 A. Pedro Gonçalves
A. Pedro Gonçalves Karen M. Chow1
Karen M. Chow1 Sara Cea-Sánchez
Sara Cea-Sánchez N. Louise Glass
N. Louise Glass- 1Department of Plant and Microbial Biology, University of California, Berkeley, Berkeley, CA, United States
- 2Departamento de Genética, Universidad de Sevilla, Sevilla, Spain
- 3Environmental Genomics and Systems Biology Division, Lawrence Berkeley National Laboratory, Berkeley, CA, United States
The formation of the fungal mycelial network is facilitated by somatic cell fusion of germinating asexual spores (or germlings). Neurospora crassa germlings in close proximity display chemotropic growth that is dependent upon an intracellular network of mitogen-activated protein kinase (MAPK) signaling cascades. Approximately 80 genes involved in intercellular communication and fusion have been identified, including three mutants with similar morphological phenotypes: Δwhi-2, Δcsp-6, and Δamph-1. Here we show that WHI-2 localizes to the cell periphery and regulates endocytosis, mitochondrial organization, sporulation, and cell fusion. WHI-2 was required to transduce signals through a conserved MAPK pathway (NRC-1/MEK-2/MAK-2) and target transcription factors (PP-1/ADV-1). The amph-1 locus encodes a Bin/Amphiphysin/Rvs domain-containing protein and mis-expression of whi-2 compensated for the cell fusion and endocytosis deficiencies of a Δamph-1 mutant. The csp-6 locus encodes a haloacid dehalogenase phosphatase whose activity was essential for cell fusion. Although fusion-deficient with themselves, cells that lacked whi-2, csp-6, or amph-1 showed a low frequency of chemotropic interactions with wild type cells. We hypothesize that WHI-2 could be important for signal perception during chemotropic interactions via a role in endocytosis.
Introduction
Fungi can sense their surroundings, receive environmental cues, interpret them, and respond accordingly. During asexual growth, germinated asexual spores (germlings) and hyphae of the ascomycete fungus Neurospora crassa display social behaviors that trigger the process of cell-cell communication and somatic cell fusion. Cell fusion creates a mycelial network that allows the circulation of nutrients, water and cellular elements including genetic material throughout an interconnected colony (Leeder et al., 2011; Fischer and Glass, 2019). Somatic cell fusion in N. crassa operates in an analogous way to somatic cell fusion events in mammalian systems, including during muscle, placenta, and bone tissue development (Hernández and Podbilewicz, 2017).
In N. crassa, somatic cell fusion of germlings is initiated when two cells (which can be genetically identical) undergo chemotropic interactions via the exchange of yet-to-be discovered signals by forming polarized cellular protrusions called conidial anastomosis tubes (CATs) (Gabriela Roca et al., 2005). After CATs from germlings come into contact, a switch from cell growth to cell wall dissolution is initiated (Gonçalves et al., 2019), followed by plasma membrane merger and cytoplasmic continuity; nuclear fusion is not a consequence of somatic cell fusion (Leeder et al., 2011). The combination of polar growth, hyphal branching, and cell fusion events is vital for the formation of the interconnected multicellular hyphal network that is the hallmark growth habit of filamentous fungi (Glass et al., 2004).
The availability of a well-annotated genome (Galagan et al., 2003; Borkovich et al., 2004) and a near full genome deletion strain collection (Colot et al., 2006) have enabled screening of deletion mutants for cell fusion phenotypes in N. crassa (Fu et al., 2011). Deletion strains affected in cell communication and fusion often show a reduction in aerial hyphae extension and flat-like growth and approximately 80 genes have been found to play a role in these processes in N. crassa. The functions of these genes have been partially characterized and range from intracellular signaling, calcium modulation, membrane merger, production of reactive oxygen species, actin regulation, vesicle trafficking and transcriptional control (Glass et al., 2004; Leeder et al., 2011; Fischer and Glass, 2019). Two conserved mitogen-activated protein kinase (MAPK) modules form the core of the intracellular signal transduction network that is activated during cell fusion: the SOFT (SO)/MIK-1/MEK-1/MAK-1 cascade—that is part of the cell wall integrity pathway—and the STE-50/STE-20/HAM-5/NRC-1/MEK-2/MAK-2 module (Fischer and Glass, 2019). During chemotropic growth that precedes cell fusion, components of both of these pathways are recruited to the tip of germling CATs and to the tips of fusion hyphae (Fleissner et al., 2009b; Dettmann et al., 2012, 2014; Jonkers et al., 2014). The tip recruitment and displacement of the MAK-2 signaling module to a single fusion tip occurs every ~8–10 min and alternates at CAT and fusion tips of chemotropic partners in a perfectly out of phase manner with localization of SO, resembling a ping-pong mechanism of signal sending and receiving (Fleissner et al., 2009b; Leeder et al., 2011; Serrano et al., 2018). These two pathways are required for the activation of the transcription factors PP-1 and ADV-1 that function as master regulators of cell fusion genes (Fischer et al., 2018).
Despite recent advances, the function of many cell fusion genes remains unclear. In particular, whi-2, the N. crassa ortholog of the Saccharomyces cerevisiae gene WHISKEY2 (WHI2), has been shown to be required for cell fusion (Fu et al., 2014). The S. cerevisiae Whi2 was initially identified as a central player in the coordination between cell proliferation and nutrient availability. Loss-of-function mutations in WHI2 result in cells that cannot properly sense the extracellular nutritional status and fail to shift from exponential to stationary growth (Saul and Sudbery, 1985), leading to cells that are smaller than normal due to cell division without sustained cell growth. Yeast whi2 mutants also show actin cytoskeleton disorganization, increased cell death, aberrant mitochondrial morphology and defects in executing endocytosis (Binley et al., 1999; Care et al., 2004; Leadsham et al., 2009). Additionally, whi2 mutants show an increase in the activity of the Ras/cAMP/PKA pathway, permitting the downstream general stress response transcription factor Msn2/4 to remain phosphorylated and outside of the nucleus, where it is unable to activate the expression of cell cycle arrest genes (Radcliffe et al., 1997a; Leadsham et al., 2009; Sadeh et al., 2011). Overexpression of WHI2 results in filamentous growth that is dependent on Ste11 and partially dependent on Ste7, Ste20 and Ste12 (Radcliffe et al., 1997b). These proteins are orthologs of N. crassa NRC-1 (Ste11), MEK-2 (Ste7), STE-20 (Ste20), and PP-1 (Ste12), components of a MAPK signaling complex and its target transcription factor that are required for somatic cell fusion in a number of filamentous fungi (Fischer and Glass, 2019). The S. cerevisiae WHI2 gene also seems to be a hot spot for adaptive mutations, indicating a central role for the respective protein in fungal development and environmental responses (Cheng et al., 2008; Gresham et al., 2008; Kvitek and Sherlock, 2013; Lang et al., 2013; Teng et al., 2013; Szamecz et al., 2014; Treusch et al., 2015; Payen et al., 2016; Comyn et al., 2017).
In this study, we examined the role of the N. crassa WHI-2 during fungal development. Our findings indicate that WHI-2 localized to the cell periphery, affected endocytosis and mitochondrial morphology, and functioned upstream of the NRC-1/MEK-2/MAK-2/PP-1/ADV-1 signaling pathway to activate cell-cell communication. Additionally, we discuss two other cell fusion proteins, CSP-6 and AMPH-1, whose functions are related to WHI-2. CSP-6 showed a localization pattern similar to WHI-2 and its deletion phenocopied the morphological and cell fusion defects of a Δwhi-2 strain. AMPH-1 is an endocytosis regulator whose absence was compensated for by the mis-expression of WHI-2. This work provides new details on genes/proteins that function upstream of a key MAPK signaling module.
Materials and Methods
Strains and Culture Media
Standard procedures for the handling of N. crassa cells were employed. Cells were grown in Vogel's minimal medium (VMM) plus 2% (w/v) sucrose and 1.5% (w/v) agar (Vogel, 1956). Crosses were performed on synthetic cross medium (Westergaard and Mitchell, 1947). Wild type and deletion strains are available from the Fungal Genetics Stock Center (FGSC) (McCluskey et al., 2010) and were constructed as part of the Neurospora Genome Project (Colot et al., 2006). Strains used in this study are listed in Table S1. For all the experiments, the indicated strains were grown for ~7 days in VMM-containing slant tubes and a conidial suspension was obtained by adding sterile ddH2O into tubes, vortexing and passing the mixture through cheesecloth to remove hyphal fragments. Hygromycin B (Thermo Fisher Scientific, Waltham, MA, USA) was used at a final concentration of 200 μg/ml.
Strain Construction
The whi-2, csp-6, and amph-1 genes were amplified from genomic DNA of the FGSC2489 wild type strain using primers ACCTCTAGAATGGCTGCCGCGGGAGGAG and CAGTTAATTAAACGCAGTCCAATCACACTCATCTCC for whi-2 (creating XbaI/PacI restriction sites), primers TTTTACTAGTATGAGCAACTCGAACCCG and TTTTTTAATTAAAAGAGTGACGTCCAGAACCAG for csp-6 (creating SpeI/PacI restriction sites) and primers TTTTTTAATTAAAAGAGTGACGTCCAGAACCAG and CGGTTAATTAAAACAGTTCCGCTGATACTC for amph-1 (creating XbaI/PacI restriction sites). The resulting PCR products were cloned into pCR-Blunt II-TOPO, excised using the respective combination of restriction enzymes and inserted into pMF272 (GenBank accession number: AY598428.1) (Freitag et al., 2004). In pMF272, whi-2, csp-6 and amph-1 were placed downstream of a ccg-1 promoter and upstream of sgfp and a ccg-1 terminator. For construction of the CSP-6D284A strain, site-directed mutagenesis using primers ccttgttctaGCTttggatgaaa and tttcatccaaAGCtagaacaagg were used. These primers were used in a Pfu-based PCR using the pMF272 vector carrying the wild type csp-6 (described above) as the DNA template that was followed by treatment with the DpnI restriction enzyme. The resulting vectors were transformed into Δwhi-2; his-3, Δcsp-6; his-3 and Δamph-1; his-3 conidia, respectively, using a Gene Pulser electroporator (Bio-Rad, Hercules, CA, USA) at 1.5 kV, 25 μF, 600 Ω. Homokaryotic strains expressing the indicated gfp-tagged genes were obtained by backcrosses. Sanger sequencing to confirm that there were no irregularities in the constructs was performed at the UC Berkeley DNA Sequencing Facility.
Microscopy
Conidia were diluted to a concentration of 1.5 × 107 cells/ml. For the evaluation of conidial morphology, 10 μl were pipetted onto a glass slide and covered with a coverslip. To examine communication frequency, 80 μl of conidial suspension was spread onto 5 cm VMM agar plates. In co-culture experiments two strains were mixed in a 1:1 proportion before plating. Staining with FM4-64 (N-(3-triethylammoniumpropyl)-4-(4-diethylaminophenylhexatrienyl) pyridinium dibromide; Thermo Fisher Scientific, Waltham, MA, USA) was carried out by incubating 2 μM FM4-64 in a 1.5 × 107 cells/ml conidial suspension in a total volume of 500 μl for 15 min in the dark; the cells were then washed twice with ddH2O and resuspended in 500 μl to reestablish the initial spore concentration. The plates were briefly dried in a fume hood and incubated at 30°C, in the dark for 3.5–4 h or 16–20 h to analyze cells at the germling or hyphal stage, respectively. Squares of ~1 cm were excised and observed. For the analysis of the accumulation of endocytic intermediates, 15 μl of 4 μM FM4-64 was added to the agar slice immediately before imaging. Conidial morphology and cell communication were assessed using a Zeiss Axioskop 2 using a 40x Plan-Neofluor oil immersion objective lens. The percentage of cell communication was determined by counting the relative frequency of cell pairs that displayed a chemotropic behavior when germinated conidia were within ~15 μm of each other. For the FM4-64 microscopy assay, as well as WHI-2, CSP-6, AMPH-1, ARG-4, MAK-2 and SO localization studies, a Leica SD6000 confocal microscope equipped with a Yokogawa CSU-X1 spinning disk head, 488 nm and 561 nm lasers and a 100 × 1.4 N.A. oil-immersion objective lens controlled by Metamorph (Molecular Devices, LLC, San Jose, CA, USA) was used. Images were analyzed using ImageJ (Schneider et al., 2012). Multiple cells were analyzed per experiment and representative examples are shown.
Flow Cytometry
In order to examine if plasma membrane material was being appropriately guided to the vacuoles by endocytosis, we adapted a previously published method (Zheng et al., 1998). Conidia at a concentration of 106/ml were inoculated into glass tubes containing 1.5 ml VMM without agar and grown at 30°C, 200 rpm, for a total duration of 4 hrs; 2 μM FM4-64 and 10 μM 7-amino-4-chloromethylcoumarin (CMAC Blue; Thermo Fisher Scientific, Waltham, MA, USA) were added to the cultures for the last 45 and 15 min, respectively. The conidia were harvested by centrifugation (5,000 rpm, 5 min, 4°C) and washed twice with cold 1x PBS before being resuspended in cold 1x PBS and at least 10,000 events acquired on a BD LSRFortessa X-20 flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). The fluorescence of FM4-64 was recorded using a 488 nm laser and 685LP (710/50 nm) filter; the fluorescence of CMAC Blue was recorded using a 355 nm laser and (515/30 nm) filter. FlowJo (FlowJo, LLC, Ashland, OR, USA) was used for analyses.
Bioinformatics and Statistical Analysis
The presence of conserved domains was assessed using InterProScan 5 (Jones et al., 2014). The WHI-2/Whi2p amino acid alignment was edited using BoxShade (https://embnet.vital-it.ch/software/BOX_form.html). The percentage of identity and similarity was calculated using the Sequence Manipulation Suite (http://www.bioinformatics.org/sms2/ident_sim.html). Statistical significance was tested by ANOVA followed by a Tukey post-hoc test using Prism (GraphPad Software, San Diego, CA, USA). At least three independent experiments were performed for all data shown in this paper. The schematic model of the function of WHI-2, CSP-6, and AMPH-1 was built using BioRender (app.biorender.com).
Results
WHI-2 Is Epistatic to the NRC-1/MEK-2/MAK-2 Signaling Pathway
The N. crassa WHI-2 (NCU10518) is a 297 amino acid protein that harbors two BTB (Broad-complex, Tramtrack, and Bric-à-brac)/POZ (POx virus and Zinc finger) conserved domains (Figure S1). These domains have been shown to be involved in protein-protein interactions during multiple cellular processes ranging from ion channel assembly and gating, actin dynamics, transcriptional regulation to chromatin remodeling (Perez-Torrado et al., 2006). A recent report suggested that whi-2 homologs might be distant relatives of members of the human disease-related potassium channel tetramerization domain (KCTD) protein family (Teng et al., 2018). N. crassa WHI-2 shows 24% identity and 33.5% similarity to S. cerevisiae Whi2p, but is substantially shorter (297 aa compared to 486 aa) (Figure S1). Previous deletion strain screenings in N. crassa identified multiple genes, including whi-2, as being required for germling fusion (Chinnici et al., 2014; Fu et al., 2014). In addition to the germling fusion defect (Figures 1A,B), the Δwhi-2 deletion strain displayed additional morphological defects (Fu et al., 2014), such as an impairment in the formation of aerial hyphae (Figure S2), production of unseparated proconidial chains (Figure 1C) and lack of hyphal fusion (Figure 1D). The introduction of an epitope-tagged whi-2 allele (whi-2-gfp) in the Δwhi-2 mutant complemented the germling and hyphal fusion (Figures 1A,B,D), conidial separation (Figure 1C) and aerial hyphae (Figure S2) developmental defects. The Δwhi-2 mutant grew slower than the wild type strain (Figure 1E) and produced profuse spores across a Petri plate, as compared to the wild type strain that sporulated mainly around the perimeter of the plate (Figure 1F). The Δwhi-2 strain also did not produce protoperithecia when grown on synthetic cross medium, as shown in a previous deletion collection screening (Fu et al., 2014). Fertility was restored in Δwhi-2; whi-2-gfp cells and this strain was used as a female in crosses in subsequent experiments (Table S1).
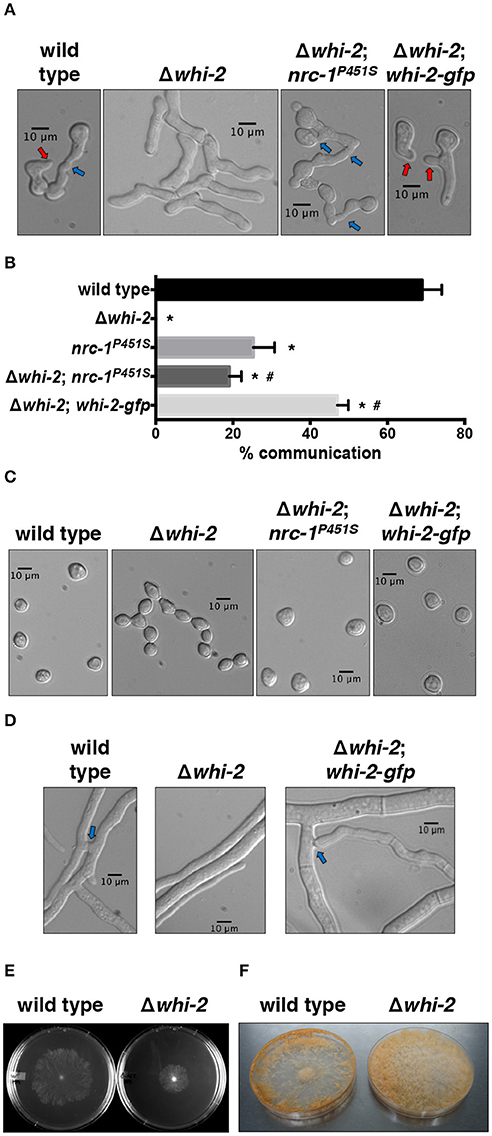
Figure 1. Cell fusion and conidiogenesis defects observed in a Δwhi-2 mutant can be partially rescued by the introduction of a constitutively active allele of nrc-1. (A,B) Cell-cell communication was analyzed by observing brightfield microscopy (A) of closely positioned germlings 4 h post-inoculation. Red and blue arrows indicate conidial anastomosis tubes (CATs) and cell communication/fusion events, respectively. The relative frequency of cell-cell communication is shown in (B). Error bars represent the standard deviation. *p-value < 0.0001 vs. wild type; #p-value < 0.001 vs. Δwhi-2. (C) Conidial morphology was assessed in WT and whi-2 strains using brightfield microscopy. Note the presence of proconidial chains in the Δwhi-2 strain. (D) Hyphal fusion was evaluated 16 h post-inoculation. Blue arrows point to hyphal fusion events. (E,F) Radial growth (E) and sporulation (F) were analyzed in wild type and Δwhi-2 colonies 24 h or 7 days after inoculation on Petri dishes, respectively.
Existing data suggests that Δwhi-2 cells can communicate with wild type partner cells at a low frequency (Fu et al., 2014), a phenotype that we confirmed in Δwhi-2/wild type germling pairs (Figure 2A). When MAK-2-GFP or SO-GFP were expressed in the Δwhi-2 mutant, recruitment of both proteins to the CATs of Δwhi-2 cells undergoing chemotropic interactions with the wild type cells was observed (Figure 2B). Thus, the absence of WHI-2 did not directly affect the ability to form CATs nor the recruitment of MAK-2 and SO to communicating cell tips in wild type + Δwhi-2 pairings.
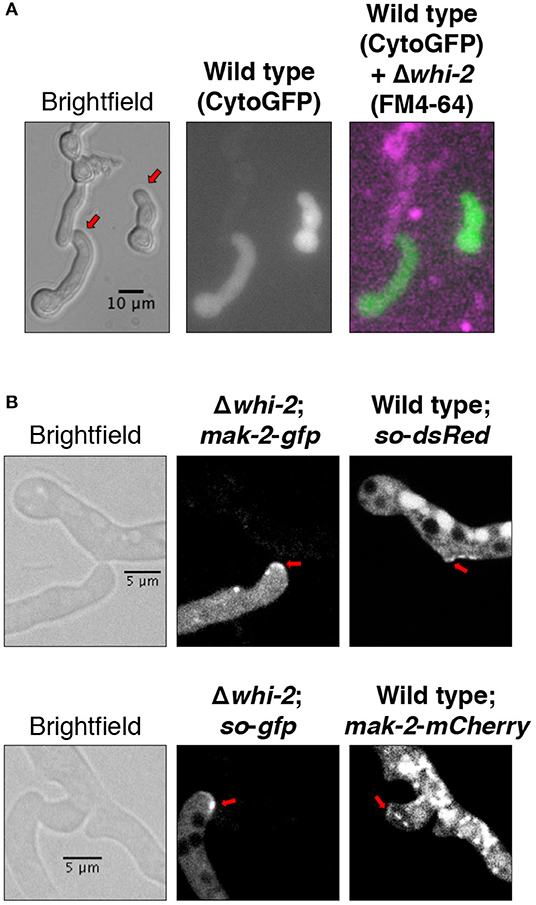
Figure 2. Cells lacking whi-2 are capable of establishing chemotropic interactions with the wild type cells and show recruitment of MAK-2 and SO to CATs. (A) Cytoplasmic GFP-expressing wild type cells were mixed with FM4-64-stained Δwhi-2 cells and observed by microscopy after germination. (B) MAK-2-GFP-expressing (upper panel) or SO-GFP-expressing (lower panel) Δwhi-2 cells were mixed, respectively, with SO-dsRed-expressing or MAK-2-mCherry-expressing wild type cells and observed by microscopy after germination. Note the formation of CATs (red arrows).
Phosphorylation of the MAP kinases MAK-1 and MAK-2 is reduced in Δwhi-2 germlings (Fu et al., 2014). To further explore the relationship between the MAK-2 pathway and WHI-2, we utilized a gain-of-function mutation nrc-1 allele (proline to serine mutation at position 451 of NRC-1). Strains containing this allele show a ~12-fold increase in MAK-2 phosphorylation (Dettmann et al., 2012). When the nrc-1P451S allele was introduced into the Δwhi-2 mutant, the formation of proconidial chains was suppressed and cell fusion-associated chemotropism was restored to levels similar to an nrc-1P451S strain (Figures 1A–C). At the macroscopic level, aerial hyphae development was also partially restored in the Δwhi-2 mutant expressing nrc-1P451S (Figure S2). These data place WHI-2 upstream of the STE-50/STE-20/HAM-5/NRC-1/MEK-2/MAK-2/PP-1/ADV-1 signal transduction pathway.
Deletion of whi-2 Results in Cells That Accumulate Endocytic Intermediates and Possess a Defective Mitochondrial Network
In S. cerevisiae, the deletion of WHI2 causes a defect in endocytosis (Care et al., 2004). Here we employed the lipophilic styryl dye FM4-64 to track the accumulation of endocytic intermediates in N. crassa germlings. FM4-64 is unable to freely cross membranes; instead it is anchored on the outer leaflet of the plasma membrane bilayer. When endocytic membranous structures are formed, the dye becomes incorporated intracellularly into the endosomes and Golgi, and subsequently into the vacuoles (Fischer-Parton et al., 2000); it has been shown that endocytosis mutants show delayed or fragmented staining of intracellular vesicles (Gachet and Hyams, 2005; Martin et al., 2005). N. crassa wild type germlings readily showed plasma membrane and intracellular staining and round membranous structures were observed in the cytoplasm as early as at ~9.5 min after the addition of FM4-64 (Video 1 and Figure 3A). After 30 min of incubation with FM4-64, wild type cells displayed large FM4-64-positive membranous structures (Figure 3B). In contrast, the intracellular accumulation of FM4-64 in Δwhi-2 cells was delayed and appeared as small elements, typical of fungal mutants with endocytosis defects (Gachet and Hyams, 2005; Martin et al., 2005) (Video 1 and Figures 3A,B).

Figure 3. Strains lacking whi-2 are defective in the accumulation of FM4-64-positive membranous structures and show defects in mitochondrial dynamics. (A,B) The uptake of FM4-64 via plasma membrane-derived structures was followed over time in wild type and Δwhi-2 germlings. A time course experiment is shown in (A). Note the formation of large FM4-64-positive membranous structures in the wild type strain (arrows). Representative images after 30 min of incubation are shown in (B). These panels are complemented by Video 1. (C) The organization of the mitochondrial network was examined using an ARG-4-GFP construct (Bowman et al., 2009). Note the appearance of fragmented (“frag”) and clusters (“cluster”) of mitochondria in Δwhi-2 cells. (D) Mitochondrial dynamics during cell fusion were evaluated by producing time-lapse videos of interacting wild type germlings. Note the absence of mitochondria at the fusion spot during the first minutes after contact (arrow). This panel is complemented by Video 2.
S. cerevisiae whi2 mutants show highly fragmented mitochondria or agglomeration of mitochondria in ball-like structures, depending on the growth phase (Leadsham et al., 2009; Mendl et al., 2011). We used a strain harboring a gfp-tagged arg-4 (encoding the mitochondrial acetylornithine-glutamate acetyltransferase) (Bowman et al., 2009) to image the mitochondrial network in N. crassa germlings. In wild type germlings, mitochondria were abundant and formed a tubular-shaped network (Figure 3C). During cell communication and germling fusion, the shape of the mitochondrial network remained unchanged, although organelles were excluded from the tip of CATs until fusion and cytoplasmic continuity were attained (Figure 3D and Video 2). In contrast to the mitochondrial organization in wild type cells, Δwhi-2 germlings displayed aberrant mitochondria that appeared to be fragmented and clustered into “masses” (Figure 3C). These observations indicated that WHI-2 was required for normal endocytosis and mitochondrial morphology in N. crassa.
CSP-6 and AMPH-1 Are Two Fusion Proteins Whose Function Is Related to WHI-2
In a previous deletion strain screening, csp-6 and amph-1 were also found to be required for cell fusion in N. crassa (Fu et al., 2011, 2014; Chinnici et al., 2014). In addition, Δcsp-6 and Δamph-1 cells also have a defect in conidial separation (Figure 4A and Figure S2), phenotypically similar to proconidial chains in Δwhi-2 mutants (Figure 1C) (Fu et al., 2014; Ghosh et al., 2014). CSP-6 (NCU08380) is a Haloacid Dehalogenase (HAD) family Ser/Thr phosphatase and the ortholog of Psr1/2 from S. cerevisiae. In yeast and in N. crassa, WHI-2 and CSP-6 have been shown to directly interact (Kaida et al., 2002; Zhou et al., 2018). AMPH-1 (NCU01069) harbors an Arfaptin Homology/Bin-Amphiphysin-Rvs (AH/BAR) domain; AMPH-1 homologs in S. cerevisiae (Rvs161p and Rvs167p) regulate the actin cytoskeleton, endocytosis and mating, while in neurons AMPH-1 plays a role in clathrin-mediated endocytosis (Takei et al., 1999; Friesen et al., 2006). Although phenotypically similar, the growth of the Δamph-1 mutant was more affected than growth of the Δwhi-2 or Δcsp-6 mutants (Figure S2).
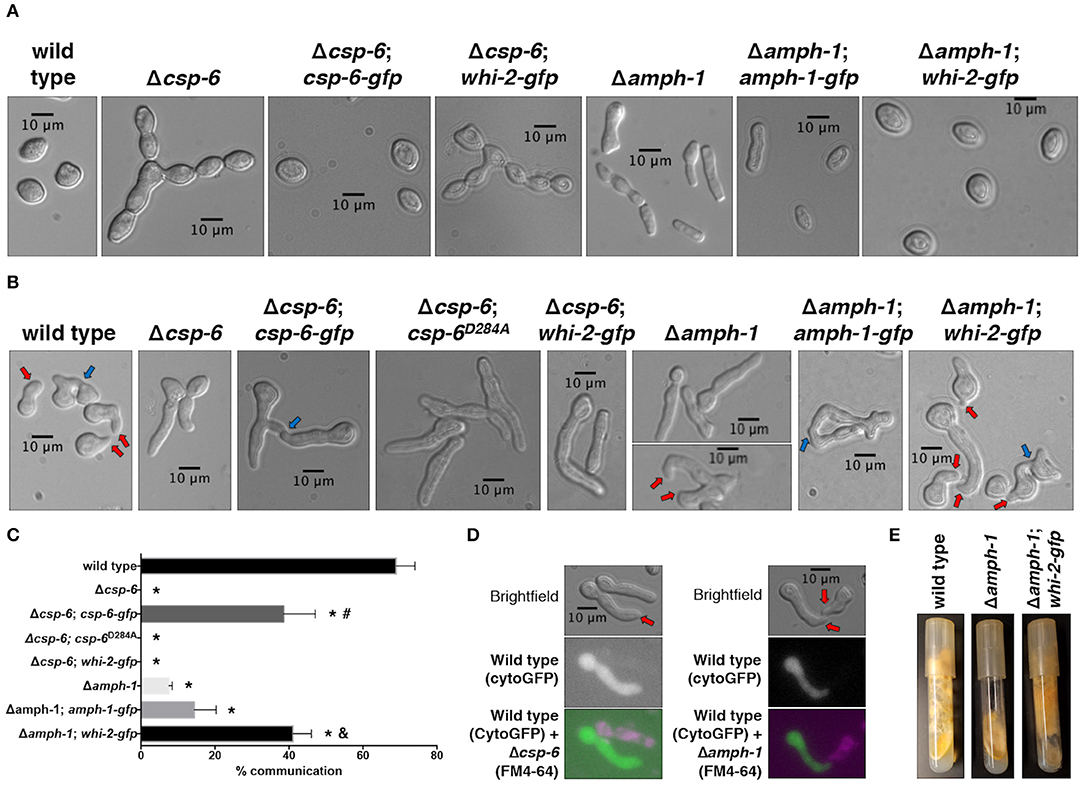
Figure 4. The mis-expression of whi-2 in cells lacking amph-1 but not csp-6 compensates for germling communication and asexual defects. (A) Conidial morphology of indicated strains was analyzed by brightfield microscopy. (B) Cell-cell communication between cells from single strains (genetically identical) were evaluated between closely positioned germlings (B, 4 h post-inoculation). In (B), red and blue arrows indicate conidial anastomosis tubes (CATs) and communication/fusion events, respectively. Note the presence of CATs in the Δamph-1 strain (lower micrograph, red arrows). The relative frequency of cell-cell communication of strains shown in (B) is shown in (C). Error bars represent the standard deviation.*p-value < 0.0001 vs. wild type; #p-value < 0.0001 vs. Δcsp-6; and, p-value < 0.0001 vs. Δamph-1. (D) Cytoplasmic GFP-expressing wild type cells were mixed with FM4-64-stained Δcsp-6 or Δamph-1 cells and observed by microscopy after germination. Note the formation of CATs (red arrows). (E) The production of aerial hyphae and sporulation was evaluated after 7 days of growth for the indicated strains.
We confirmed that cell communication in populations of Δcsp-6 germlings was absent, while a low frequency of CAT formation was observed in self-pairings of Δamph-1 germlings (Figures 4B,C). The introduction of a gfp-tagged csp-6 or amph-1 allele partially compensated for the developmental defects observed in Δcsp-6 and Δamph-1 mutants, respectively (Figures 4A–C and Figure S2). Since the complementation of amph-1 with the gfp-tagged amph-1 allele was modest (Figure 4C), we conducted a co-segregation analysis by crossing the wild type and Δamph-1 deletion strains and analyzing the resistance of the progeny to hygromycin B; all strains that displayed wild type-like growth were sensitive to hygromycin B whereas all strains with flat-like growth were resistant to it (Figure S3). The Δcsp-6 and Δamph-1 mutants communicated with wild type germlings, although at low frequency (Figure 4D); this observation had been previously suggested for Δamph-1 (Fu et al., 2014).
To explore how the predicted phosphatase activity of CSP-6 plays a role during cell fusion and asexual development in N. crassa, we constructed a mutated allele of CSP-6 predicted to abolish activity. In S. cerevisiae, a point mutation in the predicted catalytic aspartic acid residue of the hhhDxDx(T/V) motif—where “h” is a hydrophobic residue and “x” is any residue—abolished phosphatase activity (Rebay, 2015). This aspartic acid residue belongs to the HAD domain and is conserved in N. crassa. A Δcsp-6 mutant that carried a phosphatase-dead, Pccg-1-driven csp-6D284A allele showed the same defects as the Δcsp-6 deletion strain, namely the lack of aerial hyphae (Figure S2) and inability to form CATs and undergo cell fusion (Figures 4B,C).
To assess the relationship between whi-2, amph-1, and csp-6, we placed an extra copy of whi-2 under the control of a constitutive promoter (Pccg-1) in the Δamph-1 and Δcsp-6 strains. The Δamph-1;Pccg-1-whi-2 strain showed suppression of the conidial separation defect of the Δamph-1 mutant (Figure 4A), restoration of CAT formation and chemotropic interactions (Figures 4B,C), and normal aerial hyphae development (Figure 4E). In contrast, over-expression of whi-2 did not complement the morphological (Figure S2) or communication defects of the Δcsp-6 mutant (Figures 4A–C). These data showed that the deletion of whi-2, csp-6, and amph-1 leads to similar cellular phenotypes and that the mis-expression of whi-2 compensated for the absence of amph-1, indicating that AMPH-1 functions upstream of WHI-2.
In order to examine if the deletion of amph-1 results in an endocytic defect, as predicted from S. cerevisiae literature (Munn et al., 1995; Kaksonen et al., 2005), and whether the mis-expression of whi-2 also compensated for an endocytosis defect, we employed a flow cytometry-based methodology. The combined labeling of endocytic intermediates and vacuolar lumens (with FM4-64 and carboxydichlorofluorescein diacetate (CDCFDA), respectively) has been previously used to isolate S. cerevisiae mutants unable to properly execute endocytosis and transport plasma membrane material to the vacuole (Zheng et al., 1998). Such mutants display a shift in FM4-64/CDCFDA fluorescence as compared to wild type cells. We adopted a similar strategy, but utilized CMAC Blue to stain the lumen of the vacuoles instead of CDCFDA. The deletion of whi-2 or amph-1 resulted in a shift in FM4-64/CMAC Blue fluorescence profile as compared to wild type cells (Figure 5). However, the Δwhi-2 and Δamph-1 strains carrying Pccg-1-whi-2-gfp showed a profile more similar to wild type cells (Figure 5). These data suggest that endocytosis defect was at least partially compensated for by the mis-expression of whi-2-gfp in Δamph-1 cells. Additionally, the FM4-64/CMAC Blue fluorescence shift in Δwhi-2 cells was also partially recovered by the mis-expression of nrc-1P451S (Figure 5), consistent with suppression of cell fusion defects in Δwhi-2 cells (Figure 1).
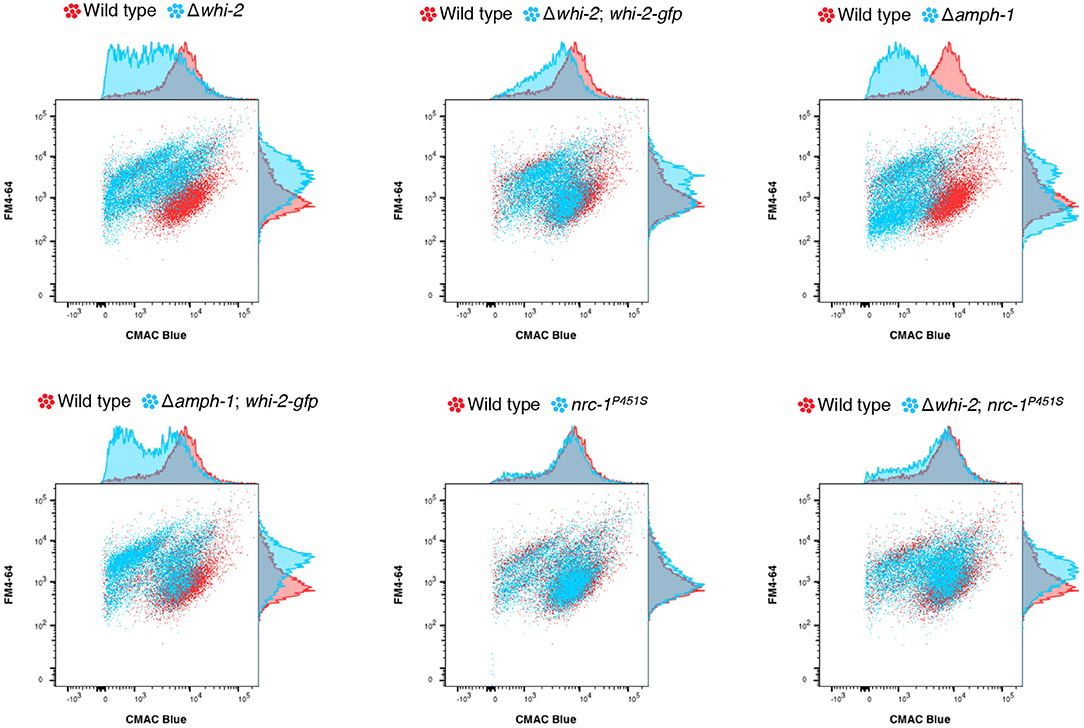
Figure 5. The mis-expression of nrc-1P451S and whi-2-gfp compensates for the endocytosis defect of Δwhi-2 and Δamph-1 cells, respectively. The wild type (red) and other indicated strains (blue) were stained with FM4-64 and CMAC Blue to label endocytic intermediates and vacuolar lumens, respectively. Note that while the wild type shows an overlap of FM4-64 and CMAC Blue, indicating that plasma membrane-derived endocytic intermediates are guided to the vacuoles, cells lacking whi-2 and amph-1 displayed a shift in red/blue fluorescence. n=4. Shown is a representative set of experiments.
WHI-2 and CSP-6 Localize Mainly to the Cell Periphery While AMPH-1 Is Present in Cortical Patches and Puncta
Previous reports showed that WHI-2 localized by immunofluorescence to the cytoplasm, small vesicles or vacuoles and by subcellular fractionation to the cytoplasmic and nuclear fractions; AMPH-1 immunolocalization studies showed a punctate pattern suggestive of small vesicles and by subcellular fractionation CSP-6 was present in the cytoplasmic and nuclear fractions (Fu et al., 2014; Zhou et al., 2018). However, the AMPH-1-RFP construct only marginally complemented the fusion defect of Δamph-1 (Fu et al., 2014), the immunofluorescence for WHI-2 was not very resolved (Fu et al., 2014) and the subcellular fractionation for WHI-2 and CSP-6 showed that both proteins were in all fractions tested (Zhou et al., 2018). Therefore, we sought to determine the localization of these proteins using confocal microscopy by tagging them with GFP, under the control of the constitutive Pccg-1 promoter, and introducing them into their respective deletion strains (Figures 1, 4). In dormant conidia, WHI-2-GFP was distributed across the cytoplasm while CSP-6-GFP was predominantly present in the cell periphery (Figure 6A). AMPH-1 was mainly localized in cortical patches although some cytoplasmic staining was observed (Figure 6A). In germlings, WHI-2 and CSP-6 were present mainly in the cell periphery and occasionally in intracellular puncta (Figure 6A); AMPH-1 was present in cytoplasmic puncta. In mature hyphae, the localization of WHI-2 and CSP-6 was mainly at the cell periphery and at septa (Figure 6A). For WHI-2, we also observed cytoplasmic localization and in the membrane of round intracellular vesicles (Figure 6A). In hyphae, AMPH-1 was present in puncta or small intracellular vesicles that tended to accumulate at the hyphal tip (Figure 6C).
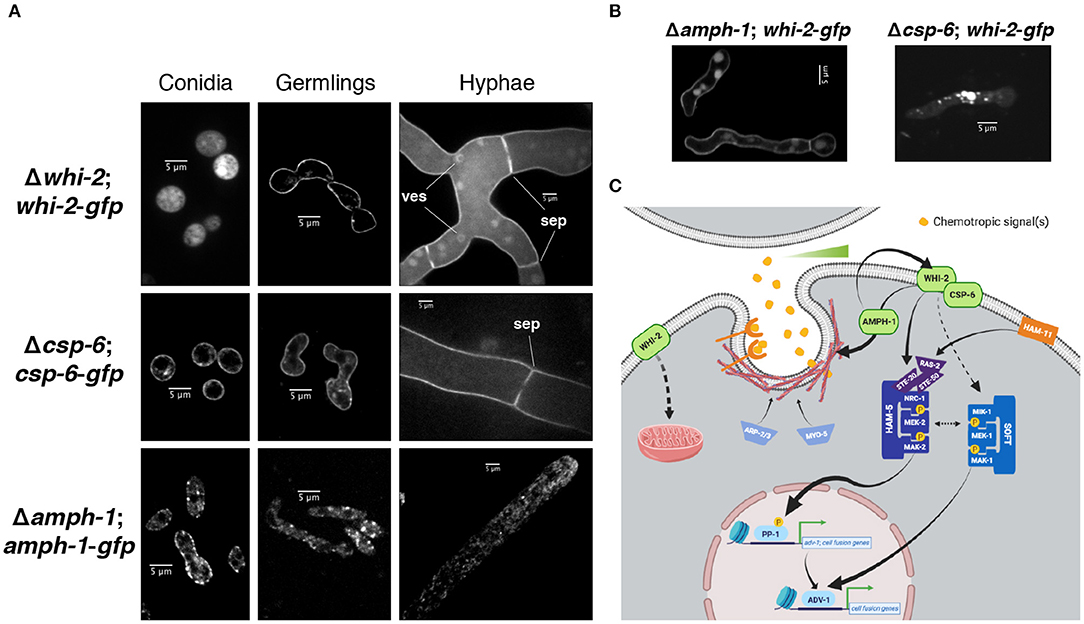
Figure 6. Localization of WHI-2, CSP-6 and AMPH-1 at different asexual stages. (A) The localization of WHI-2 (upper panel), CSP-6 (middle) and AMPH-1 (lower) was examined in the deletion strains carrying a GFP-tagged construct of the respective protein. Panels from left to right indicate conidia, germlings and mature hyphae. ves, intracellular vesicle membrane; sep, septum. (B) Localization of WHI-2 in Δamph-1 and Δcsp-6 mutant background. (C) Schematic model of the role of WHI-2, CSP-6, and AMPH-1 in signal reception during cell-cell communication of N. crassa germlings. Sensing of a gradient of the chemotropic signal(s) may occur via endocytosis of the chemotropic signal or via internalization of a receptor. WHI-2 and CSP-6 interact and sit at the cell periphery. AMPH-1 could be involved in vesicle scission during endocytosis and is epistatic to WHI-2. WHI-2 may also play a role in one or more steps of endocytosis, potentially via AMPH-1. WHI-2 functions upstream of the NRC-1/MEK-2/MAK-2 signaling pathway. Previous western blot data (Fu et al., 2014) indicated that WHI-2 could also affect signaling through another MAPK pathway composed of a MIK-1/MEK-1/MAK-1 module. These two MAPK cascades are involved in the activation of the transcription factors PP-1 and ADV-1, which directly regulate the expression of genes involved in cell communication and fusion. WHI-2 also controls the organization of the intracellular mitochondrial network. For simplicity, a number of signaling components and phosphorylation events have been left out (Please see Fischer and Glass, 2019, for an extended review).
A Δamph-1;whi-2-gfp strain was able to communicate and fuse (Figures 4B,C), but a Δcsp-6;whi-2-gfp strain displayed the same morphological defects as its parental deletion mutant (Figures 4B,C). We therefore assessed whether localization of WHI-2-GFP was affected in the Δamph-1 mutant relative to the Δcsp-6 mutant and wild type cells. In the Δamph-1 mutant, WHI-2 displayed a cell periphery localization, consistent with what was observed in a Δwhi-2;whi-2-gfp strain (Figure 6A). However, in the Δcsp-6 mutant, WHI-2-GFP was no longer present in the cell periphery, but accumulated in cytoplasmic patches and puncta (Figure 6B). These data suggest that CSP-6 is required for localization of WHI-2, consistent with reports of physical interaction of these two proteins in N. crassa and S. cerevisiae (Kaida et al., 2002; Zhou et al., 2018).
Discussion
Germinated asexual spores of N. crassa cooperate during the formation of the somatic mycelial network by undergoing regulated cell fusion. Our study highlights the importance of WHI-2, an ortholog of the S. cerevisiae Whi2p, which binds to Psr1p (HAD family of protein phosphatases; CSP-6 ortholog) and regulates the response to nutritional stress by affecting both TORC1 and the Ras-cAMP-PKA pathway (Sudbery et al., 1980; Kaida et al., 2002; Müller and Reichert, 2011; Chen et al., 2018). The deletion of whi-2 in N. crassa resulted in a panoply of morphological phenotypes, including defects in conidial separation, an inability to undergo chemotropic interactions and cell fusion, aerial hyphae formation, growth rate and female fertility. Our data showed that the lack of whi-2 also caused an accumulation of FM4-64-positive membranous structures that likely correspond to endocytic intermediates and the destabilization of the mitochondrial organization. Using a constitutive nrc-1 allele, we showed that WHI-2 plays an active role during cell-cell communication in N. crassa by transducing signals to the NRC-1/MEK-2/MAK-2 pathway. In support of this conclusion, the deletion of whi-2, csp-6 or amph-1 abolishes activation of prm-1 (Fischer et al., 2019), which encodes a protein involved in plasma membrane merger during cell fusion (Fleissner et al., 2009a). The prm-1 gene is directly regulated by the transcription factors PP-1 and ADV-1, which sit downstream of the NRC-1/MEK-2/MAK-2 pathway (Fischer et al., 2018). These data suggest that WHI-2 functions upstream in the sequence of signal transduction events that lead to chemotropism and cell fusion (Figure 6C). In yeast, Whi2p is a nutrient sensor (Müller and Reichert, 2011; Chen et al., 2018), and in mammalian models a specific form of endocytosis is regulated by nutrient availability (Pang et al., 2014). In N. crassa, the Δwhi-2 mutant was phenotypically similar when grown under various carbon and nitrogen sources, indicating further biochemical and genetic analyses are required to assess whether WHI-2 affects nutrient sensing. It will be also interesting to examine the range of phenotypes controlled by the WHI-2/MAPK axis by taking the advantage of the Δwhi-2; nrc-1P451S strain.
Although endocytosis is required for proper morphogenesis and apical growth in filamentous fungi (Riquelme et al., 2018), it has been suggested that these organisms might not undergo clathrin-dependent endocytosis (Schultzhaus et al., 2017). An alternative form of endocytosis termed fluid-phase endocytosis or pinocytosis, cells internalize small molecules present in the extracellular fluid through invaginations of the plasma membrane, forming intracellular vesicles (Epp et al., 2013) and a role for Whi2p during pinocytosis has been suggested as a way to sense cell density in yeast (Care et al., 2004). Additionally, this form of endocytosis has been proposed to mediate cell-cell communication between animal alveolar macrophages (Schneider et al., 2017). Pinocytosis could function as a mechanism to survey the extracellular space for gradients of certain peptides or metabolites and whenever the concentration is above a defined threshold, as when cells that produce a communication signal are in close proximity, the activation of the cell-cell fusion machinery would ensue.
Evidence from various organisms support the hypothesis that endocytosis could play a role in cell-cell communication in N. crassa (Figure 6C). In animals, fusogens are internalized via RAB-5 and DYNAMIN-1 GTPases-dependent clathrin-mediated endocytosis (Shin et al., 2014; Smurova and Podbilewicz, 2016). In the filamentous fungus Ustilago maydis, the pheromone receptor Pra1 cycles between the early endosomes and the plasma membrane in endocytic vesicles, triggering cell communication-associated MAPK signaling pathways when the mating pheromone is perceived. Mutations that impair endocytosis cause Pra1 to be depleted from the plasma membrane and abolish cell fusion (Fuchs et al., 2006). During mating in S. cerevisiae, the pheromone receptors and pheromone transporter are subject to regulation by endocytosis (Davis et al., 1993; Berkower et al., 1994).
Our observations that AMPH-1 is epistatic to WHI-2 adds strength to the hypothesis that endocytosis could be important for cell-cell communication in N. crassa. The yeast ortholog of AMPH-1, Rvs161p (previously termed Fus7p and End6p) is involved in endocytosis (Munn et al., 1995; Kaksonen et al., 2005) and forms a heterodimer with another BAR domain-containing protein, Rvs167p, to bind to phospholipid membranes (Friesen et al., 2006). BAR domains modulate membrane curvature by promoting vesicle scission at the neck of plasma membrane invaginations during endocytosis (Takei et al., 1999; McMahon and Gallop, 2005). Rvs161p also plays a role in yeast mating cell fusion by binding and shuttling Fus2p from the nucleus to the shmoo tip and by recruiting Cdc42p to the cell-cell contact spot (Smith et al., 2017); orthologs of Fus2p are absent in N. crassa. Despite the role of Rvs161p during mating cell fusion, some mutations in RVS161 cause a defect in endocytosis but not in mating (Brizzio et al., 1998). Further investigations are required to ascertain if the situation in N. crassa is analogous. In addition to AMPH-1, mutations in other components of the endocytosis machinery in N. crassa, namely MYO-5 (Dettmann et al., 2014; Ramírez-Del Villar et al., 2019) and the ARP-2/ARP-3 complex proteins (Roca et al., 2010), also result in cell-cell communication defects (Figure 6C).
A strain containing a deletion of csp-6 phenocopied a Δwhi-2 mutant. WHI-2 physically interacts with CSP-6 to dephosphorylate the circadian clock regulator WC-1, which in turn results in loss of activation of ADV-1 (Zhou et al., 2018); ADV-1 directly regulates cell communication and fusion genes in N. crassa (Fischer et al., 2018). Our localization studies showed that WHI-2 and CSP-6 share the same subcellular distribution at the cell periphery. This observation supports our results that put WHI-2 and CSP-6 upstream of a signaling cascade that activates ADV-1. Importantly, a CSP-6 allele that lacks phosphatase activity was unable to complement the defects of a strain lacking csp-6, indicating that the phosphatase function of this protein is essential for CSP-6 function.
Although a receptor-mediated system where an extracellular receptor binds a chemotropic ligand has been suggested to underlie cell-cell communication in N. crassa, we speculate that endocytosis could be an important mechanism of signal sensing (Figure 6C). The peripheral localization of WHI-2, in line with previous results in S. cerevisiae (Huh et al., 2003), fits our proposed role in endocytosis, since this is where the endocytic machinery is recruited to initiate vesicle budding. WHI-2 and CSP-6 may be regulators of this machinery, act as sensors, or both; AMPH-1 is potentially involved in endocytosis vesicle scission (McMahon and Gallop, 2005). The fact that a Δwhi-2 mutant is capable of a low level of communication with the wild type strain, including the recruitment of MAK-2 and SO to the fusion tips, suggests that the absence of whi-2 does not hinder the production of pro-fusion cues, attracting the wild type partner, but may obstruct proper sensing. Signal receiving may be desensitized in the Δwhi-2, Δcsp-6, and Δamph-1 mutants and perhaps can only occur when the levels of chemotropic ligand(s) that are much higher than the levels required for communication in wild type cells. Only a single additional mutant previously identified in N. crassa, ham-11, shows a defect in self-fusion, but can undergo robust chemotropic interactions and fusion with wild type cells (Leeder et al., 2013; Fischer et al., 2019). Recent evidence suggests that HAM-11 also functions upstream of the NRC-1/MEK-2/MAK-2 signal transduction pathway (Fischer et al., 2019). HAM-11 is predicted to encode a hypothetical plasma membrane protein enriched in plasma membrane fractions. These data and data presented here suggest the AMPH-1, CSP-6-WHI-2, and HAM-11 function upstream of the MAK-2 signaling complex and play a role in sensitizing cells for signal reception during chemotropic interactions.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Author Contributions
AG and NG designed the study and wrote the article. AG performed all experiments. KC contributed to the quantification of cell communication. SC-S contributed to the flow cytometry assay and mutant segregation analysis.
Funding
This work was supported in part by a National Science Foundation Grant (MCB1412411) and a Laboratory Directed Research and Development Program of Lawrence Berkeley National Laboratory under U.S. Department of Energy Contract No. DE-AC02-05CH11231 to NLG.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Wilfried Jonkers for early comments and suggestions on this study, Jens Heller for critical reading of this manuscript, Anne Dettmann and Stephan Seiler for kindly providing the NRC-1P451S strain and Barry Bowman for kindly providing the ARG-4-GFP strain.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.03162/full#supplementary-material
References
Berkower, C., Loayza, D., and Michaelis, S. (1994). Metabolic instability and constitutive endocytosis of STE6, the a-factor transporter of Saccharomyces cerevisiae. Mol. Biol. Cell 5, 1185–1198. doi: 10.1091/mbc.5.11.1185
Binley, K. M., Radcliffe, P. A., Trevethick, J., Duffy, K. A., and Sudbery, P. E. (1999). The yeast PRS3 gene is required for cell integrity, cell cycle arrest upon nutrient deprivation, ion homeostasis and the proper organization of the actin cytoskeleton. Yeast 15, 1459–1469.
Borkovich, K. A., Alex, L. A., Yarden, O., Freitag, M., Turner, G. E., Read, N. D., et al. (2004). Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68, 1–108. doi: 10.1128/MMBR.68.1.1-108.2004
Bowman, B. J., Draskovic, M., Freitag, M., and Bowman, E. J. (2009). Structure and distribution of organelles and cellular location of calcium transporters in Neurospora crassa. Eukaryotic Cell 8, 1845–1855. doi: 10.1128/EC.00174-09
Brizzio, V., Gammie, A. E., and Rose, M. D. (1998). Rvs161p interacts with Fus2p to promote cell fusion in Saccharomyces cerevisiae. J. Cell Biol. 141, 567–584. doi: 10.1083/jcb.141.3.567
Care, A., Vousden, K. A., Binley, K. M., Radcliffe, P., Trevethick, J., Mannazzu, I., et al. (2004). A synthetic lethal screen identifies a role for the cortical actin patch/endocytosis complex in the response to nutrient deprivation in Saccharomyces cerevisiae. Genetics 166, 707–719. doi: 10.1534/genetics.166.2.707
Chen, X., Wang, G., Zhang, Y., Dayhoff-Brannigan, M., Diny, N. L., Zhao, M., et al. (2018). Whi2 is a conserved negative regulator of TORC1 in response to low amino acids. PLoS Genet. 14:e1007592. doi: 10.1371/journal.pgen.1007592
Cheng, W. C., Teng, X., Park, H. K., Tucker, C. M., Dunham, M. J., and Hardwick, J. M. (2008). Fis1 deficiency selects for compensatory mutations responsible for cell death and growth control defects. Cell Death Differ. 15, 1838–1846. doi: 10.1038/cdd.2008.117
Chinnici, J. L., Fu, C., Caccamise, L. M., Arnold, J. W., and Free, S. J. (2014). Neurospora crassa female development requires the PACC and other signal transduction pathways, transcription factors, chromatin remodeling, cell-to-cell fusion, and autophagy. PLoS ONE 9:e110603. doi: 10.1371/journal.pone.0110603
Colot, H. V., Park, G., Turner, G. E., Ringelberg, C., Crew, C. M., Litvinkova, L., et al. (2006). A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. U.S.A. 103, 10352–10357. doi: 10.1073/pnas.0601456103
Comyn, S. A., Flibotte, S., and Mayor, T. (2017). Recurrent background mutations in WHI2 impair proteostasis and degradation of misfolded cytosolic proteins in Saccharomyces cerevisiae. Sci. Rep. 7:4183. doi: 10.1038/s41598-017-04525-8
Davis, N. G., Horecka, J. L., and Sprague, G. F. Jr. (1993). Cis- and trans-acting functions required for endocytosis of the yeast pheromone receptors. J. Cell Biol. 122, 53–65. doi: 10.1083/jcb.122.1.53
Dettmann, A., Heilig, Y., Valerius, O., Ludwig, S., and Seiler, S. (2014). Fungal communication requires the MAK-2 pathway elements STE-20 and RAS-2, the NRC-1 adapter STE-50 and the MAP kinase scaffold HAM-5. PLoS Genet. 10:e1004762. doi: 10.1371/journal.pgen.1004762
Dettmann, A., Illgen, J., März, S., Schürg, T., Fleissner, A., and Seiler, S. (2012). The NDR kinase scaffold HYM1/MO25 is essential for MAK2 map kinase signaling in Neurospora crassa. PLoS Genet. 8:e1002950. doi: 10.1371/journal.pgen.1002950
Epp, E., Nazarova, E., Regan, H., Douglas, L. M., Konopka, J. B., Vogel, J., et al. (2013). Clathrin- and Arp2/3-independent endocytosis in the fungal pathogen Candida albicans. MBio 4:e00476–13. doi: 10.1128/mBio.00476-13
Fischer, M. S., and Glass, N. L. (2019). Communicate and fuse: how filamentous fungi establish and maintain an interconnected mycelial network. Front. Microbiol. 10:619. doi: 10.3389/fmicb.2019.00619
Fischer, M. S., Jonkers, W., and Glass, N. L. (2019). Integration of self and non-self recognition modulates asexual cell-to-cell communication in Neurospora crassa. Genetics 211, 1255–1267. doi: 10.1534/genetics.118.301780
Fischer, M. S., Wu, V. W., Lee, J. E., O'Malley, R. C., and Glass, N. L. (2018). Regulation of cell-to-cell communication and cell wall integrity by a network of map kinase pathways and transcription factors in Neurospora crassa. Genetics 209, 489–506. doi: 10.1534/genetics.118.300904
Fischer-Parton, S., Parton, R. M., Hickey, P. C., Dijksterhuis, J., Atkinson, H. A., and Read, N. D. (2000). Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J. Microsc. 198, 246–259. doi: 10.1046/j.1365-2818.2000.00708.x
Fleissner, A., Diamond, S., and Glass, N. L. (2009a). The Saccharomyces cerevisiae PRM1 homolog in Neurospora crassa is involved in vegetative and sexual cell fusion events but also has postfertilization functions. Genetics 181, 497–510. doi: 10.1534/genetics.108.096149
Fleissner, A., Leeder, A. C., Roca, M. G., Read, N. D., and Glass, N. L. (2009b). Oscillatory recruitment of signaling proteins to cell tips promotes coordinated behavior during cell fusion. Proc. Natl. Acad. Sci. U.S.A. 106, 19387–19392. doi: 10.1073/pnas.0907039106
Freitag, M., Hickey, P. C., Raju, N. B., Selker, E. U., and Read, N. D. (2004). GFP as a tool to analyze the organization, dynamics and function of nuclei and microtubules in Neurospora crassa. Fungal Genet. Biol. 41, 897–910. doi: 10.1016/j.fgb.2004.06.008
Friesen, H., Humphries, C., Ho, Y., Schub, O., Colwill, K., and Andrews, B. (2006). Characterization of the yeast amphiphysins Rvs161p and Rvs167p reveals roles for the Rvs heterodimer in vivo. Mol. Biol. Cell 17, 1306–1321. doi: 10.1091/mbc.e05-06-0476
Fu, C., Ao, J., Dettmann, A., Seiler, S., and Free, S. J. (2014). Characterization of the Neurospora crassa cell fusion proteins, HAM-6, HAM-7, HAM-8, HAM-9, HAM-10, AMPH-1 and WHI-2. PLoS ONE 9:e107773. doi: 10.1371/journal.pone.0107773
Fu, C., Iyer, P., Herkal, A., Abdullah, J., Stout, A., and Free, S. J. (2011). Identification and characterization of genes required for cell-to-cell fusion in Neurospora crassa. Eukaryotic Cell 10, 1100–1109. doi: 10.1128/EC.05003-11
Fuchs, U., Hause, G., Schuchardt, I., and Steinberg, G. (2006). Endocytosis is essential for pathogenic development in the corn smut fungus Ustilago maydis. Plant Cell 18, 2066–2081. doi: 10.1105/tpc.105.039388
Gabriela Roca, M., Read, N. D., and Wheals, A. E. (2005). Conidial anastomosis tubes in filamentous fungi. FEMS Microbiol. Lett. 249, 191–198. doi: 10.1016/j.femsle.2005.06.048
Gachet, Y., and Hyams, J. S. (2005). Endocytosis in fission yeast is spatially associated with the actin cytoskeleton during polarised cell growth and cytokinesis. J. Cell Sci. 118 (Pt 18), 4231–4242. doi: 10.1242/jcs.02530
Galagan, J. E., Calvo, S. E., Borkovich, K. A., Selker, E. U., Read, N. D., Jaffe, D., et al. (2003). The genome sequence of the filamentous fungus Neurospora crassa. Nature 422, 859–868. doi: 10.1038/nature01554
Ghosh, A., Servin, J. A., Park, G., and Borkovich, K. A. (2014). Global analysis of serine/threonine and tyrosine protein phosphatase catalytic subunit genes in Neurospora crassa reveals interplay between phosphatases and the p38 mitogen-activated protein kinase. G3 4, 349–365. doi: 10.1534/g3.113.008813
Glass, N. L., Rasmussen, C., Roca, M. G., and Read, N. D. (2004). Hyphal homing, fusion and mycelial interconnectedness. Trends Microbiol. 12, 135–141. doi: 10.1016/j.tim.2004.01.007
Gonçalves, A. P., Heller, J., Span, E. A., Rosenfield, G., Do, H. P., Palma-Guerrero, J., et al. (2019). Allorecognition upon fungal cell-cell contact determines social cooperation and impacts the acquisition of multicellularity. Curr. Biol. 29, 3006–3017. doi: 10.1016/j.cub.2019.07.060
Gresham, D., Desai, M. M., Tucker, C. M., Jenq, H. T., Pai, D. A., Ward, A., et al. (2008). The repertoire and dynamics of evolutionary adaptations to controlled nutrient-limited environments in yeast. PLoS Genet. 4:e1000303. doi: 10.1371/journal.pgen.1000303
Hernández, J. M., and Podbilewicz, B. (2017). The hallmarks of cell-cell fusion. Development 144, 4481–4495. doi: 10.1242/dev.155523
Huh, W. K., Falvo, J. V., Gerke, L. C., Carroll, A. S., Howson, R. W., Weissman, J. S., et al. (2003). Global analysis of protein localization in budding yeast. Nature 425, 686–691. doi: 10.1038/nature02026
Jones, P., Binns, D., Chang, H. Y., Fraser, M., Li, W., McAnulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Jonkers, W., Leeder, A. C., Ansong, C., Wang, Y., Yang, F., Starr, T. L., et al. (2014). HAM-5 functions as a MAP kinase scaffold during cell fusion in Neurospora crassa. PLoS Genet. 10:e1004783. doi: 10.1371/journal.pgen.1004783
Kaida, D., Yashiroda, H., Toh-e, A., and Kikuchi, Y. (2002). Yeast Whi2 and Psr1-phosphatase form a complex and regulate STRE-mediated gene expression. Genes Cells 7, 543–552. doi: 10.1046/j.1365-2443.2002.00538.x
Kaksonen, M., Toret, C. P., and Drubin, D. G. (2005). A modular design for the clathrin- and actin-mediated endocytosis machinery. Cell 123, 305–320. doi: 10.1016/j.cell.2005.09.024
Kvitek, D. J., and Sherlock, G. (2013). Whole genome, whole population sequencing reveals that loss of signaling networks is the major adaptive strategy in a constant environment. PLoS Genet. 9:e1003972. doi: 10.1371/journal.pgen.1003972
Lang, G. I., Rice, D. P., Hickman, M. J., Sodergren, E., Weinstock, G. M., Botstein, D., et al. (2013). Pervasive genetic hitchhiking and clonal interference in forty evolving yeast populations. Nature 500, 571–574. doi: 10.1038/nature12344
Leadsham, J. E., Miller, K., Ayscough, K. R., Colombo, S., Martegani, E., Sudbery, P., et al. (2009). Whi2p links nutritional sensing to actin-dependent Ras-cAMP-PKA regulation and apoptosis in yeast. J. Cell Sci. 122(Pt 5), 706–715. doi: 10.1242/jcs.042424
Leeder, A. C., Jonkers, W., Li, J., and Glass, N. L. (2013). Early colony establishment in Neurospora crassa requires a MAP kinase regulatory network. Genetics 195, 883–898. doi: 10.1534/genetics.113.156984
Leeder, A. C., Palma-Guerrero, J., and Glass, N. L. (2011). The social network: deciphering fungal language. Nat. Rev. Microbiol. 9, 440–451. doi: 10.1038/nrmicro2580
Martin, R., Walther, A., and Wendland, J. (2005). Ras1-induced hyphal development in Candida albicans requires the formin Bni1. Eukaryotic Cell 4, 1712–1724. doi: 10.1128/EC.4.10.1712-1724.2005
McCluskey, K., Wiest, A., and Plamann, M. (2010). The fungal genetics stock center: a repository for 50 years of fungal genetics research. J. Biosci. 35, 119–126. doi: 10.1007/s12038-010-0014-6
McMahon, H. T., and Gallop, J. L. (2005). Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 438, 590–596. doi: 10.1038/nature04396
Mendl, N., Occhipinti, A., Müller, M., Wild, P., Dikic, I., and Reichert, A. S. (2011). Mitophagy in yeast is independent of mitochondrial fission and requires the stress response gene WHI2. J. Cell Sci. 124(Pt 8), 1339–1350. doi: 10.1242/jcs.076406
Müller, M., and Reichert, A. S. (2011). Mitophagy, mitochondrial dynamics and the general stress response in yeast. Biochem. Soc. Trans. 39, 1514–1519. doi: 10.1042/BST0391514
Munn, A. L., Stevenson, B. J., Geli, M. I., and Riezman, H. (1995). end5, end6, and end7: mutations that cause actin delocalization and block the internalization step of endocytosis in Saccharomyces cerevisiae. Mol. Biol. Cell 6, 1721–1742. doi: 10.1091/mbc.6.12.1721
Pang, H. B., Braun, G. B., Friman, T., Aza-Blanc, P., Ruidiaz, M. E., Sugahara, K. N., et al. (2014). An endocytosis pathway initiated through neuropilin-1 and regulated by nutrient availability. Nat. Commun. 5:4904. doi: 10.1038/ncomms5904
Payen, C., Sunshine, A. B., Ong, G. T., Pogachar, J. L., Zhao, W., and Dunham, M. J. (2016). High-throughput identification of adaptive mutations in experimentally evolved yeast populations. PLoS Genet. 12:e1006339. doi: 10.1371/journal.pgen.1006339
Perez-Torrado, R., Yamada, D., and Defossez, P. A. (2006). Born to bind: the BTB protein-protein interaction domain. Bioessays 28, 1194–1202. doi: 10.1002/bies.20500
Radcliffe, P., Trevethick, J., Tyers, M., and Sudbery, P. (1997a). Deregulation of CLN1 and CLN2 in the Saccharomyces cerevisiae whi2 mutant. Yeast 13, 707–715.
Radcliffe, P. A., Binley, K. M., Trevethick, J., Hall, M., and Sudbery, P. E. (1997b). Filamentous growth of the budding yeast Saccharomyces cerevisiae induced by overexpression of the WHi2 gene. Microbiology 143, 1867–1876. doi: 10.1099/00221287-143-6-1867
Ramírez-Del Villar, A., Roberson, R. W., Callejas-Negrete, O. A., and Mouriño-Pérez, R. R. (2019). The actin motor MYO-5 effect in the intracellular organization of Neurospora crassa. Fungal Genet. Biol. 125, 13–27. doi: 10.1016/j.fgb.2018.11.008
Rebay, I. (2015). Multiple functions of the eya phosphotyrosine phosphatase. Mol. Cell. Biol. 36, 668–677. doi: 10.1128/MCB.00976-15
Riquelme, M., Aguirre, J., Bartnicki-García, S., Braus, G. H., Feldbrügge, M., Fleig, U., et al. (2018). Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol. Mol. Biol. Rev. 82:e00068–17. doi: 10.1128/MMBR.00068-17
Roca, M. G., Kuo, H. C., Lichius, A., Freitag, M., and Read, N. D. (2010). Nuclear dynamics, mitosis, and the cytoskeleton during the early stages of colony initiation in Neurospora crassa. Eukaryotic Cell 9, 1171–1183. doi: 10.1128/EC.00329-09
Sadeh, A., Movshovich, N., Volokh, M., Gheber, L., and Aharoni, A. (2011). Fine-tuning of the Msn2/4-mediated yeast stress responses as revealed by systematic deletion of Msn2/4 partners. Mol. Biol. Cell 22, 3127–3138. doi: 10.1091/mbc.e10-12-1007
Saul, D. J., and Sudbery, P. E. (1985). Molecular cloning of WHI2, a gene involved in the regulation of cell proliferation in Saccharomyces cerevisiae. J. Gen. Microbiol. 131, 1797–1806. doi: 10.1099/00221287-131-7-1797
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Schneider, D. J., Speth, J. M., Penke, L. R., Wettlaufer, S. H., Swanson, J. A., and Peters-Golden, M. (2017). Mechanisms and modulation of microvesicle uptake in a model of alveolar cell communication. J. Biol. Chem. 292, 20897–20910. doi: 10.1074/jbc.M117.792416
Schultzhaus, Z., Johnson, T. B., and Shaw, B. D. (2017). Clathrin localization and dynamics in Aspergillus nidulans. Mol. Microbiol. 103, 299–318. doi: 10.1111/mmi.13557
Serrano, A., Illgen, J., Brandt, U., Thieme, N., Letz, A., Lichius, A., et al. (2018). Spatio-temporal MAPK dynamics mediate cell behavior coordination during fungal somatic cell fusion. J. Cell Sci. 131:jcs213462. doi: 10.1242/jcs.213462
Shin, N. Y., Choi, H., Neff, L., Wu, Y., Saito, H., Ferguson, S. M., et al. (2014). Dynamin and endocytosis are required for the fusion of osteoclasts and myoblasts. J. Cell Biol. 207, 73–89. doi: 10.1083/jcb.201401137
Smith, J. A., Hall, A. E., and Rose, M. D. (2017). Membrane curvature directs the localization of Cdc42p to novel foci required for cell-cell fusion. J. Cell Biol. 216, 3971–3980. doi: 10.1083/jcb.201703169
Smurova, K., and Podbilewicz, B. (2016). RAB-5- and DYNAMIN-1-Mediated Endocytosis of EFF-1 Fusogen Controls Cell-Cell Fusion. Cell Rep. 14, 1517–1527. doi: 10.1016/j.celrep.2016.01.027
Sudbery, P. E., Goodey, A. R., and Carter, B. L. (1980). Genes which control cell proliferation in the yeast Saccharomyces cerevisiae. Nature 288, 401–404. doi: 10.1038/288401a0
Szamecz, B., Boross, G., Kalapis, D., Kovács, K., Fekete, G., Farkas, Z., et al. (2014). The genomic landscape of compensatory evolution. PLoS Biol. 12:e1001935. doi: 10.1371/journal.pbio.1001935
Takei, K., Slepnev, V. I., Haucke, V., and De Camilli, P. (1999). Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat. Cell Biol. 1, 33–39. doi: 10.1038/9004
Teng, X., Dayhoff-Brannigan, M., Cheng, W. C., Gilbert, C. E., Sing, C. N., Diny, N. L., et al. (2013). Genome-wide consequences of deleting any single gene. Mol. Cell 52, 485–494. doi: 10.1016/j.molcel.2013.09.026
Teng, X., Yau, E., Sing, C., and Hardwick, J. M. (2018). Whi2 signals low leucine availability to halt yeast growth and cell death. FEMS Yeast Res. 18:foy095. doi: 10.1093/femsyr/foy095
Treusch, S., Albert, F. W., Bloom, J. S., Kotenko, I. E., and Kruglyak, L. (2015). Genetic mapping of MAPK-mediated complex traits across S. cerevisiae. PLoS Genet 11:e1004913. doi: 10.1371/journal.pgen.1004913
Vogel, H. J. (1956). A convenient growth medium for Neurospora (Medium N). Microbial Genet. Bull. 13, 42–43.
Westergaard, M., and Mitchell, H. K. (1947). Neurospora-V - a synthetic medium favoring sexual reproduction. Am. J. Bot. 34, 573–577. doi: 10.1002/j.1537-2197.1947.tb13032.x
Zheng, B., Wu, J. N., Schober, W., Lewis, D. E., and Vida, T. (1998). Isolation of yeast mutants defective for localization of vacuolar vital dyes. Proc. Natl. Acad. Sci. U.S.A. 95, 11721–11726. doi: 10.1073/pnas.95.20.11721
Keywords: cell fusion, WHI-2, CSP-6, AMPH-1, endocytosis, MAPK
Citation: Gonçalves AP, Chow KM, Cea-Sánchez S and Glass NL (2020) WHI-2 Regulates Intercellular Communication via a MAP Kinase Signaling Complex. Front. Microbiol. 10:3162. doi: 10.3389/fmicb.2019.03162
Received: 15 November 2019; Accepted: 31 December 2019;
Published: 22 January 2020.
Edited by:
Monika Schmoll, Austrian Institute of Technology (AIT), AustriaReviewed by:
Jesus Aguirre, National Autonomous University of Mexico, MexicoJinkui Yang, Yunnan University, China
Copyright © 2020 Gonçalves, Chow, Cea-Sánchez and Glass. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: N. Louise Glass, lglass@berkeley.edu
†Present address: A. Pedro Gonçalves, Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan