Sulfonamide Inhibition Profile of the β-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions

 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Integrity of the Target Enzyme
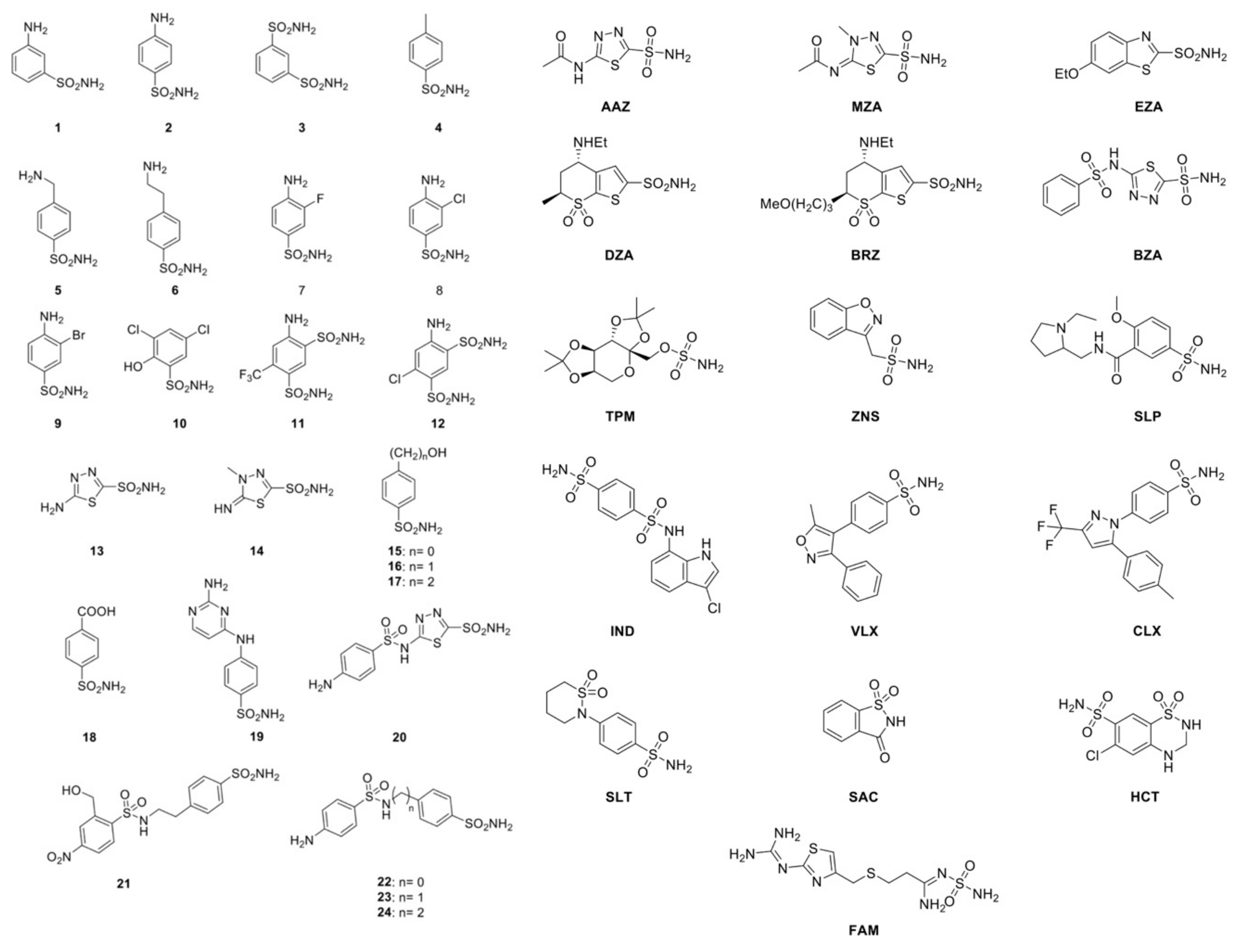
2.2. Sulfonamide Inhibition Profile
- Many of the investigated compounds, such as 3, 5, 6, 8, 9, 10, 12, 13, 14, 15, 17, 18, 23, 24, ZNS, and FAM, showed a weak MreCA inhibitory activity, with an inhibition constant (KI) higher than 10 μM. This is remarkable because most of these inhibitors block the human isoenzyme (hCA II) and the homologous fungal enzyme MgCA rather effectively. For example, 3, 5, 6, 8, 9, 10, 12, 15, 17, and 18 showed a KI in the nanomolar range of 63–174 nM for MgCA, while hCA II, with these compounds, was inhibited with a KI in the range of 8–170 nM. Intriguingly, hCA I was effectively inhibited by only two compounds on this list (3 and 18), with a KI in the range 68–79 nM.
- Several compounds of the series 1–24, such as 1, 2, 4, 7, 11, 16, 19, 21, and 22, had a moderate inhibitory effect on the MreCA, showing a KI between 3.74–7.79 μM. Most of these inhibitors were potent inhibitors of hCA II (KI = 11–300 nM) and weak inhibitors of hCA I (KI = 5.8–28 μM), except for compounds 19, 21, and 22 (KI = 16.4–109 nM). Similar behavior was shown by numerous clinically used compounds belonging to the series AZZ-HTC, such as MZA, EZA, BZA, TPM, SLP, CLX, SAC, HTC, and DCP. For these inhibitors, an inhibition constant ranging from 3.06 to 8.5 μM has been determined. Intriguingly, most of them are strong inhibitors of the two human isoenzymes (hCA I and hCA II), while most of those inhibitors, which resulted in moderate inhibitors of MreCA, were mild inhibitors of MgCA and vice-versa.
- Among all the compounds investigated for MreCA inhibition, only seven of them showed inhibition constants of < 1.0 μM. This is the case for inhibitors 20, DZA, BRZ, IND, VLX, SLT, and AAZ. These compounds had a KI in the range of 0.1–0.91 μM. It is interesting to note that the MreCA “strong inhibitors” were mild inhibitors of MgCA (KI = 31.5–79 μM), except for compound 20 (Table 3).
- As shown in Table 3, the two homologous fungal enzymes have an inhibition pattern very different from each other. Furthermore, the inhibition profiles of MreCA and MgCA were highly distinct from those of the two human isoenzymes.
3. Materials and Methods
3.1. Chemicals and Instruments
3.2. Enzyme Integrity Determination
3.3. Determination of the Kinetic Parameters and Inhibition Constants
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grimalt, R. A practical guide to scalp disorders. J. Investig. Dermatol. Symp. Proc. 2007, 12, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borda, L.J.; Wikramanayake, T.C. Seborrheic Dermatitis and Dandruff: A Comprehensive Review. J. Clin. Investig. Dermatol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Karakadze, M.A.; Hirt, P.A.; Wikramanayake, T.C. The genetic basis of seborrhoeic dermatitis: A review. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Paulino, L.C. New perspectives on dandruff and seborrheic dermatitis: Lessons we learned from bacterial and fungal skin microbiota. Eur. J. Dermatol. 2017, 27, 4–7. [Google Scholar] [CrossRef]
- Clavaud, C.; Jourdain, R.; Bar-Hen, A.; Tichit, M.; Bouchier, C.; Pouradier, F.; El Rawadi, C.; Guillot, J.; Menard-Szczebara, F.; Breton, L.; et al. Dandruff is associated with disequilibrium in the proportion of the major bacterial and fungal populations colonizing the scalp. PLoS ONE 2013, 8, e58203. [Google Scholar] [CrossRef]
- Turner, G.A.; Hoptroff, M.; Harding, C.R. Stratum corneum dysfunction in dandruff. Int. J. Cosmet. Sci. 2012, 34, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, R.J. Malassezia, dandruff and seborrhoeic dermatitis: An overview. Br. J. Dermatol. 2011, 165, 2–8. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Capasso, C.; Supuran, C.T. Cloning, Purification, and Characterization of a beta-Carbonic Anhydrase from Malassezia restricta, an Opportunistic Pathogen Involved in Dandruff and Seborrheic Dermatitis. Int. J. Mol. Sci. 2019, 20, 2447. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; De Luca, V.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Carginale, V.; Supuran, C.T.; Capasso, C. A new procedure for the cloning, expression and purification of the beta-carbonic anhydrase from the pathogenic yeast Malassezia globosa, an anti-dandruff drug target. J. Enzym. Inhib. Med. Chem. 2016, 31, 1156–1161. [Google Scholar] [CrossRef]
- Alterio, V.; Di Fiore, A.; D’Ambrosio, K.; Supuran, C.T.; De Simone, G. Multiple binding modes of inhibitors to carbonic anhydrases: How to design specific drugs targeting 15 different isoforms? Chem. Rev. 2012, 112, 4421–4468. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzym. Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.L.; Clement, R.; Kosta, A.; Maberly, S.C.; Gontero, B. A new widespread subclass of carbonic anhydrase in marine phytoplankton. ISME J. 2019, 13, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal theta-carbonic anhydrase critical for growth and photosynthesis in the marine diatom Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. An Overview of the Carbonic Anhydrases from Two Pathogens of the Oral Cavity: Streptococcus mutans and Porphyromonas gingivalis. Curr. Top. Med. Chem. 2016, 16, 2359–2368. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzym. Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. An Overview of the Selectivity and Efficiency of the Bacterial Carbonic Anhydrase Inhibitors. Curr. Med. Chem. 2015, 22, 2130–2139. [Google Scholar] [CrossRef]
- Prete, S.D.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Anion Inhibition Profile of the beta-Carbonic Anhydrase from the Opportunist Pathogenic Fungus Malassezia Restricta Involved in Dandruff and Seborrheic Dermatitis. Metabolites 2019, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, A.; Bua, S.; Del Prete, S.; Heravi, Y.E.; Saboury, A.A.; Karioti, A.; Bilia, A.R.; Capasso, C.; Gratteri, P.; Supuran, C.T. Natural Polyphenols Selectively Inhibit beta-Carbonic Anhydrase from the Dandruff-Producing Fungus Malassezia globosa: Activity and Modeling Studies. Chem. Med. Chem. 2018, 13, 816–823. [Google Scholar] [CrossRef]
- Nocentini, A.; Vullo, D.; Del Prete, S.; Osman, S.M.; Alasmary, F.A.S.; AlOthman, Z.; Capasso, C.; Carta, F.; Gratteri, P.; Supuran, C.T. Inhibition of the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa with monothiocarbamates. J. Enzym. Inhib. Med. Chem. 2017, 32, 1064–1070. [Google Scholar] [CrossRef] [Green Version]
- Entezari Heravi, Y.; Bua, S.; Nocentini, A.; Del Prete, S.; Saboury, A.A.; Sereshti, H.; Capasso, C.; Gratteri, P.; Supuran, C.T. Inhibition of Malassezia globosa carbonic anhydrase with phenols. Bioorg. Med. Chem. 2017, 25, 2577–2582. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. Bacterial, fungal and protozoan carbonic anhydrases as drug targets. Expert Opin. Ther. Targets 2015, 19, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Klengel, T.; Liang, W.J.; Chaloupka, J.; Ruoff, C.; Schroppel, K.; Naglik, J.R.; Eckert, S.E.; Mogensen, E.G.; Haynes, K.; Tuite, M.F.; et al. Fungal adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence. Curr. Biol. 2005, 15, 2021–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitson, K.S.; Vullo, D.; Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Molecular cloning, characterization, and inhibition studies of a beta-carbonic anhydrase from Malassezia globosa, a potential antidandruff target. J. Med. Chem. 2012, 55, 3513–3520. [Google Scholar] [CrossRef]
- Stalhberger, T.; Simenel, C.; Clavaud, C.; Eijsink, V.G.; Jourdain, R.; Delepierre, M.; Latge, J.P.; Breton, L.; Fontaine, T. Chemical organization of the cell wall polysaccharide core of Malassezia restricta. J. Biol. Chem. 2014, 289, 12647–12656. [Google Scholar] [CrossRef] [Green Version]
- Morand, S.C.; Bertignac, M.; Iltis, A.; Kolder, I.; Pirovano, W.; Jourdain, R.; Clavaud, C. Complete Genome Sequence of Malassezia restricta CBS 7877, an Opportunist Pathogen Involved in Dandruff and Seborrheic Dermatitis. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Angeli, A.; Pinteala, M.; Maier, S.S.; Del Prete, S.; Capasso, C.; Simionescu, B.C.; Supuran, C.T. Inhibition of alpha-, beta-, gamma-, delta-, zeta- and eta-class carbonic anhydrases from bacteria, fungi, algae, diatoms and protozoans with famotidine. J. Enzym. Inhib. Med. Chem. 2019, 34, 644–650. [Google Scholar] [CrossRef] [Green Version]
- Elleuche, S.; Poggeler, S. Carbonic anhydrases in fungi. Microbiology 2010, 156, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Lehneck, R.; Poggeler, S.; Supuran, C.T. Sulfonamide inhibition studies of two beta-carbonic anhydrases from the ascomycete fungus Sordaria macrospora, CAS1 and CAS2. J. Enzym. Inhib. Med. Chem. 2018, 33, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Anion inhibition studies of the dandruff-producing fungus Malassezia globosa beta-carbonic anhydrase MgCA. Bioorg. Med. Chem. Lett. 2015, 25, 5194–5198. [Google Scholar] [CrossRef]
- Singh, S.; Supuran, C.T. In silico modeling of beta-carbonic anhydrase inhibitors from the fungus Malassezia globosa as antidandruff agents. J. Enzym. Inhib. Med. Chem. 2016, 31, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Schlicker, C.; Hall, R.A.; Vullo, D.; Middelhaufe, S.; Gertz, M.; Supuran, C.T.; Muhlschlegel, F.A.; Steegborn, C. Structure and inhibition of the CO2-sensing carbonic anhydrase Can2 from the pathogenic fungus Cryptococcus neoformans. J. Mol. Biol. 2009, 385, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. How many carbonic anhydrase inhibition mechanisms exist? J. Enzym. Inhib. Med. Chem. 2016, 31, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Monti, S.M.; Maresca, A.; Viparelli, F.; Carta, F.; De Simone, G.; Muhlschlegel, F.A.; Scozzafava, A.; Supuran, C.T. Dithiocarbamates are strong inhibitors of the beta-class fungal carbonic anhydrases from Cryptococcus neoformans, Candida albicans and Candida glabrata. Bioorg. Med. Chem. Lett. 2012, 22, 859–862. [Google Scholar] [CrossRef] [Green Version]
- Carta, F.; Innocenti, A.; Hall, R.A.; Muhlschlegel, F.A.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzymes from the fungal pathogens Candida albicans and Cryptococcus neoformans with branched aliphatic/aromatic carboxylates and their derivatives. Bioorg. Med. Chem. Lett. 2011, 21, 2521–2526. [Google Scholar] [CrossRef]
- Vullo, D.; Del Prete, S.; Nocentini, A.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Bozdag, M.; Carta, F.; Gratteri, P.; Supuran, C.T. Dithiocarbamates effectively inhibit the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa. Bioorg. Med. Chem. 2017, 25, 1260–1265. [Google Scholar] [CrossRef]
- De Luca, V.; Del Prete, S.; Supuran, C.T.; Capasso, C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J. Enzym. Inhib. Med. Chem. 2015, 30, 277–282. [Google Scholar] [CrossRef]
- Guzel, O.; Maresca, A.; Hall, R.A.; Scozzafava, A.; Mastrolorenzo, A.; Muhlschlegel, F.A.; Supuran, C.T. Carbonic anhydrase inhibitors. The beta-carbonic anhydrases from the fungal pathogens Cryptococcus neoformans and Candida albicans are strongly inhibited by substituted-phenyl-1H-indole-5-sulfonamides. Bioorg. Med. Chem. Lett. 2010, 20, 2508–2511. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug. Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef]
- Angeli, A.; Ferraroni, M.; Supuran, C.T. Famotidine, an Antiulcer Agent, Strongly Inhibits Helicobacter pylori and Human Carbonic Anhydrases. ACS Med. Chem. Lett. 2018, 9, 1035–1038. [Google Scholar] [CrossRef]
- Supuran, C.T. Advances in structure-based drug discovery of carbonic anhydrase inhibitors. Expert Opin. Drug. Discov. 2017, 12, 61–88. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; Del Prete, S.; Fisher, G.M.; Andrews, K.T.; Poulsen, S.A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the eta-class carbonic anhydrase from the malaria pathogen Plasmodium falciparum. Bioorg. Med. Chem. 2015, 23, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the Antarctic bacterium Pseudoalteromonas haloplanktis. Bioorg. Med. Chem. Lett. 2015, 25, 3550–3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the Antarctic cyanobacterium Nostoc commune. Bioorg. Med. Chem. 2015, 23, 1728–1734. [Google Scholar] [CrossRef]
- Dedeoglu, N.; DeLuca, V.; Isik, S.; Yildirim, H.; Kockar, F.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition study of the beta-class carbonic anhydrase from the caries producing pathogen Streptococcus mutans. Bioorg. Med. Chem. Lett. 2015, 25, 2291–2297. [Google Scholar] [CrossRef]
- Alafeefy, A.M.; Ceruso, M.; Al-Tamimi, A.M.; Del Prete, S.; Supuran, C.T.; Capasso, C. Inhibition studies of quinazoline-sulfonamide derivatives against the gamma-CA (PgiCA) from the pathogenic bacterium, Porphyromonas gingivalis. J. Enzym. Inhib. Med. Chem. 2015, 30, 592–596. [Google Scholar] [CrossRef]
- Alafeefy, A.M.; Abdel-Aziz, H.A.; Vullo, D.; Al-Tamimi, A.M.; Awaad, A.S.; Mohamed, M.A.; Capasso, C.; Supuran, C.T. Inhibition of human carbonic anhydrase isozymes I, II, IX and XII with a new series of sulfonamides incorporating aroylhydrazone-, [1,2,4] triazolo [3,4-b] [1,3,4]t hiadiazinyl- or 2-(cyanophenylmethylene)-1, 3, 4-thiadiazol-3(2H)-yl moieties. J. Enzym. Inhib. Med. Chem. 2015, 30, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.R.; Fernandez Baldo, M.; Echeverria, G.; Baldoni, H.; Vullo, D.; Soria, D.B.; Supuran, C.T.; Cami, G.E. A substituted sulfonamide and its Co (II), Cu (II), and Zn (II) complexes as potential antifungal agents. J. Enzym. Inhib. Med. Chem. 2016, 31, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Comparison of the sulfonamide inhibition profiles of the alpha-, beta- and gamma-carbonic anhydrases from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. Lett. 2016, 26, 1941–1946. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, expression, purification and sulfonamide inhibition profile of the complete domain of the eta-carbonic anhydrase from Plasmodium falciparum. Bioorg. Med. Chem. Lett. 2016, 26, 4184–4190. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Ferraroni, M.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition studies of the beta-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. 2016, 24, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Abdel Gawad, N.M.; Amin, N.H.; Elsaadi, M.T.; Mohamed, F.M.; Angeli, A.; De Luca, V.; Capasso, C.; Supuran, C.T. Synthesis of 4-(thiazol-2-ylamino)-benzenesulfonamides with carbonic anhydrase I, II and IX inhibitory activity and cytotoxic effects against breast cancer cell lines. Bioorg. Med. Chem. 2016, 24, 3043–3051. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Legionella pneumophila Carbonic Anhydrases: Underexplored Antibacterial Drug Targets. Pathogens 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimori, I.; Vullo, D.; Minakuchi, T.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of two beta-carbonic anhydrases from the bacterial pathogen Legionella pneumophila. Bioorg. Med. Chem. 2014, 22, 2939–2946. [Google Scholar] [CrossRef]
- Vullo, D.; Sai Kumar, R.S.; Scozzafava, A.; Capasso, C.; Ferry, J.G.; Supuran, C.T. Anion inhibition studies of a beta-carbonic anhydrase from Clostridium perfringens. Bioorg. Med. Chem. Lett. 2013, 23, 6706–6710. [Google Scholar] [CrossRef]
- Nishimori, I.; Minakuchi, T.; Maresca, A.; Carta, F.; Scozzafava, A.; Supuran, C.T. The beta-carbonic anhydrases from Mycobacterium tuberculosis as drug targets. Curr. Pharm. Des. 2010, 16, 3300–3309. [Google Scholar] [CrossRef]
- Carta, F.; Maresca, A.; Covarrubias, A.S.; Mowbray, S.L.; Jones, T.A.; Supuran, C.T. Carbonic anhydrase inhibitors. Characterization and inhibition studies of the most active beta-carbonic anhydrase from Mycobacterium tuberculosis, Rv3588c. Bioorg. Med. Chem. Lett. 2009, 19, 6649–6654. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibition and the management of neuropathic pain. Expert Rev. Neurother. 2016, 16, 961–968. [Google Scholar] [CrossRef]
- Supuran, C.T. Drug interaction considerations in the therapeutic use of carbonic anhydrase inhibitors. Expert Opin. Drug. Metab. Toxicol. 2016, 12, 423–431. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Iandolo, E.; Supuran, C.T.; Capasso, C. Protonography, a powerful tool for analyzing the activity and the oligomeric state of the gamma-carbonic anhydrase identified in the genome of Porphyromonas gingivalis. Bioorg. Med. Chem. 2015, 23, 3747–3750. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Supuran, C.T.; Capasso, C. Protonography, a technique applicable for the analysis of eta-carbonic anhydrase activity. J. Enzym. Inhib. Med. Chem. 2015, 30, 920–924. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; Caminiti-Segonds, N.; Zoccola, D.; Tambutte, S.; Supuran, C.T.; Capasso, C. Protonography and anion inhibition profile of the alpha-carbonic anhydrase (CruCA4) identified in the Mediterranean red coral Corallium rubrum. Bioorg. Chem. 2018, 76, 281–287. [Google Scholar] [CrossRef]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; di Fonzo, P.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Anion inhibition profiles of alpha-, beta- and gamma-carbonic anhydrases from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. 2016, 24, 3413–3417. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; di Fonzo, P.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Anion inhibition profiles of the complete domain of the eta-carbonic anhydrase from Plasmodium falciparum. Bioorg. Med. Chem. 2016, 24, 4410–4414. [Google Scholar] [CrossRef]
- De Luca, V.; Vullo, D.; Del Prete, S.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, characterization and anion inhibition studies of a gamma-carbonic anhydrase from the Antarctic bacterium Colwellia psychrerythraea. Bioorg. Med. Chem. 2016, 24, 835–840. [Google Scholar] [CrossRef]




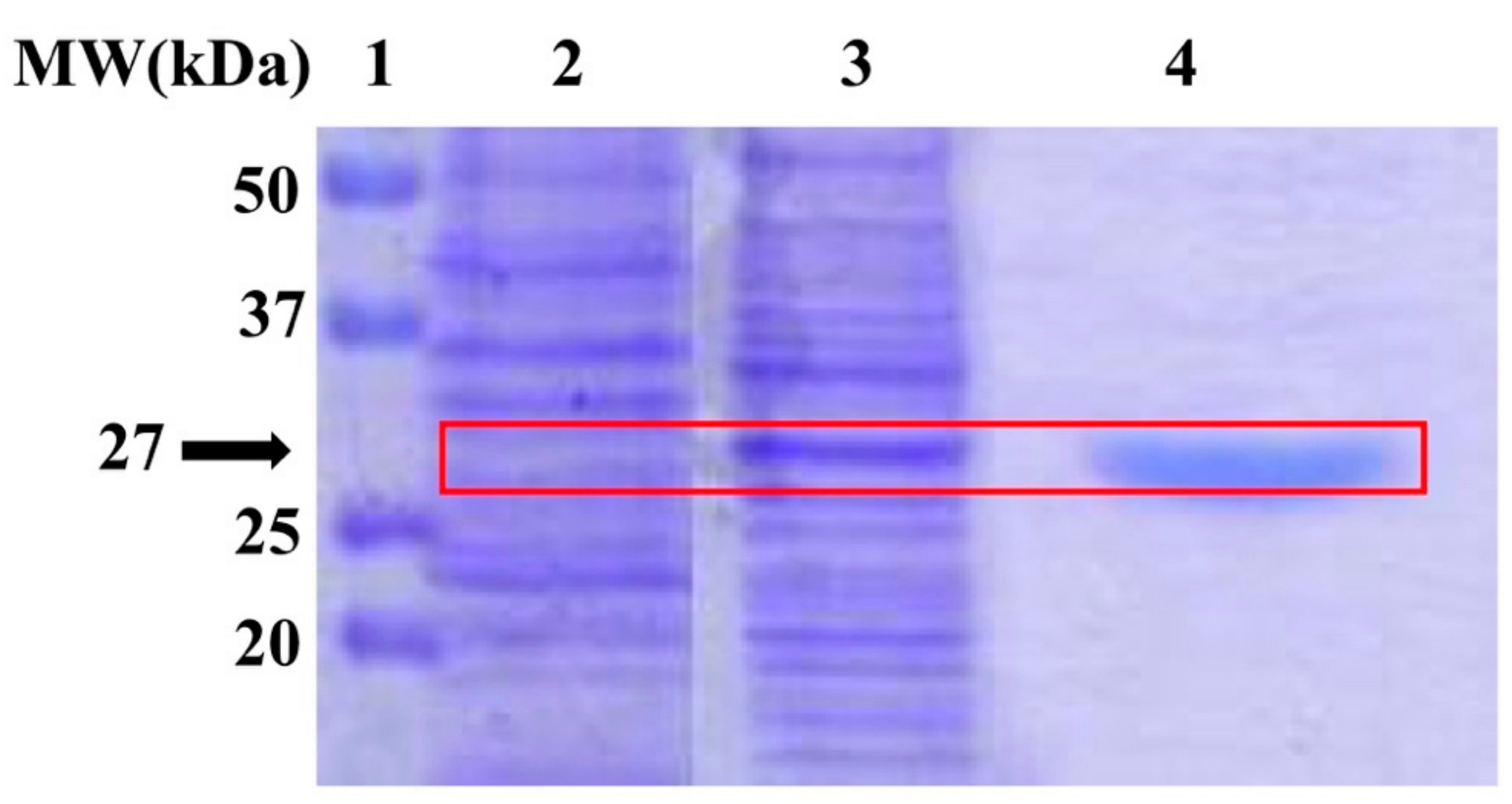{kind=link}
{kind=link}
{kind=link}
| Organisms | Acronym | Class | Kcat (s−1) | kcat/KM (M−1·s−1) |
|---|---|---|---|---|
| Homo sapiens | hCA I a | 𝛼 | 2.0 × 105 | 5.0 × 107 |
| hCA II a | 𝛼 | 1.4 × 106 | 1.5 × 108 | |
| Malassezia retricta | MreCA | β | 1.06 × 106 | 1.07 × 108 |
| Malassezia globosa | MgCA b | β | 9.2 × 105 | 8.3 × 107 |
| Cryptococcus neoformans | Can2 c | β | 3.9 × 105 | 4.3 × 107 |
| Candida albicans | CaNce103 d | β | 8.0 × 105 | 9.7 × 107 |
| Candida glabrata | CgNce103 e | β | 3.8 × 105 | 4.8 × 107 |
| CAI Acronym | Commercial Name |
|---|---|
| AAZ | Acetazolamide |
| MZA | Methazolamide |
| EZA | Ethoxzolamide |
| DCP | Dichlorophenamide |
| DZA | Dorzolamide |
| BRZ | Brinzolamide |
| BZA | Benzolamide |
| ZNS | Zonisamide |
| SLT | Sulthiame |
| FAM | Famotidine |
| TPM | Topiramate |
| SLP | Sulpiride |
| IND | Indisulam |
| CLX | Celecoxib |
| VLX | Valdecoxib |
| SAC | Saccharin |
| HCT | Hydrochlorothiazide |
| KI (µM) * | ||||
|---|---|---|---|---|
| Compound | MreCA | MgCA a | hCA I b | hCA II b |
| 1 | 4.12 | 9.8 | 28.0 | 0.300 |
| 2 | 4.62 | 0.245 | 25.0 | 0.240 |
| 3 | >10 | 0.152 | 0.079 | 0.008 |
| 4 | 4.04 | 6.74 | 78.5 | 0.320 |
| 5 | >10 | 0.174 | 25.0 | 0.170 |
| 6 | >10 | 0.079 | 21.0 | 0.160 |
| 7 | 4.59 | 0.116 | 8.3 | 0.060 |
| 8 | >10 | 0.121 | 9.8 | 0.110 |
| 9 | >10 | 0.349 | 6.5 | 0.040 |
| 10 | >10 | 0.543 | 7.3 | 0.054 |
| 11 | 6.76 | 0.09 | 5.8 | 0.063 |
| 12 | >10 | 0.092 | 8.4 | 0.075 |
| 13 | >10 | 79.0 | 8.6 | 0.060 |
| 14 | >10 | 85.0 | 9.3 | 0.019 |
| 15 | >10 | 0.236 | 5.5 | 0.080 |
| 16 | 6.51 | 0.104 | 9.5 | 0.094 |
| 17 | >10 | 0.063 | 21.0 | 0.125 |
| 18 | >10 | 0.068 | 0.164 | 0.046 |
| 19 | 7.79 | 35.0 | 0.109 | 0.033 |
| 20 | 0.91 | 0.234 | 0.006 | 0.002 |
| 21 | 7.40 | 0.118 | 0.069 | 0.011 |
| 22 | 3.74 | 0.094 | 0.0164 | 0.046 |
| 23 | >10 | 45.3 | 0.109 | 0.033 |
| 24 | >10 | 2.56 | 0.095 | 0.030 |
| MZA | 3.90 | 74.55 | 0.05 | 0.014 |
| EZA | 3.79 | 38.0 | 0.025 | 0.008 |
| DZA | 0.81 | 79.0 | 50.0 | 0.009 |
| BRZ | 0.70 | 84.0 | 45.0 | 0.003 |
| BZA | 7.15 | 0.482 | 0.015 | 0.009 |
| TPM | 3.83 | 1.46 | 0.25 | 0.01 |
| SLP | 4.85 | 0.32 | 1.2 | 0.04 |
| IND | 0.87 | n.d. | 0.031 | 0.015 |
| ZNS | >10 | 7.65 | 0.056 | 0.035 |
| CLX | 1.4 | 34.8 | 50.0 | 0.021 |
| VLX | 0.77 | 31.5 | 54.0 | 0.043 |
| SLT | 0.67 | n.d. | 0.374 | 0.009 |
| SAC | 6.2 | n.d. | 18.54 | 5.959 |
| HCT | 8.5 | n.d. | 0.328 | 0.29 |
| FAM | >10 | n.d. | 0.922 c | 0.058 c |
| DCP | 3.06 | 0.346 | 1.2 | 0.038 |
| AAZ | 0.1 | 76.0 | 0.25 | 0.012 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Prete, S.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Sulfonamide Inhibition Profile of the β-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions. Metabolites 2020, 10, 39. https://doi.org/10.3390/metabo10010039
Del Prete S, Angeli A, Ghobril C, Hitce J, Clavaud C, Marat X, Supuran CT, Capasso C. Sulfonamide Inhibition Profile of the β-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions. Metabolites. 2020; 10(1):39. https://doi.org/10.3390/metabo10010039
Chicago/Turabian StyleDel Prete, Sonia, Andrea Angeli, Cynthia Ghobril, Julien Hitce, Cécile Clavaud, Xavier Marat, Claudiu T. Supuran, and Clemente Capasso. 2020. "Sulfonamide Inhibition Profile of the β-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions" Metabolites 10, no. 1: 39. https://doi.org/10.3390/metabo10010039