Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
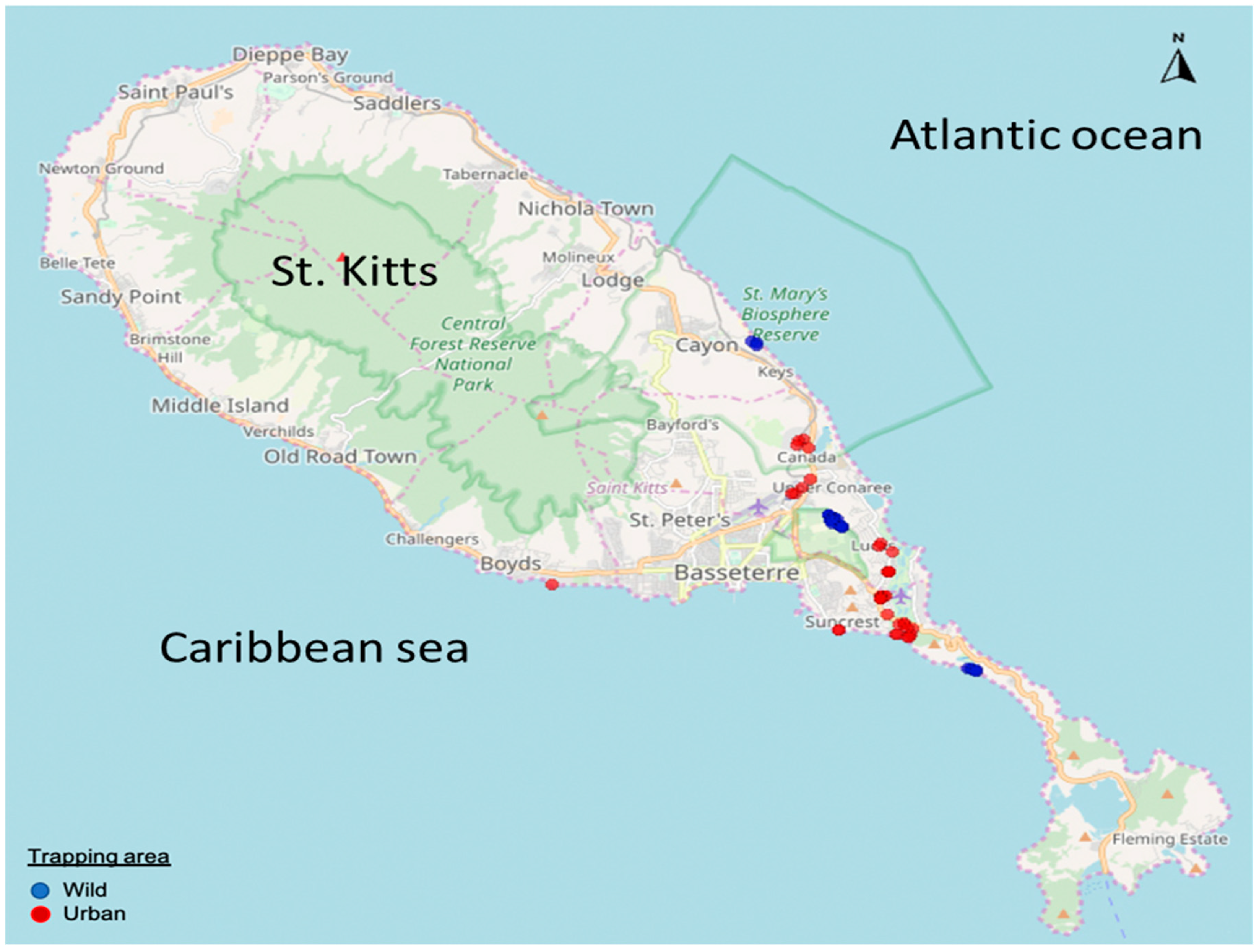
2.2. Sampling
2.3. Screening for PBVs
2.4. Amplification of Complete/Nearly Complete Gene Segment-2 of Mongoose PBV GI Strains
2.5. Amplification of the Nearly Full-Length RdRp Genes of a Novel PBV-Like Strain and a PBV GI Strain from the Same Fecal Sample
2.6. Nucleotide Sequencing
2.7. Sequence Analysis
2.8. GenBank Accession Numbers
3. Results and Discussion
3.1. Detection of PBVs in Mongooses
3.2. Analysis of Partial Gene Segment-2 of Mongoose PBV GI Strains
3.3. Molecular Characterization of Complete/Nearly Complete Gene Segment-2 of Mongoose PBV GI Strains
3.4. Identification of a Novel PBV-Like RdRp Gene Sequence That Uses an Alternative Genetic Code for Translation
3.5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delmas, B.; Attoui, H.; Ghosh, S.; Malik, Y.S.; Mundt, E.; Vakharia, V.N. Ictv virus taxonomy profile: Picobirnaviridae. J. Gen. Virol. 2019, 100, 133–134. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Teng, J.L.L.; Bai, R.; Tang, Y.; Wong, A.Y.P.; Li, K.S.M.; Lam, C.S.F.; Fan, R.Y.Y.; Lau, S.K.P.; Yuen, K.Y. Novel Picobirnaviruses in Respiratory and Alimentary Tracts of Cattle and Monkeys with Large Intra- and Inter-Host Diversity. Viruses 2019, 11, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, S.L.; Poon, L.L.M.; van Leeuwen, M.; Lau, P.N.; Perera, H.K.K.; Peiris, J.S.M.; Simon, J.H.; Osterhaus, A.D.M.E. Genogroup I and II picobirnaviruses in respiratory tracts of pigs. Emerg. Infect. Dis. 2011, 17, 2328–2330. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; van Leeuwen, M.; Schapendonk, C.M.E.; Schürch, A.C.; Bodewes, R.; Haagmans, B.L.; Osterhaus, A.D.M.E. Picobirnaviruses in the human respiratory tract. Emerg. Infect. Dis. 2012, 18, 1539–1540. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.S.; Kumar, N.; Sharma, K.; Dhama, K.; Shabbir, M.Z.; Ganesh, B.; Kobayashi, N.; Banyai, K. Epidemiology, phylogeny, and evolution of emerging enteric picobirnaviruses of animal origin and their relationship to human strains. Biomed. Res. Int. 2014, 2014, 780752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, B.; Masachessi, G.; Mladenova, Z. Animal Picobirnavirus. Virus Dis. 2014, 25, 223–238. [Google Scholar] [CrossRef]
- Krishnamurthy, S.R.; Wang, D. Extensive conservation of prokaryotic ribosomal binding sites in known and novel picobirnaviruses. Virology 2018, 516, 108–114. [Google Scholar] [CrossRef]
- Boros, Á.; Polgár, B.; Pankovics, P.; Fenyvesi, H.; Engelmann, P.; Phan, T.G.; Delwart, E.; Reuter, G. Multiple divergent picobirnaviruses with functional prokaryotic Shine-Dalgarno ribosome binding sites present in cloacal sample of a diarrheic chicken. Virology 2018, 525, 62–72. [Google Scholar] [CrossRef]
- Wolf, Y.I.; Kazlauskas, D.; Iranzo, J.; Lucía-Sanz, A.; Kuhn, J.H.; Krupovic, M.; Dolja, V.V.; Koonin, E.V. Origins and evolution of the global RNA virome. MBio 2018, 9, e02329-18. [Google Scholar] [CrossRef] [Green Version]
- Yinda, C.K.; Ghogomu, S.M.; Conceição-Neto, N.; Beller, L.; Deboutte, W.; Vanhulle, E.; Maes, P.; Van Ranst, M.; Matthijnssens, J. Cameroonian fruit bats harbor divergent viruses, including rotavirus H, bastroviruses, and picobirnaviruses using an alternative genetic code. Virus Evol. 2018, 4, vey008. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Knox, M.A.; Gedye, K.R.; Hayman, D.T.S. The challenges of analysing highly diverse picobirnavirus sequence data. Viruses 2018, 10, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Shiokawa, K.; Aung, M.S.; Malik, Y.S.; Kobayashi, N. High detection rates of picobirnaviruses in free roaming rats (Rattus spp.): Molecular characterization of complete gene segment-2. Infect. Genet. Evol. 2018, 65, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.; Yibin, C.; Nair, R.; Peda, A.; Aung, M.S.; Ketzis, J.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Molecular characterization of complete genomic segment-2 of picobirnavirus strains detected in a cat and a dog. Infect. Genet. Evol. 2017, 54, 200–204. [Google Scholar] [CrossRef]
- Gallagher, C.A.; Navarro, R.; Cruz, K.; Aung, M.S.; Ng, A.; Bajak, E.; Beierschmitt, A.; Lawrence, M.; Dore, K.M.; Ketzis, J.; et al. Detection of picobirnaviruses in vervet monkeys (Chlorocebus sabaeus): Molecular characterization of complete genomic segment-2. Virus Res. 2017, 230, 13–18. [Google Scholar] [CrossRef]
- Ghosh, S.; Kobayashi, N.; Nagashima, S.; Naik, T.N. Molecular characterization of full-length genomic segment 2 of a bovine picobirnavirus (PBV) strain: Evidence for high genetic diversity with genogroup I PBVs. J. Gen. Virol. 2009, 90, 2519–2524. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Bai, R.; Teng, J.L.L.; Lee, P.; Martelli, P.; Hui, S.-W.; Yuen, K.-Y. Complete Genome Sequence of a Novel Picobirnavirus, Otarine Picobirnavirus, Discovered in California Sea Lions. J. Virol. 2012, 86, 6377–6378. [Google Scholar] [CrossRef] [Green Version]
- Verma, H.; Mor, S.K.; Erber, J.; Goyal, S.M. Prevalence and complete genome characterization of turkey picobirnaviruses. Infect. Genet. Evol. 2015, 30, 134–139. [Google Scholar] [CrossRef]
- Wakuda, M.; Pongsuwanna, Y.; Taniguchi, K. Complete nucleotide sequences of two RNA segments of human picobirnavirus. J. Virol. Methods 2005, 126, 165–169. [Google Scholar] [CrossRef]
- Conceição-Neto, N.; Mesquita, J.R.; Zeller, M.; Yinda, C.K.; Álvares, F.; Roque, S.; Petrucci-Fonseca, F.; Godinho, R.; Heylen, E.; Van Ranst, M.; et al. Reassortment among picobirnaviruses found in wolves. Arch. Virol. 2016, 161, 2859–2862. [Google Scholar] [CrossRef] [Green Version]
- Barun, A.; Hanson, C.C.; Campell, K.J.; Simberloff, D. A review of small Indian mongoose management and eradications on islands. In Island Invasives: Eradication and Management; Veitch, C.R., Clout, M.N., Towns, D.R., Eds.; IUCN: Gland, Switzerland, 2011; pp. 17–25. [Google Scholar]
- Veron, G.; Jennings, A.P. Javan mongoose or small Indian mongoose—Who is where? Mamm. Biol. 2017, 87, 62–70. [Google Scholar] [CrossRef]
- Rosen, B.I.; Fang, Z.Y.; Glass, R.I.; Monroe, S.S. Cloning of human picobirnavirus genomic segments and development of an RT-PCR detection assay. Virology 2000, 277, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Malik, Y.S.; Sircar, S.; Dhama, K.; Singh, R.; Ghosh, S.; Bányai, K.; Vlasova, A.N.; Nadia, T.; Singh, R.K. Molecular epidemiology and characterization of picobirnaviruses in small ruminant populations in India. Infect. Genet. Evol. 2018, 63, 39–42. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, T.; Halper, B.; Siebert, J.; Cruz-Martinez, L.; Chapwanya, A.; Kelly, P.; Ketzis, J.K.; Vessell, J.; Köster, L.; Yao, C. Parasites of small Indian mongoose, Herpestes auropunctatus, on St. Kitts, West Indies. Parasitol. Res. 2018, 117, 989–994. [Google Scholar] [CrossRef] [Green Version]
- Collier, A.M.; Lyytinen, O.L.; Guo, Y.R.; Toh, Y.; Poranen, M.M.; Tao, Y.J. Initiation of RNA Polymerization and Polymerase Encapsidation by a Small dsRNA Virus. PLoS Pathog. 2016, 12, e1005523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shackelton, L.A.; Holmes, E.C. The role of alternative genetic codes in viral evolution and emergence. J. Theor. Biol. 2008, 254, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.; Eusebio-Cope, A.; Kondo, H.; Hillman, B.I.; Suzuki, N. Investigation of Host Range of and Host Defense against a Mitochondrially Replicating Mitovirus. J. Virol. 2019, 93, e01503-18. [Google Scholar] [CrossRef] [Green Version]
- Bányai, K.; Potgieter, C.; Gellért, Á.; Ganesh, B.; Tempesta, M.; Lorusso, E.; Buonavoglia, C.; Martella, V. Genome sequencing identifies genetic and antigenic divergence of porcine picobirnaviruses. J. Gen. Virol. 2014, 95, 2233–2239. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Length of Nucleotide (nt) Sequence Obtained | Putative Open Reading Frame Encoding RdRp | Length of Putative RdRp | Maximum Deduced aa Identity (%) of Putative RdRp | |
|---|---|---|---|---|---|
| With Other Mongoose PBV Strains 1 | With PBV Strains from Other Host Species 2, (GenBank Accession Number) | ||||
| PBV-like/Mongoose/KNA/M17A/2017 | 1662 nt 3 | nt 107-nt 1639 4 | 510 aa 4 | 48% with PBV/Mongoose/KNA/M33/2017 and PBV/Mongoose/KNA/M45/2017 | 51% with PBV/Marmot/CHN/c159756/2013, (AVX53249) |
| PBV/Mongoose/KNA/M17B/2017 | 1715 nt 3 | nt 47-nt 1705 | 552 aa | 64% with PBV/Mongoose/KNA/M33/2017 | 69% with PBV/Marmot/CHN/c206099/2013, (AVX53281) |
| PBV/Mongoose/KNA/M33/2017 | 1701 nt 3 | nt 105-nt 1691 | 528 aa | 68% with PBV/Mongoose/KNA/M45/2017, PBV/Mongoose/KNA/M58/2017 and PBV/Mongoose/KNA/M67/2017 | 71% with PBV/Cat/KNA/K40/2014, (ASK85695) |
| PBV/Mongoose/KNA/M45/2017 | 1651 nt 3 | nt 47-nt 1639 | 530 aa | 99% with PBV/Mongoose/KNA/M58/2017 and PBV/Mongoose/KNA/M67/2017 | 78% with PBV/Gorilla/COG/141-RdRp1718/2015, (AVA30701) |
| PBV/Mongoose/KNA/M46/2017 | 1650 nt 3 | nt 47-nt 1636 | 529 aa | 64% with PBV/Mongoose/KNA/M33/2017 | 72% with PBV/Simian/CHN/1R/2009, (ATY68945) |
| PBV/Mongoose/KNA/M58/2017 | 1689 nt 5 | nt 47-nt 1639 | 530 aa | 99% with PBV/Mongoose/KNA/M45/2017 and PBV/Mongoose/KNA/M67/2017 | 77% with PBV/Gorilla/COG/141-RdRp1718/2015, (AVA30701) |
| PBV/Mongoose/KNA/M67/2017 | 1654 nt 3 | nt 47-nt 1639 | 530 aa | 99% with PBV/Mongoose/KNA/M45/2017 and PBV/Mongoose/KNA/M58/2017 | 77% with PBV/Gorilla/COG/141-RdRp1718/2015, (AVA30701) |
| PBV/Mongoose/KNA/M72/2017 | 1691 nt 3 | nt 75-nt 1679 | 534 aa | 65% with PBV/Mongoose/KNA/M33/2017 | 81% with PBV/Chicken/BRA/RS/BR/15/1S-1/2015, (AXL64617) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleymann, A.; Becker, A.A.M.J.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code. Viruses 2020, 12, 99. https://doi.org/10.3390/v12010099
Kleymann A, Becker AAMJ, Malik YS, Kobayashi N, Ghosh S. Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code. Viruses. 2020; 12(1):99. https://doi.org/10.3390/v12010099
Chicago/Turabian StyleKleymann, Alyssa, Anne A. M. J. Becker, Yashpal S. Malik, Nobumichi Kobayashi, and Souvik Ghosh. 2020. "Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code" Viruses 12, no. 1: 99. https://doi.org/10.3390/v12010099