2.7. Statistical Analyses
3. Results
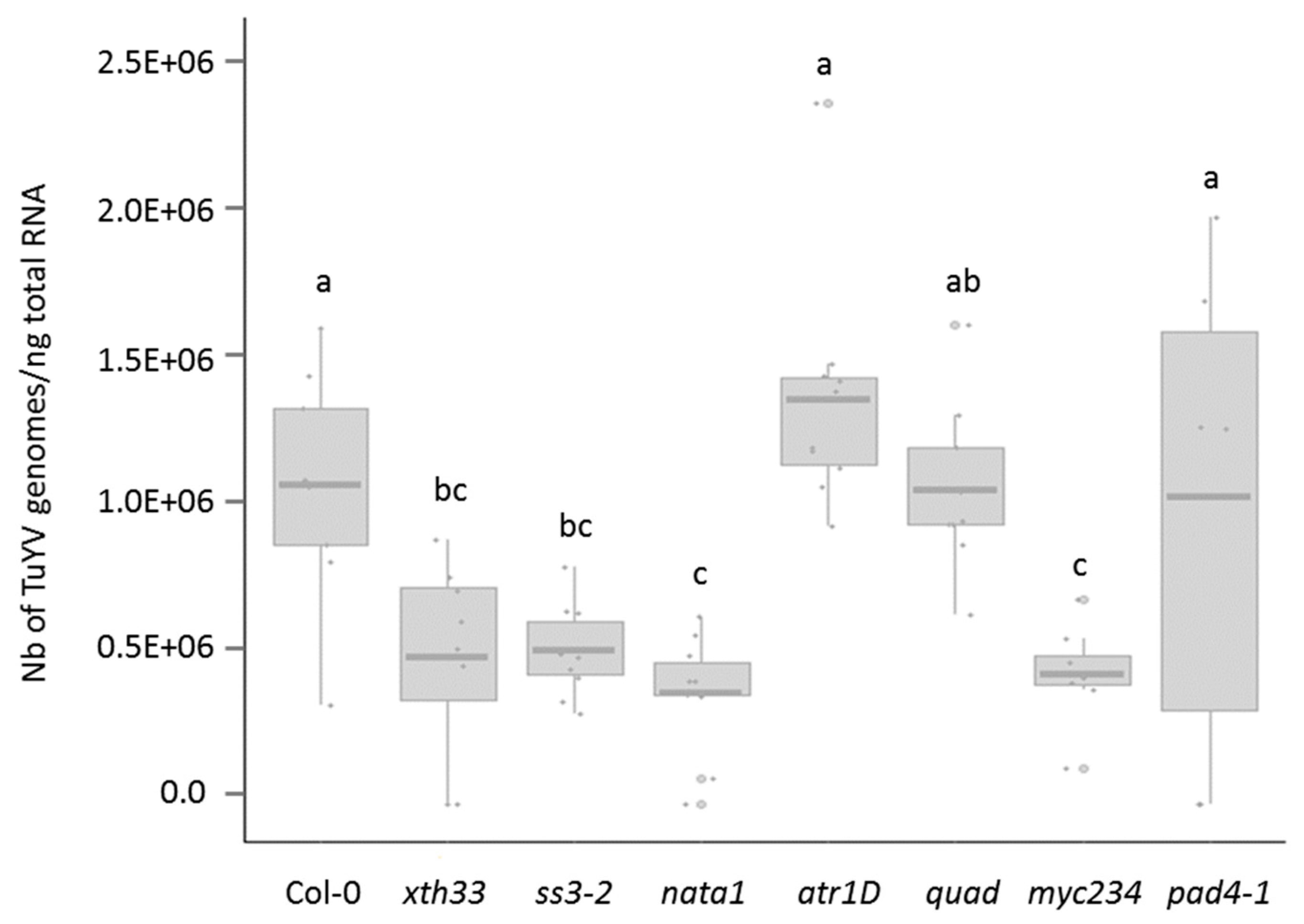
3.1. TuYV Infection Assay of the A. thaliana Mutant Lines
3.2. TuYV Transmission Efficiency by M. persicae from Infected atr1D, quad, myc234, and pad4-1 Mutants
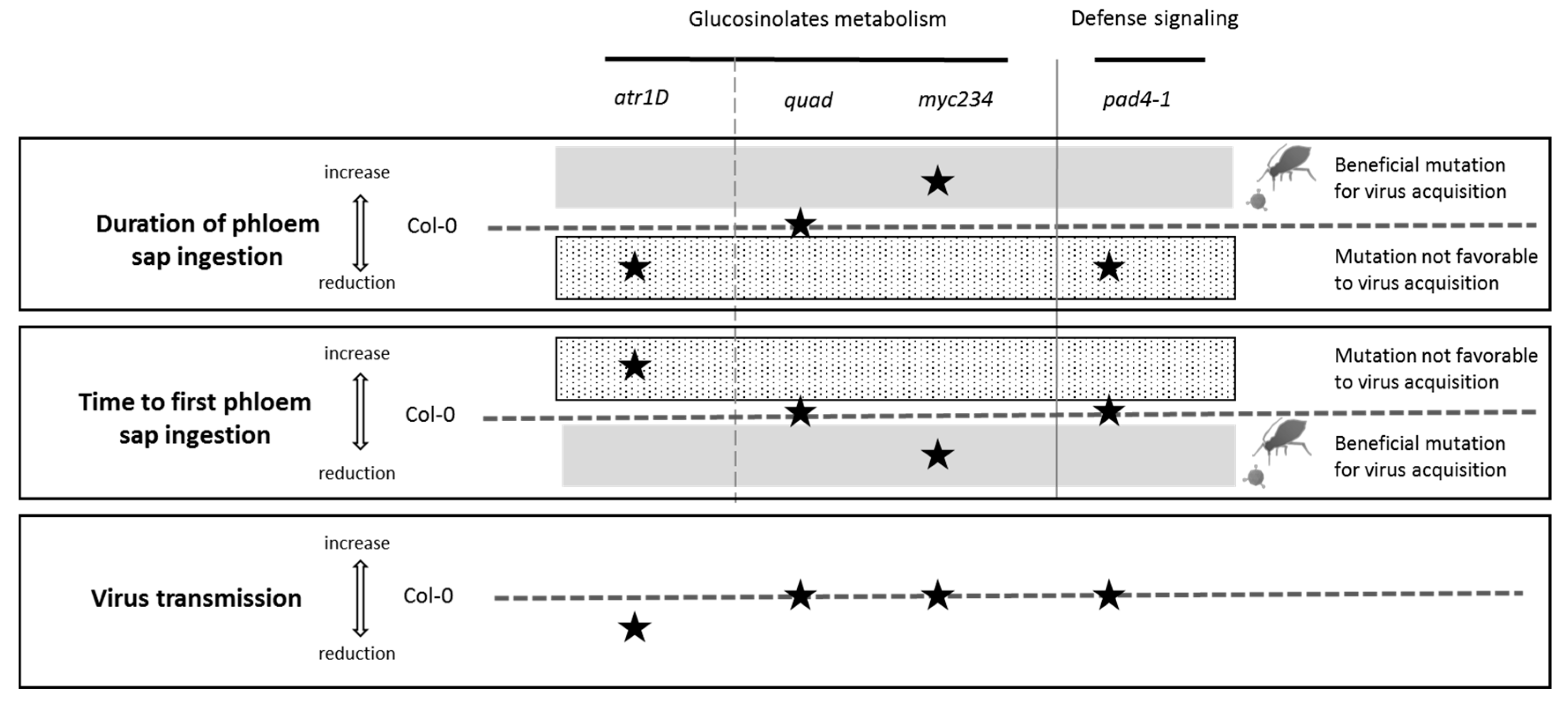
3.3. Effect of the Mutations and TuYV Infection of A. thaliana on Phloem Feeding Activity of M. persicae
3.4. Biomass of Aphids Raised on Non-Infected and TuYV-Infected A. thaliana Mutant Lines
4. Discussion
4.1. Effect of the Mutations and TuYV Infection on Aphid Feeding Activity
4.2. Effect of the Mutations and TuYV Infection on Aphid Biomass
4.3. Impact of the Mutations on TuYV Transmission by Aphids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; Wiley: Chichester, UK, 2000; 466p. [Google Scholar]
- Chaudhary, R.; Atamian, H.S.; Shen, Z.; Briggs, S.P.; Kaloshian, I. GroEL from the endosymbiont Buchnera aphidicola betrays the aphid by triggering plant defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vos, M.; Jander, G. Myzus persicae (green peach aphid) salivary components induce defence responses in Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1548–1560. [Google Scholar] [CrossRef] [PubMed]
- Elzinga, D.A.; De Vos, M.; Jander, G. Suppression of plant defenses by a Myzus persicae (green peach aphid) salivary effector protein. Mol. Plant Microbe Interact. 2014, 27, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Thompson, G.A.; Goggin, F.L. Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Bot. 2006, 57, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Jaouannet, M.; Rodriguez, P.A.; Thorpe, P.; Lenoir, C.J.; MacLeod, R.; Escudero-Martinez, C.; Bos, J.I. Plant immunity in plant-aphid interactions. Front. Plant Sci. 2014, 5, 663. [Google Scholar] [CrossRef] [Green Version]
- Louis, J.; Shah, J. Arabidopsis thaliana-Myzus persicae interaction: Shaping the understanding of plant defense against phloem-feeding aphids. Front. Plant Sci. 2013, 4, 213. [Google Scholar] [CrossRef] [Green Version]
- Nalam, V.; Louis, J.; Shah, J. Plant defense against aphids, the pest extraordinaire. Plant Sci. 2019, 279, 96–107. [Google Scholar] [CrossRef]
- Rodriguez, P.A.; Bos, J.I. Toward understanding the role of aphid effectors in plant infestation. Mol. Plant Microbe Interact. 2013, 26, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Kettles, G.J.; Drurey, C.; Schoonbeek, H.J.; Maule, A.J.; Hogenhout, S.A. Resistance of Arabidopsis thaliana to the green peach aphid, Myzus persicae, involves camalexin and is regulated by microRNAs. New Phytol. 2013, 198, 1178–1190. [Google Scholar] [CrossRef] [Green Version]
- Louis, J.; Leung, Q.; Pegadaraju, V.; Reese, J.; Shah, J. PAD4-dependent antibiosis contributes to the ssi2-conferred hyper-resistance to the green peach aphid. Mol. Plant Microbe Interact. 2010, 23, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Pegadaraju, V.; Louis, J.; Singh, V.; Reese, J.C.; Bautor, J.; Feys, B.J.; Cook, G.; Parker, J.E.; Shah, J. Phloem-based resistance to green peach aphid is controlled by Arabidopsis PHYTOALEXIN DEFICIENT4 without its signaling partner ENHANCED DISEASE SUSCEPTIBILITY1. Plant J. 2007, 52, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jander, G. Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a deterrent indole glucosinolate. Plant J. 2007, 49, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, B.W.; Schroeder, F.C.; Jander, G. Identification of indole glucosinolate breakdown products with antifeedant effects on Myzus persicae (green peach aphid). Plant J. 2008, 54, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Mewis, I.; Tokuhisa, J.G.; Schultz, J.C.; Appel, H.M.; Ulrichs, C.; Gershenzon, J. Gene expression and glucosinolate accumulation in Arabidopsis thaliana in response to generalist and specialist herbivores of different feeding guilds and the role of defense signaling pathways. Phytochemistry 2006, 67, 2450–2462. [Google Scholar] [CrossRef]
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. C. R. Biol. 2010, 333, 524–538. [Google Scholar] [CrossRef]
- Mulot, M.; Monsion, B.; Boissinot, S.; Rastegar, M.; Meyer, S.; Bochet, N.; Brault, V. Transmission of Turnip yellows virus by Myzus persicae Is Reduced by Feeding Aphids on Double-Stranded RNA Targeting the Ephrin Receptor Protein. Front. Microbiol. 2018, 9, 457. [Google Scholar] [CrossRef] [Green Version]
- Gildow, F. Luteovirus transmission mechanisms regulating vector specificity. In The Luteoviridae; Smith, H.G., Barker, H., Eds.; CAB International: Oxon, UK, 1999; pp. 88–111. [Google Scholar]
- Dader, B.; Then, C.; Berthelot, E.; Ducousso, M.; Ng, J.C.K.; Drucker, M. Insect transmission of plant viruses: Multilayered interactions optimize viral propagation. Insect Sci. 2017, 24, 929–946. [Google Scholar] [CrossRef]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Evolutionary determinants of host and vector manipulation by plant viruses. Adv. Virus Res. 2018, 101, 189–250. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Semiz, G.; Blande, J.D. Life-history strategies affect aphid preference for yellowing leaves. Biol. Lett. 2009, 5, 603–605. [Google Scholar] [CrossRef] [Green Version]
- Bruyère, A.; Brault, V.; Ziegler-Graff, V.; Simonis, M.T.; Van den Heuvel, J.F.; Richards, K.; Guilley, H.; Jonard, G.; Herrbach, E. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging and on virus accumulation, symptoms, and aphid transmission. Virology 1997, 230, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Chay, C.; Gildow, F.E.; Gray, S.M. Readthrough protein associated with virions of barley yellow dwarf luteovirus and its potential role in regulating the efficiency of aphid transmission. Virology 1995, 206, 954–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudel, P.; Chesnais, Q.; Fouche, Q.; Krieger, C.; Halter, D.; Bogaert, F.; Meyer, S.; Boissinot, S.; Hugueney, P.; Ziegler-Graff, V.; et al. The Aphid-Transmitted Turnip yellows virus Differentially Affects Volatiles Emission and Subsequent Vector Behavior in Two Brassicaceae Plants. Int. J. Mol. Sci. 2018, 19, 2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eigenbrode, S.D.; Ding, H.; Shiel, P.; Berger, P.H. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae). Proc. Biol. Sci. 2002, 269, 455–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmo-Sousa, M.; Moreno, A.; Plaza, M.; Garzo, E.; Fereres, A. Cucurbit aphid-borne yellows virus (CABYV) modifies the alighting, settling and probing behaviour of its vector Aphis gossypii favouring its own spread. Ann. Appl. Biol. 2016, 169, 284–297. [Google Scholar] [CrossRef]
- Chesnais, Q.; Couty, A.; Uzest, M.; Brault, V.; Ameline, A. Plant infection by two different viruses induce contrasting changes of vectors fitness and behavior. Insect Sci. 2019, 26, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Perez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 578. [Google Scholar] [CrossRef] [Green Version]
- Rajabaskar, D.; Bosque-Perez, N.A.; Eigenbrode, S.D. Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 2014, 186, 32–37. [Google Scholar] [CrossRef]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef]
- Hillung, J.; Garcia-Garcia, F.; Dopazo, J.; Cuevas, J.M.; Elena, S.F. The transcriptomics of an experimentally evolved plant-virus interaction. Sci. Rep. 2016, 6, 24901. [Google Scholar] [CrossRef] [Green Version]
- Whitham, S.A.; Yang, C.; Goodin, M.M. Global impact: Elucidating plant responses to viral infection. Mol. Plant Microbe Interact. 2006, 19, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Mauck, K.E.; Kenney, J.; Chesnais, Q. Progress and challenges in identifying molecular mechanisms underlying host and vector manipulation by plant viruses. Curr. Opin. Insect Sci. 2019, 33, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Patton, M.F.; Bak, A.; Sayre, J.; Heck, M.; Casteel, C.L. A polerovirus, Potato Leafroll virus, alters plant-vector interactions using three viral proteins. Plant Cell Environ. 2020, 2020 43, 387–399. [Google Scholar] [CrossRef]
- Guo, H.; Gu, L.; Liu, F.; Chen, F.; Ge, F.; Sun, Y. Aphid-borne Viral Spread Is Enhanced by Virus-induced Accumulation of Plant Reactive Oxygen Species. Plant Physiol. 2019, 179, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, A.; Patton, M.F.; Perilla-Henao, L.M.; Aegerter, B.J.; Casteel, C.L. Ethylene signaling mediates potyvirus spread by aphid vectors. Oecologia 2019, 190, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Casteel, C.L.; De Alwis, M.; Bak, A.; Dong, H.; Whitham, S.A.; Jander, G. Disruption of Ethylene Responses by Turnip mosaic virus Mediates Suppression of Plant Defense against the Green Peach Aphid Vector. Plant Physiol. 2015, 169, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteel, C.L.; Yang, C.; Nanduri, A.C.; De Jong, H.N.; Whitham, S.A.; Jander, G. The NIa-Pro protein of Turnip mosaic virus improves growth and reproduction of the aphid vector, Myzus persicae (green peach aphid). Plant J. 2014, 77, 653–663. [Google Scholar] [CrossRef]
- Bak, A.; Cheung, A.L.; Yang, C.; Whitham, S.A.; Casteel, C.L. A viral protease relocalizes in the presence of the vector to promote vector performance. Nat. Commun. 2017, 8, 14493. [Google Scholar] [CrossRef]
- Van den Heuvel, J.F.; Boerma, T.M.; Peters, D. Transmission of potato leafroll virus from plants and artificial diets by Myzus persicae. Phytopathology 1991, 81, 150–154. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Membrane potentials as an indication for plant cell penetration by aphid stylets. Entomol. Exp. Appl. 1985, 38, 187–193. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Hogen Esch, T. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Divol, F.; Vilaine, F.; Thibivilliers, S.; Kusiak, C.; Sauge, M.H.; Dinant, S. Involvement of the xyloglucan endotransglycosylase/hydrolases encoded by celery XTH1 and Arabidopsis XTH33 in the phloem response to aphids. Plant Cell Environ. 2007, 30, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Myers, A.M.; James, M.G. Mutations affecting starch synthase III in Arabidopsis alter leaf starch structure and increase the rate of starch synthesis. Plant Physiol. 2005, 138, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adio, A.M.; Casteel, C.L.; De Vos, M.; Kim, J.H.; Joshi, V.; Li, B.; Juery, C.; Daron, J.; Kliebenstein, D.J.; Jander, G. Biosynthesis and defensive function of Ndelta-acetylornithine, a jasmonate-induced Arabidopsis metabolite. Plant Cell 2011, 23, 3303–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Calvo, P.; Chini, A.; Fernandez-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, F.; Fernandez-Calvo, P.; Zander, M.; Diez-Diaz, M.; Fonseca, S.; Glauser, G.; Lewsey, M.G.; Ecker, J.R.; Solano, R.; Reymond, P. Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 2013, 25, 3117–3132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.Y.; Sonderby, I.E.; Halkier, B.A.; Jander, G.; de Vos, M. Non-volatile intact indole glucosinolates are host recognition cues for ovipositing Plutella xylostella. J. Chem. Ecol. 2009, 35, 1427–1436. [Google Scholar] [CrossRef]
- Celenza, J.L.; Quiel, J.A.; Smolen, G.A.; Merrikh, H.; Silvestro, A.R.; Normanly, J.; Bender, J. The Arabidopsis ATR1 Myb transcription factor controls indolic glucosinolate homeostasis. Plant Physiol. 2005, 137, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Louis, J.; Shah, J. Plant defence against aphids: The PAD4 signalling nexus. J. Exp. Bot. 2015, 66, 449–454. [Google Scholar] [CrossRef]
- Pegadaraju, V.; Knepper, C.; Reese, J.; Shah, J. Premature leaf senescence modulated by the Arabidopsis PHYTOALEXIN DEFICIENT4 gene is associated with defense against the phloem-feeding green peach aphid. Plant Physiol. 2005, 139, 1927–1934. [Google Scholar] [CrossRef] [Green Version]
- Singh, V. Studies on Plant-Aphid Interactions: A Novel Role for Trehalose Metabolism in Arabidopsis Defense Against Green Peach Aphid. Dissertation, University of North Texas, Denton, TX, USA, 2012; 136p. [Google Scholar]
- Wu, D.; Qi, T.; Li, W.X.; Tian, H.; Gao, H.; Wang, J.; Ge, J.; Yao, R.; Ren, C.; Wang, X.B.; et al. Viral effector protein manipulates host hormone signaling to attract insect vectors. Cell Res. 2017, 27, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.D.; Woldemariam, M.G.; Mescher, M.C.; Jander, G.; De Moraes, C.M. Glucosinolates from Host Plants Influence Growth of the Parasitic Plant Cuscuta gronovii and Its Susceptibility to Aphid Feeding. Plant Physiol 2016, 172, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Bender, J.; Fink, G.R. A Myb homologue, ATR1, activates tryptophan gene expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 5655–5660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolamiol-Becet, D.; Monsion, B.; Chapuis, S.; Hleibieh, K.; Scheidecker, D.; Alioua, A.; Bogaert, F.; Revers, F.; Brault, V.; Ziegler-Graff, V. Phloem-Triggered Virus-Induced Gene Silencing Using a Recombinant Polerovirus. Front. Microbiol. 2018, 9, 2449. [Google Scholar] [CrossRef] [PubMed]
- Kos, M.; Houshyani, B.; Achhami, B.B.; Wietsma, R.; Gols, R.; Weldegergis, B.T.; Kabouw, P.; Bouwmeester, H.J.; Vet, L.E.; Dicke, M.; et al. Herbivore-mediated effects of glucosinolates on different natural enemies of a specialist aphid. J. Chem. Ecol. 2012, 38, 100–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kos, M.; Houshyani, B.; Wietsma, R.; Kabouw, P.; Vet, L.E.; van Loon, J.J.; Dicke, M. Effects of glucosinolates on a generalist and specialist leaf-chewing herbivore and an associated parasitoid. Phytochemistry 2012, 77, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofius, D.; Herbers, K.; Melzer, M.; Omid, A.; Tacke, E.; Wolf, S.; Sonnewald, U. Evidence for expression level-dependent modulation of carbohydrate status and viral resistance by the potato leafroll virus movement protein in transgenic tobacco plants. Plant J. 2001, 28, 529–543. [Google Scholar] [CrossRef]
- Madsen, S.R.; Kunert, G.; Reichelt, M.; Gershenzon, J.; Halkier, B.A. Feeding on Leaves of the Glucosinolate Transporter Mutant gtr1gtr2 Reduces Fitness of Myzus persicae. J. Chem. Ecol. 2015, 41, 975–984. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Number | Mutant | Gene Name | Effects on M. persicae Observed on the Corresponding Mutant | References |
|---|---|---|---|---|
| At1g10550 | xth33 | xyloglucan:xyloglucosyl transferase33 | Settling preference. | [44] |
| At1g11720 | ss3-2 | starch synthase III | Reduced population. | [45,53] |
| At2g39030 | nata1 | Gcn5-related N-acetyltransferase | Reduced aphid progeny when Nδ-acetylornithine was added to an artificial medium or when transiently produced in leaves. No effect on aphids when NATA1 is expressed in transgenic plants. | [46] |
| At1g32640 At5g46760 At4g17880 | myc234 | MYC2, MYC3, MYC4 | Higher aphid attractiveness. | [47,54] |
| At2g22330 At4g39950 At5g61420 At5g07690 | quad | glucosinolate quadruple mutant | No increase of aphid population. | [49,55] |
| At5g60890 | atr1D | altered tryptophan regulation 1 | Decreased aphid reproduction at the rosette stage. No increase of aphid population. | [50,55,56] |
| At3g52430 | pad4-1 | phytoalexin-deficient 4 | Better feeding activity and enhanced susceptibility to aphids. | [12,52] |
| Virus Source Plant | Exp. 1 1 | Exp. 2 2 | Exp. 3 2 | Exp. 4 3 |
|---|---|---|---|---|
| atr1D | 23/60 4 (38.33%) a 5 | 26/45 (57.70%) a | / | 41/60 (68.33%) a |
| Quad | 43/60 (71.67%) b | 41/45 (91.11%) b | / | 48/60 (80.00%) b |
| myc234 | / | / | 16/20 (80.00%) | |
| pad4-1 | 43/60 (71.67%) b | / | / | 42/60 (70.00%) a |
| Col-0 | 49/60 (81.67%) b | 42/43 (93.33%) b | 18/20 (90.00%) | 53/60 (88.33%) b |
| Col-0 | xth33 | ss3-2 | nata1 | atr1D | quad | myc234 | pad4-1 | ||
|---|---|---|---|---|---|---|---|---|---|
| s_E2 (min) 2 | n1 | 19 | 18 | 20 | 18 | 21 | 20 | 22 | 20 |
| non-inf. | 199.54 ± 30.02 ab | 254.72 ± 33.76 ab | 274.05 ± 30.95 a | 129.09 ± 23.14 b | 183.04 ± 27.83 ab | 292.55 ± 26.18 ab | 277.90 ± 20.16 a | 165.82 ± 24.88 ab | |
| TuYV-infected | 229.09 ± 30.74 AB | 268.44 ± 27.88 A | 227.06 ± 23.97AB | 182.61 ± 36.73 AB | 139.60 ± 33.79 B | 154.84 ± 28.87 AB | 265.82 ± 26.07 A | 132.28 ± 24.63 B | |
| non-inf. versus inf. | ns | ns | ns | ns | ns | ns | ns | ns | |
| t > 1E2 (min) 3 | n | 18 | 22 | 20 | 17 | 18 | 20 | 20 | 20 |
| non-inf. | 167.14 ± 23.47 ab | 134.49 ± 35.50 ab | 104.73 ± 16.21 ab | 181.67 ± 31.40 a | 179.15 ± 22.98 ab | 108.19 ± 21.26 ab | 130.94 ± 17.95 b | 167.43 ± 28.20 ab | |
| TuYV-infected | 162.55 ± 25.99 AB | 105.71 ± 14.00 B | 131.53 ± 25.95 B | 287.20 ± 31.67 A | 218.47 ± 36.48 A | 218.12 ± 39.56 AB | 108.96 ± 17.86 B | 108.34 ± 25.38 AB | |
| non-inf. versus inf. | ns | * | ns | ns | * | ** | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogaert, F.; Marmonier, A.; Pichon, E.; Boissinot, S.; Ziegler-Graff, V.; Chesnais, Q.; Villeroy, C.; Drucker, M.; Brault, V. Impact of Mutations in Arabidopsis thaliana Metabolic Pathways on Polerovirus Accumulation, Aphid Performance, and Feeding Behavior. Viruses 2020, 12, 146. https://doi.org/10.3390/v12020146
Bogaert F, Marmonier A, Pichon E, Boissinot S, Ziegler-Graff V, Chesnais Q, Villeroy C, Drucker M, Brault V. Impact of Mutations in Arabidopsis thaliana Metabolic Pathways on Polerovirus Accumulation, Aphid Performance, and Feeding Behavior. Viruses. 2020; 12(2):146. https://doi.org/10.3390/v12020146
Chicago/Turabian StyleBogaert, Florent, Aurélie Marmonier, Elodie Pichon, Sylvaine Boissinot, Véronique Ziegler-Graff, Quentin Chesnais, Claire Villeroy, Martin Drucker, and Véronique Brault. 2020. "Impact of Mutations in Arabidopsis thaliana Metabolic Pathways on Polerovirus Accumulation, Aphid Performance, and Feeding Behavior" Viruses 12, no. 2: 146. https://doi.org/10.3390/v12020146