Identification of a Novel Papillomavirus Associated with Squamous Cell Carcinoma in a Domestic Cat

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
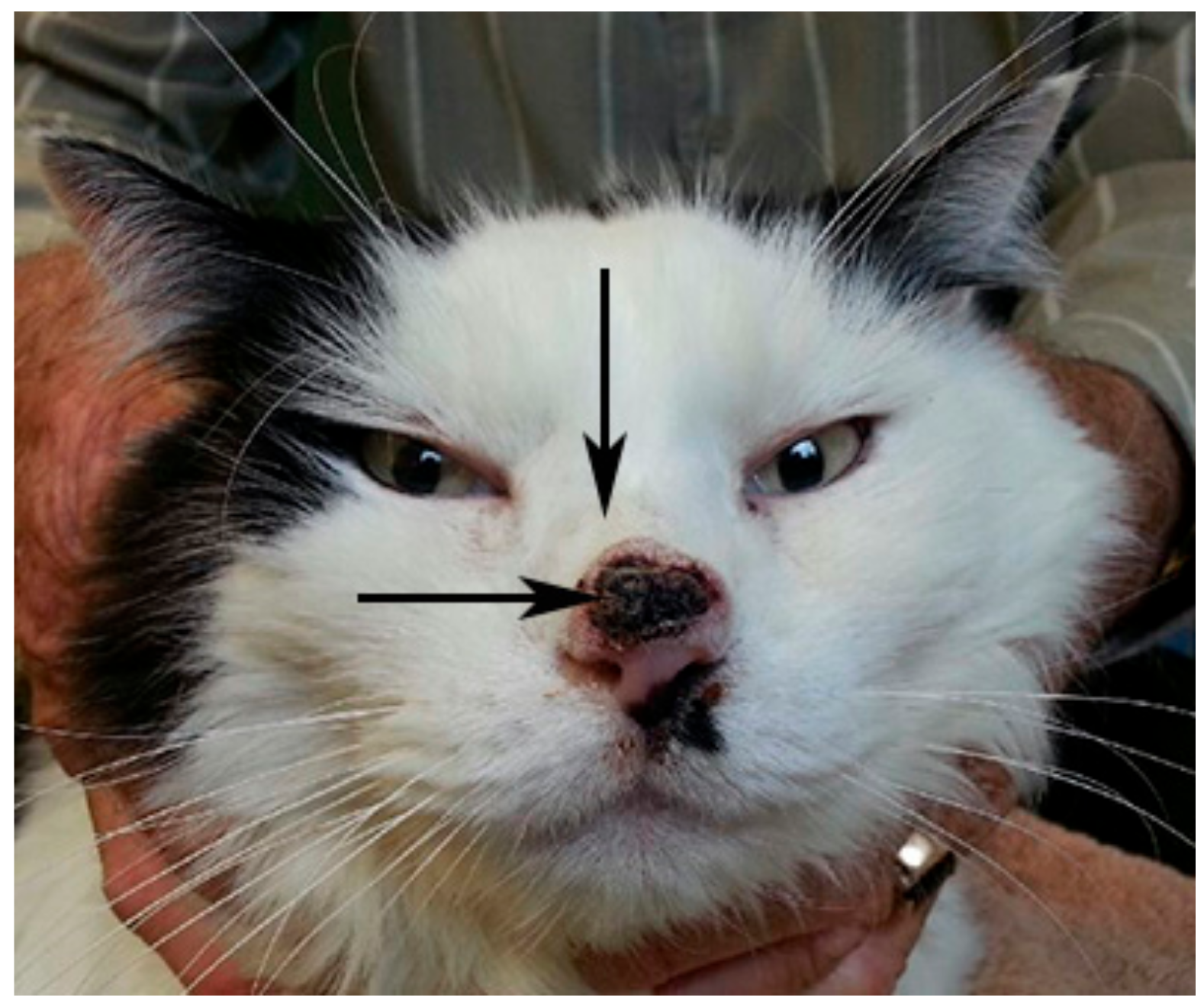
2.1. Clinical Samples
2.2. DNA Sequencing and Virus Discovery
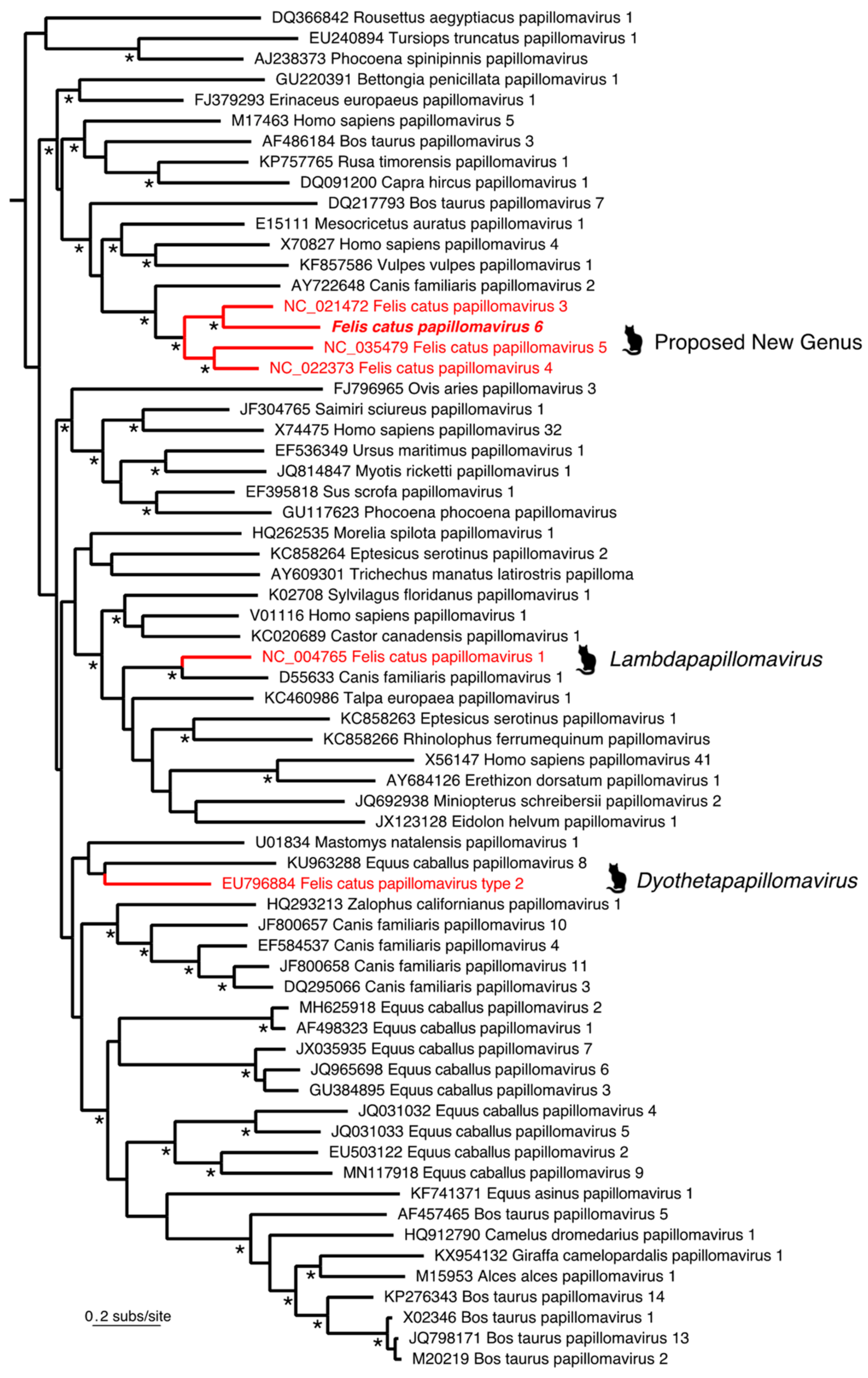
2.3. Phylogenetic Analysis
2.4. Investigation of Disease Association
2.4.1. DNA Extraction from FFPE SCC Biopsy
2.4.2. Papillomavirus-Specific PCR
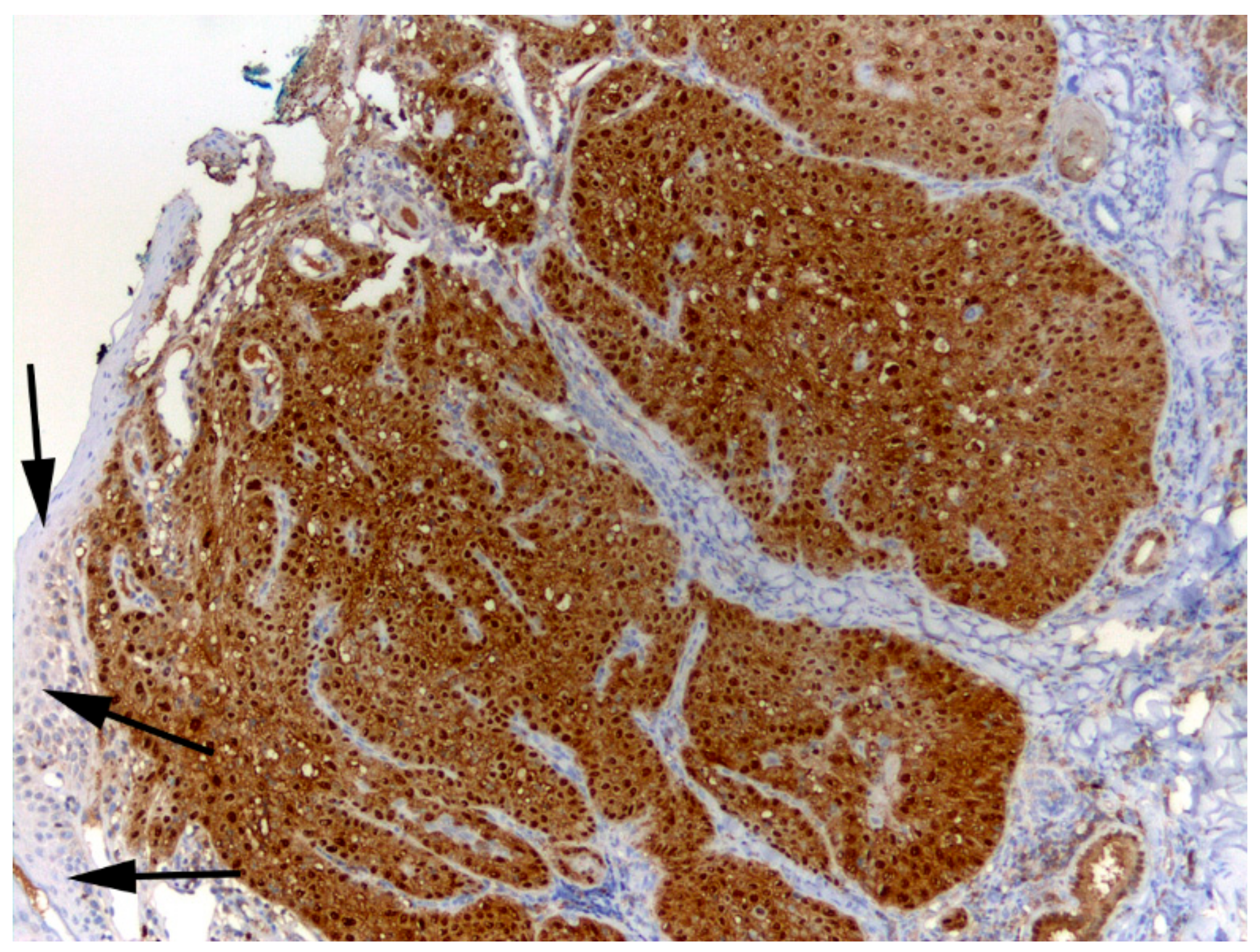
2.4.3. p16 Immunostaining
2.4.4. Papillomavirus Consensus PCR
3. Results and Discussion
3.1. Identification of Felis Catus Papillomavirus 6
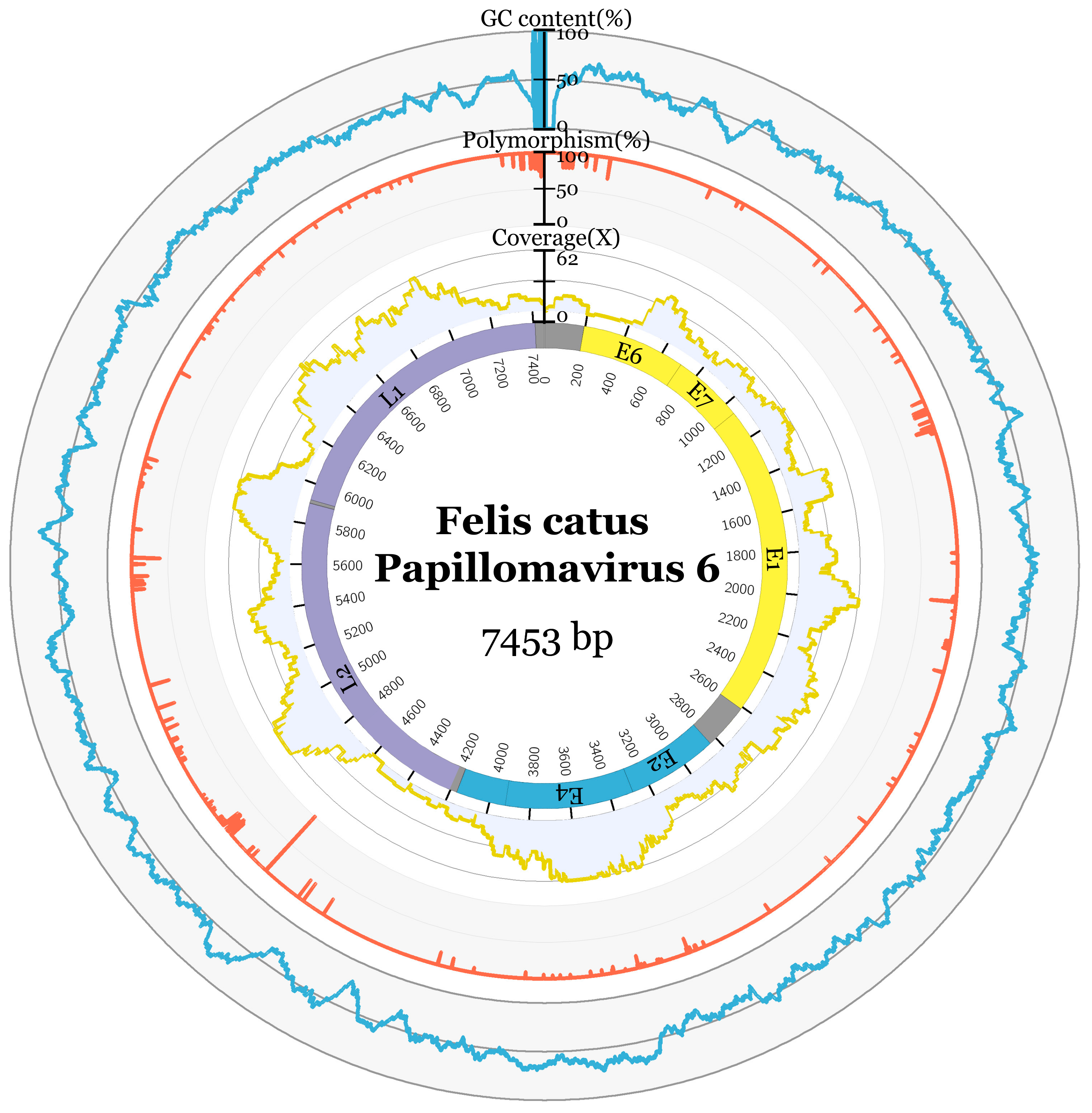
3.2. Genome Characteristics
3.3. Association of FcaPV6 with Squamous Cell Carcinoma
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McBride, A.A. Oncogenic human papillomaviruses. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, J.S.; Sharp, C.R.; Beatty, J.A. Novel viruses: Update on the significance of papillomavirus infections in cats. J. Feline Med. Surg. 2019, 21, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Dittmer, K.E.; Thomson, N.A.; Hills, S.F.; Laurie, R.E. Genomic characterisation of Felis catus papillomavirus type 5 with proposed classification within a new papillomavirus genus. Vet. Microbiol. 2017, 207, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; French, A.F. Felis catus papillomavirus types 1 and 4 are rarely present in neoplastic and inflammatory oral lesions of cats. Res. Vet. Sci. 2015, 100, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.L.; Ihrke, P.; Walder, E.J.; Affolter, V.K. (Eds.) Epidermal Tumors. In Skin Diseases of the Dog and Cat; Blackwell Publishing: Oxford, UK, 2005; pp. 559–603. [Google Scholar]
- Hoggard, N.; Munday, J.S.; Luff, J. Localization of Felis catus Papillomavirus Type 2 E6 and E7 RNA in Feline Cutaneous Squamous Cell Carcinoma. Vet. Pathol. 2018, 55, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altamura, G.; Corteggio, A.; Pacini, L.; Conte, A.; Pierantoni, G.M.; Tommasino, M.; Accardi, R.; Borzacchiello, G. Transforming properties of Felis catus papillomavirus type 2 E6 and E7 putative oncogenes in vitro and their transcriptional activity in feline squamous cell carcinoma in vivo. Virology 2016, 496, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5448. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, J.S.; Knight, C.G.; French, A.F. Evaluation of feline oral squamous cell carcinomas for p16CDKN2A protein immunoreactivity and the presence of papillomaviral DNA. Res. Vet. Sci. 2011, 90, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Kiupel, M.; French, A.F.; Howe, L. Amplification of papillomaviral DNA sequences from a high proportion of feline cutaneous in situ and invasive squamous cell carcinomas using a nested polymerase chain reaction. Vet. Dermatol. 2008, 19, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.-U.; Burk, R.D.; Chen, Z.; van Doorslaer, K.; Hausen, H.Z.; de Villiers, E.-M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, D.; Bates, S.; Mann, D.J.; Peters, G. Lack of cyclin D-Cdk complexes in Rb-negative cells correlates with high levels of p16INK4/MTS1 tumour suppressor gene product. EMBO J. 1995, 14, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Antonsson, A.; Erfurt, C.; Hazard, K.; Holmgren, V.; Simon, M.; Kataoka, A.; Hossain, S.; Hakangard, C.; Hansson, B.G. Prevalence and type spectrum of human papillomaviruses in healthy skin samples collected in three continents. J. Gen. Virol. 2003, 84 Pt 7, 1881–1886. [Google Scholar] [CrossRef]
- Munday, J.S.; Witham, A.I. Frequent detection of papillomavirus DNA in clinically normal skin of cats infected and noninfected with feline immunodeficiency virus. Vet. Dermatol. 2010, 21, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Intaraphet, S.; Farkas, D.K.; Johannesdottir Schmidt, S.A.; Cronin-Fenton, D.; Søgaard, M. Human papillomavirus infection and lymphoma incidence using cervical conization as a surrogate marker: A Danish nationwide cohort study. Hematol. Oncol. 2017, 35, 172–176. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | FcaPV2 | FcaPV1 | FcaPV3 | FcaPV6 | FcaPV5 | FcaPV4 |
|---|---|---|---|---|---|---|
| FcaPV2 | 55.7 | 56.8 | 55.2 | 56.1 | 55.1 | |
| FcaPV1 | 55.7 | 56.4 | 54.2 | 56.5 | 57.4 | |
| FcaPV3 | 56.8 | 56.4 | 66.1 | 62.8 | 63.8 | |
| FcaPV6 | 55.2 | 54.2 | 66.1 | 59.6 | 60.5 | |
| FcaPV5 | 56.1 | 56.5 | 62.8 | 59.6 | 64.4 | |
| FcaPV4 | 55.1 | 57.4 | 63.8 | 60.5 | 64.4 |
| ORF | Start | End | Length (nt) | Length (aa) | GC% |
|---|---|---|---|---|---|
| L1 | 5919 | 7409 | 1491 | 497 | 46.5 |
| L2 | 4261 | 5901 | 1641 | 547 | 54.4 |
| E1 | 1059 | 2858 | 1800 | 600 | 44.8 |
| E2 | 2836 | 4167 | 1332 | 444 | 51.2 |
| E4 | 3287 | 3922 | 636 | 292 | 57.4 |
| E6 | 195 | 785 | 591 | 197 | 48.2 |
| E7 | 372 | 707 | 1078 | 124 | 50.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrai, M.; Van Brussel, K.; Shi, M.; Li, C.-X.; Chang, W.-S.; Munday, J.S.; Voss, K.; McLuckie, A.; Taylor, D.; Laws, A.; et al. Identification of a Novel Papillomavirus Associated with Squamous Cell Carcinoma in a Domestic Cat. Viruses 2020, 12, 124. https://doi.org/10.3390/v12010124
Carrai M, Van Brussel K, Shi M, Li C-X, Chang W-S, Munday JS, Voss K, McLuckie A, Taylor D, Laws A, et al. Identification of a Novel Papillomavirus Associated with Squamous Cell Carcinoma in a Domestic Cat. Viruses. 2020; 12(1):124. https://doi.org/10.3390/v12010124
Chicago/Turabian StyleCarrai, Maura, Kate Van Brussel, Mang Shi, Ci-Xiu Li, Wei-Shan Chang, John S. Munday, Katja Voss, Alicia McLuckie, David Taylor, Andrew Laws, and et al. 2020. "Identification of a Novel Papillomavirus Associated with Squamous Cell Carcinoma in a Domestic Cat" Viruses 12, no. 1: 124. https://doi.org/10.3390/v12010124