
Abstract
The present study investigates the production and partial biochemical characterization of an extracellular thermostable xylanase from the Bacillus oceanisediminis strain SJ3 newly recovered from Algerian soil using three phase partitioning (TPP). The maximum xylanase activity recorded after 2 days of incubation at 37 °C was 20.24 U/ml in the presence of oat spelt xylan. The results indicated that the enzyme recovered in the middle phase of TPP system using the optimum parameters were determined as 50% ammonium sulfate saturation with 1.0:1.5 ratio of crude extract: t-butanol at pH and temperature of 8.0 and 10 °C, respectively. The xylanase was recovered with 3.48 purification fold and 107% activity recovery. The enzyme was optimally active at pH 7.0 and was stable over a broad pH range of 5.0–10. The optimum temperature for xylanase activity was 55 °C and the half-life time at this temperature was of 6 h. At this time point the enzyme retained 50% of its activity after incubation for 2 h at 95 °C. The crude enzyme resist to sodium dodecyl sulfate and β-mercaptoethanol, while all the tested ions do not affect the activity of the enzyme. The recovered enzyme is, at least, stable in tested organic solvents except in propanol where a reduction of 46.5% was observed. Further, the stability of the xylanase was higher in hydrophobic solvents where a maximum stability was observed with cyclohexane. These properties make this enzyme to be highly thermostable and may be suggested as a potential candidate for application in some industrial processes. To the best of our knowledge, this is the first report of xylanase activity and recoverey using three phase partitioning from B. oceanisediminis.
Similar content being viewed by others

Background
Hemicellulose is the second most abundant renewable biomass after cellulose in nature (Collins et al. 2005). Xylan is the major component of hemicelluloses in wood from angiosperms, where it accounts for 15–30% of the total dry weight. In gymnosperms, however, xylans contribute only 7–12% of the total dry weight. The structure of xylan is complex, and its complete biodegradation requires the concerted action of xylanolytic enzymes (Trajano et al. 2014; Zhang and Viikari 2014). Xylans are heterogeneous polysaccharides with a backbone consisting of β-1,4 linked d-xylosyl residues.
Endo-β-1,4 xylanases (EC 3.2.1.8) are the main enzymes responsible for cleavage of the linkages within the xylan backbone (Collins et al. 2005), to which short side chains of O-acetyl, α-l-arabinofuranosyl, d-α glucuronic, and phenolic acid residues are attached (Collins et al. 2005; Terrasan et al. 2010; Xie et al. 2015). Xylanases have been used in a wide range of industrial applications and processes. They have been applied in the bioconversion of lignocellulosic material and agro-wastes to fermentative products, clarification of juices, improvement in consistency of beer, and the digestibility of animal feed stock (Badhan et al. 2007; Elgharbi et al. 2015a, b; Shameer 2016; Jain and Krishnan 2017). Due to their important activity at alkaline pH (8.0–11) and high temperature (50–90 °C), thermostable alkaline xylanases have attracted special attention in the pulp bio-bleaching industry (Techapun et al. 2003; Bouacem et al. 2014; Boucherba et al. 2014; Bouanane-Darenfed et al. 2016). Xylanase, together with other hydrolytic enzymes, have also proved useful for the generation of bio-fuels, including ethanol, from lignocellulosic biomass. Xylanases are used in pulp pre-bleaching process to remove the hemicelluloses, which bind to the pulp. The hydrolysis of pulp bound hemicelluloses releases the lignin in the pulp, reducing the amount of chlorine required for conventional chemical bleaching and minimizing the toxic, chloroorganic waste. Therefore, xylanases from alkalophilic bacteria and actinomycetes and fungi have been studied widely (Perez-Rodriguez et al. 2014; Wang et al. 2014). However, large scale cultivation of fungi and actinomycetes is often difficult because of their slow generation time, coproduction of highly viscous polymers, and poor oxygen transfer (Wong et al. 1997; Garg et al. 2011). Bacillus genus is used more extensively than other bacteria in industrial fermentations, since they produce most of their enzymes. Some Bacillus strains have been reported as xylanolytic enzymes producers (Lindner et al. 1994; Seo et al. 2013; Tarayre et al. 2013; Elgharbi et al. 2015a; Zouari et al. 2015).
Bacillus oceanisediminis sp. nov. was first isolated from a marine sediment collected in the South Sea of China (Zhang et al. 2010). Considering the above, the present study was undertaken to described, for the first time, the production of a thermostable xylanase from B. oceanisediminis strain SJ3 recently isolated by our laboratory from Algerian soil, an attempt was made to biochemically characterize the xylanase activity secreted by this strain. Also, preliminary investigation using three phase partitioning (TPP) system (Gagaoua et al. 2014; Gagaoua and Hafid 2016) for xylanase purification was performed. In TPP process, firstly an inorganic salt (generally ammonium sulfate) is added to the crude extract containing proteins then mixted with tert-butanol in an appropriate amount (Gagaoua et al. 2015, 2016, 2017). When t-butanol is added in the presence of ammonium sulfate, it pushes the protein out of the solution. In this process t-butanol binds to hydrophobic part of the proteins to reduce the density of the proteins, leading to float above the denser aqueous salt phase. Within approximately an hour, it forms an interfacial (middle) precipitate between the lower aqueous and upper organic phase that usually contains t-butanol (Gagaoua and Hafid 2016).
Methods
Substrates, reagents, and chemicals
Birchwood xylan, oat spelt xylan, starch, carboxymethyl cellulose (CMC, low viscosity), tert-butanol, ammonium sulfate, and 3,5-dinitrosalicylic acid (DNS) were purchased from Sigma Chemical Company (St. Louis, MO, USA). Unless otherwise specified, all other reagents and chemicals were of the analytical grade or highest level of purity available.
Collection of samples and culture conditions of microorganisms
The garden soil samples were collected from Bejaia north east of Algeria (Kabylia region) in March 2015. The soil was collected from five places and samples were pooled. Sub-samples of approximately 1 g were suspended in 100 ml sterile distilled water. Mixtures were allowed to settle and serial dilutions were prepared. From each dilution, 0.1 ml was taken and spread on agar plates of medium containing in g/l oat spelt xylan 10, yeast extract 2, NaCl 2.5, NH4Cl 5, KH2PO4 15, Na2HPO4 30, MgSO4·7H2O 0.25, and bacteriological agar 15. In this medium, there is a little modification of the main carbon source, the oat spelt xylan was used instead the birchwood xylan (Viet et al. 1991). The plates were incubated at pH 7 and 37 °C for 2 days at 250 rpm. Those colonies that grew well under such conditions and showed an orange zone around the colonies after red Congo were retained for second screening. Colonies with a clear zone formation following the hydrolysis of xylan were evaluated as xylanase producers. Several xylanlolytic strains were isolated and SJ3, which exhibited a large clear zone of hydrolysis, was selected and retained for further experimental study.
Bacterial identification of the isolate SJ3
Analytical profile index (API) strip tests and 16S rRNA gene sequencing were carried out for the identification of the genus to which the strain belong.
API 50 CHB/E and the API 20E strips (bioMérieux, SA, Marcy-l’Etoile, France) were used to investigate the physiological and biochemical characteristics of strain SJ3, as recommended elsewhere (Logan and Berkeley 1984). The growth temperature (4, 10, 15, 20, 25, 30, 35, 40, and 45 °C), pH level values (4, 5, 6, 7, 8, 9, 10, 11, and 12) and sodium chloride regimes were determined.
The 16S rRNA gene was amplified by PCR using forward primer F-d1 5′-AGAGTTTGATCCTGGCTCAG-3′, and reverse primer R-d1 5′-AAGGAGGTGATCCAAGCC-3′, designed from base positions 8–27 and 1541–1525, respectively, which were the conserved zones within the rRNA operon of Escherichia coli (Gurtler and Stanisich 1996). The genomic DNA of strain SJ3 was purified using the Wizard® Genomic DNA Purification Kit (Promega, Madison, WI, USA) and then used as a template for PCR amplification (30 cycles, 94 °C for 45 s denaturation, 60 °C for 45 s primer annealing, and 72 °C for 60 s extension). The amplified ~1.5 kb PCR product was cloned in the pGEM-T Easy vector (Promega, Madison, WI, USA), leading to pSJ3-16S plasmid (this study). The E. coli DH5α (F− supE44 Φ80 δlacZ ΔM15 Δ(lacZYA - argF) U169 endA1 recA1 hsdR17 (r − k , m + k ) deoR thi - 1 λ − gyrA96 relA1) (Invitrogen, Carlsbad, CA, USA) was used as a host strain. All recombinant clones of E. coli were grown in Luria–Bertani (LB) broth media with the addition of ampicillin, isopropyl-thio-β- d -galactopyranoside (IPTG), and X-gal for screening. DNA electrophoresis, DNA purification, restriction, ligation, and transformation were all performed according to the method previously described elsewhere (Sambrook et al. 1989).
DNA sequencing and molecular phylogenetic analysis
The nucleotide sequences of the cloned 16S rRNA gene were determined on both strands using BigDye Terminator Cycle Sequencing Ready Reaction kits and the automated DNA sequencer ABI PRISM® 3100-Avant Genetic Analyser (Applied Biosystems, Foster City, CA, USA. The RapidSeq36_POP6 run module was used, and the samples were analyzed using the ABI sequencing analysis software v. 3.7 NT.
The sequences obtained were compared to those present in the public sequence databases and with the EzTaxon-e server (http://eztaxon-e.ezbiocloud.net/), a web-based tool for the identification of prokaryotes based on 16S rRNA gene sequences from type strains (Kim et al. 2012).
Phylogenetic and molecular evolutionary genetic analyses were conducted via the the molecular evolutionary genetics analysis (MEGA) software version 5 (http://www.megasoftware.net). Distances and clustering were calculated using the neighbor-joining method. The tree topology of the neighbor-joining data was evaluated by Bootstrap analysis with 100 re-samplings.
Xylanase assay
Xylanase activity was determined by measuring the release of reducing sugar from soluble xylan using the DNS method (Miller 1959). In brief, 0.9 ml buffer A (10 mg/ml oat spelt xylan in 50 mM sodium-phosphate buffer at pH 7) were mixed with 0.1 ml of the recovered enzyme solution (1 mg/ml). After incubation at 55 °C for 10 min, the reaction was terminated by adding 1.5 ml of the DNS reagent (Maalej et al. 2009). The mixture was then boiled for 5 min and cooled. Absorption was measured at 540 nm.
One unit of xylanase activity was defined as the amount of enzyme that released 1 µmol of reducing sugar equivalent to xylose per min under the assay conditions.
Xylanase production
Gowth condition of the xylanase activity
To study the properties of the xylanase activity production, the isolates having high xylanase activities were cultivated in 250 ml shake-flasks containing 50 ml basic xylanase production medium at 37 °C. The basic xylanase production medium was prepared at pH 7.0 containing oat spelt xylan. The culture was harvested after 48 h, and centrifuged (10,000 rpm for 10 min). Growth was measured by determining absorbance at 600 nm. The sample was then kept at 4 °C in the refrigerator.
Effect of incubation time on xylanase production
Pre-culture (2%) was used to inoculate 250 ml xylan defined medium at 37 °C for 72 h. culture samples were collected each 4 h during the cultivation period. Immediately after collection, the samples were centrifuged at 4 °C and 10,000g for 20 min. Supernatants were analyzed for xylanase activity as described above.
Partial biochemical characterization of the recovered enzyme by TPP
Extraction and partial purification of xylanase by TPP
Aqueous systems such as three phase partitioning (TPP), known as simple, economical and quick methods, were described for the fast recovery of enzymes (Gagaoua and Hafid 2016). This elegant non-chromatographic tool may be performed in a purification process to be used successfully in food or other industries. For its application in this study, the crude extract was first collected after 48 h of batch incubation (Boucherba et al. 2014). The culture supernatant containing secreted xylanases was concentrated using Sartorius membranes (with 10-kDa cutoff membrane; Millipore) after a centrifugation at 10,000 rpm for 10 min. Then, TPP experiments were carried out following the recommendations of Gagaoua et al. (2015). The enzyme exclusively recovered in the interfacial phase was gently separated from the other phases and dissolved in 50 mM Tris–HCl buffer (pH 8.5) and dialyzed overnight at 4–5 °C and used for enzyme characterization.
Effect of temperature and pH on xylanase activity
Optimal temperature was determined by assaying the enzyme activity between 20 and 100 °C, by incubating the enzyme along with the substrate for 10 min at the respective temperature. Relative xylanase activity was determined using 10 mg/ml oat spelt xylan at various pHs. The pH range used varied from 4 to 10. Three different buffers (50 mM) were used. Sodium acetate buffer was used for pH 4–6; Sodium-phosphate buffer was used for pH from 6 to 7 and Tris–HCl buffer for pH 7–10.
Effect of temperature on xylanase stability
The thermostability was determined at temperatures of 50, 55, 60, and 95 °C, after incubation with the substrate for different times (from 0.5 to 7 h); remaining xylanase activity was measured under standard assay conditions. The non-heated enzyme, which was left at room temperature, was considered as control (100%).
Effect of pH on xylanase stability
For pH stability, the enzyme was incubated with different buffers viz. 50 mM acetate buffer for pH range 4–6, 50 mM phosphate buffer for pH range 6–7, and 50 mM Tris–HCl buffer for pH range 7–10 at 55 °C for 1 h. Thereafter, enzyme activity was determined using the enzyme assay as described above.
Effect of metal ions and reagents on activity
The effect of metallic ions at concentration of 5 mM, chelating agents, surfactants, and inhibitors on the activity of crude xylanase were determined by preincubating the enzyme in the presence of Na+, Mg2+, Ca2+, Mn2+, Fe2+, Zn2+, Cu2+, K +, Hg2+, and Cd2+, EDTA (5 Mm), SDS (1%), β-mercaptoethanol (20 mM), and Triton X-100 (1%) for 30 min at 55 °C before adding the substrate (Ozcan et al. 2011). Subsequently, relative xylanase activities were measured at standard enzyme assay conditions. Relative activity was expressed as the percentage of the activity observed in the absence of any compound.
Activity of crude enzyme on various carbohydrate substrate
The presence of other carbohydrase was analyzed using oat spelt xylan, birchwood xylan, starch, and CMC (10 mg/ml). The reducing sugar released during the assay was quantified by spectroscopy at λ 540.
Effect of organic solvents on xylanase activity
Cell free supernatant having maximum xylanase activity was incubated with 30% (v/v) of different organic solvents, namely, acetone, propanol, ethanol, methanol, chloroform, heptane, cyclohexane, and toluene for 30 min at 55 °C. The residual xylanase activity was estimated against the control, in which solvent was not present.
Statistical analysis
All determinations were performed at least in three independent replicates, and the control experiment without xylanase was carried out under the same conditions. The experimental results were expressed as the mean of the replicate determinations and standard deviation (mean ± SD). The statistical significance was evaluated using t tests for two-sample comparison and one-way analysis of variance (ANOVA) followed by Duncan test. The results were considered statistically significant for P values of less than 0.05. The statistical analysis was performed using the R package Version 3.1.1 (Vanderbilt University, USA).
Nucleotide sequence accession number
The data reported in this work for the nucleotide sequence of the 16S rRNA (1089 bp) gene of the isolate SJ3 have been deposited in the DDBJ/EMBL/GenBank databases under Accession Number KT222887.
Results and discussion
Screening of xylanase-producing bacteria from Algerain soil and molecular characterization of the target microorganism
In the current study, ten candidates were obtained from the first screening as xylanase producers. Among them, a bacterium called SJ3, displayed the highest extracellular xylanase activity after 2 days incubation in an initial medium (data not shown) and was, therefore, retained for all subsequent studies.
The physiological and biochemical characteristics of the SJ3 isolate presented in this study were investigated according to well-established protocols and criteria described in the Bergey’s Manual of Systematic Bacteriology as well as the API 50 CHB/E and the API 20E galleries for representative strains. The findings indicated that the SJ3 isolate was Gram-stain-positive, motile, rod-shaped, catalase-positive, aerobic, and endospore forming microorganism. Optimal growth temperature was 37 °C; optimal pH was 7.0. According to the results obtained using the API 50 CHB/E medium and the API 20E strips, the characteristics strongly confirmed that the isolate belongs to Bacillaceae order and Bacillus genus. The physiological and some biochemical properties of the isolate SJ3 are given in Table 1.
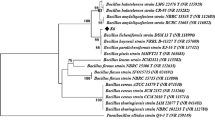
The 16S rRNA gene sequence (KT222887) obtained was submitted to GenBank BLAST search analyses, which yielded a strong homology of up to 99% with those of several cultivated strains of Bacillus. From the analysis of the almost-complete 16S rRNA gene sequence, this strain was found to be similar to B. oceanisediminis strain H T2 (99.16% sequence identity). Through the alignment of homologous nucleotide sequence of known bacteria, phylogenetic relationships could be inferred, and the phylogenetic position of the strain and related strains based on the 16S rDNA sequence is shown in Fig. 1. Taken together, the results suggest that this isolate may be assigned as B. oceanisediminis strain SJ3.

Phylogenetic tree based on 16S rRNA gene sequences showing the position of strain SJ3 within the radiation of the genus Bacillus. The sequence of E. coli strain ATCC 11775T (Accession No. X80725) was chosen arbitrarily as an outgroup. Bar 0.02 nt substitutions per base. Numbers at nodes (>50%) indicate support for the internal branches within the tree obtained by bootstrap analysis (percentages of 100 bootstraps). NCBI accession numbers are presented in parentheses
Optimization of xylanase production by strain SJ3
In the current study, the bacterial strains were newly isolated from Algerain soil samples (Bejaia north east, Algeria), were screened for their xylanase activities. Using the ratio of the clear zone diameter (onto xylan agar plates) and that of the colony, five isolates exhibiting the highest ratio were tested for xylanase production in liquid culture. Among those strains, a bacterium called strain SJ3, displayed the highest extracellular xylanase activity (20.24 U/ml) after 48 h incubation in an optimized medium (Fig. 2) and was, therefore, retained for all subsequent studies.

Time course of Bacillus oceanisediminis strain SJ3 cell growth (open diamond) monitored by measuring the OD at 600 nm and xylanase production (closed diamond). Vertical bars indicate standard error of the mean (n = 3)
Time course of xylanase production showed maximum enzyme activity at 48 h of incubation and thereafter, it remained less constant till 72 h (Fig. 2).
It is the same case with Bacillus subtilis strain ASH (Sanghi et al. 2009). The optimum time resulting in maximum enzyme titre is likely to depend on several factors including the microbial strain. A survey of the literature revealed the highest enzyme production from Bacillus pumilus strain SV-85S after 36 h (Nagar et al. 2010) and Bacillus sp. strain SSP-34 after 96 h (Subramaniyan and Prema 2000) and B. pumilus strain VLK-1 after 56 h of incubation (Kumar et al. 2014). In the above reports, the activity of xylanase exhibited a decline after reaching a maximum value, which might be due to proteolysis of the enzyme. However, in the present study, though the incubation period for xylanase production from B. oceanisediminis strain SJ3 was shorter than some other Bacillus sp. yet it did not decline after attaining the highest level.
Some biochemical properties of the crude enzyme
Xylanase activity from B. oceanisediminis strain SJ3 was efficiently recovered using the TPP technique. A purification fold of 3.48 and a recovery yield of 107% were obtained. Using macroaffinity ligand-facilitated TPP, Sharma and Gupta (2002) purified a xylanase from Aspergillus niger with a recovery yield of 60% and a 95-fold purification. The authors reported other recovery parameters using the denatured xylanase and the optimal parameters were 93% and a purification factor of 21 (Roy et al. 2004). TPP has been reported to recover different enzyme activities (e.g., xylanase, cellulase, cellobiase, β-glucosidase, and α-chymotrypsin) from their inactivated/denatured forms (Roy et al. 2004, 2005; Sardar et al. 2007). These findings suggest that TPP may be a valuable technique for the simultaneous renaturation/purification of the multiple enzymes present in a protein mixture. Concerning the high yield recovery obtained in this preliminary study several studies reported high recovery yields (>100%) for the purification of enzymes using the TPP system (Gagaoua and Hafid 2016; Gagaoua et al. 2017).
Effect of temperature on xylanase activity
The effect of temperature on the xylanase activity from B. oceanisediminis strain SJ3 is shown in Fig. 3a, for 10 min reaction the optimum temperature was 55 °C (assayed in the range 20–100 °C), the xylanase produced by Bacillus brevis is also optimally active at the same temperature (Goswami et al. 2013). The optimum temperature of the enzyme is near to that of the xylanases from B. subtilis strain CXJZ isolated from the degumming line (60 °C) (Guo et al. 2012) and Bacillus sp. strain 41M-1 which showed maximum activity at 50 °C (Nakamura et al. 1995) and Bacillus sp. strain BP-23 (50 °C) (Blanco et al. 1995) but distant from that of the xylanases produced by Bacillus halodurans strain TSEV1 (80 °C) (Kumar and Satyanarayana 2014), Caldicoprobacter algeriensis strain TH7C1T (Bouacem et al. 2014), B. subtilis strain GN156 (40 °C) (Pratumteep et al. 2010), and Bacillus amyloliquefaciens strain CH51 (25 °C) (Baek et al. 2012).

Effects of temperature (a) and pH (b) on xylanase activity produced by Bacillus oceanisediminis strain SJ3 and recovered by three phase partitioning. a The enzyme activity was determined by incubating the enzyme with 10 mg/ml oat spelt xylan dissolved in 50 mM phosphate buffer at pH 7. The activity of the enzyme at 55 °C was taken as 100%. b The enzyme was incubated at 55 °C with 10 mg/ml oat spelt xylan dissolved in different buffer. Buffer solutions used for pH activity are presented in “Results and discussion”. The activity of the enzyme at pH 7.0 was taken as 100%. Each point represents the mean of three independent experiments. Vertical bars indicate standard error of the mean (n = 3)
Effect of pH on xylanase activity
The optimum pH of B. oceanisediminis strain SJ3 xylanase activity (assayed in the range 4–10) is 7 (Fig. 3b). Other xylanases from Bacillus strains so far characterized generally show wide differences in their optimal pH, going from acidic values, such as 4 for the glycosyl hydrolase family 11 xylanase from B. amyloliquefaciens strain CH51 (Baek et al. 2012), 5 for the xylanase activity produced by B. subtilis strain GN156 (Pratumteep et al. 2010), 5.8 for the xylanase from B. subtilis strain CXJZ (Gang et al. 2012), up to 9 in the case of the endoxylanase activity from B. halodurans strain TSEV1 (Kumar and Satyanarayana 2013, 2014).
Thermostability profile of the xylanase activity
Thermal stability was carried out by preincubating xylanase up to 7 h at 50, 55, 60, and 95 °C (Fig. 4), at 50 °C there was no significant decrease in xylanase activity during 4 h. The enzyme was stable at 50 °C, with a half-life time of 9 h, a half-life time of 6 and 4.72 h was respectively observed at 55 and 60 °C. B. brevis xylanase is less thermostable, it showed a half-life time of 3 h at 55 °C (Goswami et al. 2013).

Thermostability profile of Bacillus oceanisediminis strain SJ3 xylanase at pH 7 at different temperatures. (closed diamond): 50 °C, (closed square): 55 °C, (closed triangle): 60 °C, (closed circle): 95 °C. Samples were taken at 1 h interval and relative activity was determined. The activity of the non-heated enzyme was taken to be 100%. Each point represents the mean of three independent experiments. Vertical bars indicate standard error of the mean (n = 3)
At 95 °C the profile obtained for thermostability showed that 50% of the original activity was retained after 2 h exposure, the results clearly indicated that the suitable temperature range for industrial application for xylanase from B. oceanisediminis strain SJ3 was 50–95 °C. This xylanase is more thermostable than B. amyloliquefaciens strain XR44A xylanase activity which showed a half-life time of 5 min at 70 °C, 15 min at both 50 and 60 °C, and 2 h at 40 °C. Interestingly, it retained 90% of activity for at least 2 days at 30 °C, with a half-life time of 7 days. The enzyme immediately loses activity at temperatures higher than 70 °C (Amore et al. 2015), the xylanase produced by Bacillus aerophilus strain KGJ2, retained more than 90% activity after incubation at 80–90 °C for 60 min (Gowdhaman et al. 2014). The enzyme produced by Bacillus sp. strain DM-15 was stable for 15 min at 60 °C while 95% of the original activity was lost at 90 °C (Ozcan et al. 2011).
The xylanase from Pseudomonas macquariensis had half-life time of 2 h at 50 °C whereas it had a half-life time of 1 h at 60 °C. At high temperatures, enzyme gets partly unfolded (Sharma et al. 2008). The xylanase of B. oceanisediminis strain SJ3 is highly thermostable, such enzymes with high thermostability and an ability to function at wide pH range are desirable for many industrial processes which take place at very high or low pH and high temperature. With this respect, the strain could be a good source for industrial and biotechnological applications.
pH stability profile of the xylanase activity
It is observed that the highest xylanase activity was established at pH 7.0; on the other hand, it was found to be most stable at pH 7.0–8.0 but it was also stable in a range of pH 5–10 and at pH 10 approximately 80% of its activity was retained (Fig. 5). The enzyme stable in alkaline conditions were characterized by a decreased number of acidic residues and an increased number of arginines (Hakulinen et al. 2003). The similar pattern of pH stability was also found in Bacillus vallismortis strain RSPP-15 (Gaur et al. 2015).

pH stability of the xylanase activity produced by Bacillus oceanisediminis strain SJ3 and recovered by three phase partitioning. The crude enzyme was incubated with 50 mM buffers at 55 °C for 1 h and relative activity was measured under the standard assay conditions. The activity of the enzyme at optimum pH was taken as 100%. Buffer solutions used for pH stability are presented in “Results and discussion”. Each point represents the mean of three independent experiments. Vertical bars indicate standard error of the mean (n = 3)
Effect of metallic ions, reagents, and inhibitors on xylanase activity
We investigated the effects of metallic ions and other reagents on the activities of the crude xylanase (Table 2). Most of the metallic ions (at concentration of 5 mM) tested had little influence on the activity, the same results were obtained with the xylanases produced by Bacillus sp. strain SPS-0 (Bataillon et al. 2000); in this experiment, maximum xylanase production was reported in the presence of Ca2+ (138%); some other researchers also reported that Ca2+ ion strongly stimulated xylanase activity. Slightly stimulation was also observed by addition of Mg2+ (106%) (Mamo et al. 2006; Lv et al. 2008; Ozcan et al. 2011).
On the other hand, the inhibition of xylanases by calcuim and magnesium ions have also been reported (Hmida-Sayari et al. 2012; Chang et al. 2017). Xylanase was strongly inhibited in the presence of Hg2+. Similar results were observed in case of B. subtilis (Sanghi et al. 2010) and Bacillus halodurans strain PPKS-2 (Prakash et al. 2012), it has been reported that the xylanase activity was inhibited by mercury ion, which might be due to its interaction with sulfhydryl groups of cysteine residue in or close to the active site of the enzyme (Bastawde 1992).
The chelating agent EDTA enveloping metal ions extensively did not change the xylanase activity (Table 2) that means the enzyme did not require metal ions for its catalysis.
Triton X-100 and β-mercaptoethanol had little effect on the xylanase activity (Table 2) whereas the Bacillus DM-15 xylanase is sensitive (Ozcan et al. 2011).
Total inactivation due to SDS has already been reported for xylanases of different origins (Fujimoto et al. 1995), in contrast to the resistance to SDS was found in this study, with 87% relative activity after 10 min at 55 °C (Table 2).
Activity of the crude xylanase on various carbohydrate substrates
Activity of the crude enzyme on some carbohydrate was showed at Fig. 6, the crude enzyme mainly contained xylanase as indicated by the highest activity on birchwood xylan (25 U/ml) and oat spelt xylan (20 U/ml). The crude enzyme did not contain amylase but hardly cellulase (1.99 U/ml). Crude enzymes produced by Bacillus sp. strain AQ1 not only showed xylanolytic activity but also amylolytic and cellulolytic activity (Wahyuntari et al. 2009). Comparisons to the large literature studies as summarized in Table 3.

The effect of the carbohydrate substrate source on the xylanase activity produced by Bacillus oceanisediminis strain SJ3 and recovered by three phase partitioning. The enzyme was incubated with 10 mg/ml of substrate at 55 °C and pH 7.0. Each point represents the mean of three independent experiments. Vertical bars indicate standard error of the mean (n = 3)
Based on the available data from this experiment, the difference in crude enzyme on the different xylan substrate could not be explained yet. It is still needed more complete studies to elaborate the type of xylanolytic activities present in the crude enzyme of B. oceanisediminis strain SJ3. From preliminary study, it can be observed that the strain SJ3 was able to grow and produce xylanases using commercial xylan. The pH and temperature optima of the preparation were 7 and 55 °C, respectively, and the enzyme was stable in a range of pH 5–10 retained 50% of its activity during 6 h at 55 °C. The enzyme is also resistant to hydrophobic solvents, these properties place this enzyme as promising for industrial and biotechnological applications especially lignocellulose bioconversion and bioethanol production.
Effect of organic solvents of the xylanase activity
The xylanase from B. oceanisediminis strain SJ3 is resistant to hydrophobic solvents: heptan, chloroform, toluene, and cyclohexane (the relative activity is 99.2%) but a loss of the enzyme activity was observed by addition of 30% (v/v) of methanol, ethanol, propanol, and acetone (Fig. 7). These alcohols completely inhibited the enzyme from Termitomyces sp. and Macrotermes subhyalinus at 30% (v/v) and 60% (v/v), respectively. Primary alcohols including methanol, ethanol, and isopropanol as well as polyhydric alcohol containing glycol and glycerol, all showed inhibitory effects on A. niger strain C3486 xylanase activity which retained around 90% at the concentration of 2% (v/v) and less than 60% of its initial activity at 30% (Yang et al. 2010).

Effect of organic solvents on xylanase activity produced by Bacillus oceanisediminis strain SJ3 and recovered by three phase partitioning. Relative xylanase activity was expressed as a percentage of the control reaction without solvent. Each point represents the mean of three independent experiments. Vertical bars indicate standard error of the mean (n = 3)
In some cases, the presence of solvents enhanced the xylanase activity, for example the xylanase of B. vallismortis is extraordinarily stable in the presence of all organic solvents under study. After incubation with n-dodecane, isooctane, n-decane, xylene, toluene, n-hexane, n-butanol, and cyclohexane, the xylanase activity increased to 230.8, 137.7, 219.8, 107, 190.5, 194.7, 179.3, and 111.6%, respectively (Gaur and Tiwari 2015).
Conclusion
In conclusion, a new extracellular thermostable xylanase from B. oceanisediminis strain SJ3 was produced and characterized in this study. The preliminary results of the use of Three phase partitioning for the recovery of the xylanase were presented. The time course for xylanase accumulation by strain SJ3 in xylan-based medium showed that the highest xylanase activity reached 20.24 U/ml in an optimized medium with oats spelt xylan used as a substrate after 48 h of cultivation. The crude xylanase from strain SJ3 was biochemically characterized. The results revealed that the enzyme was highly stable and active at high temperature (55 °C) and alkaline pH 7.0. Properties of this enzyme such as high specific activity, wide range of pH optimum and stability, and thermostability at elevated temperature as well as organic solvents tolerance, are appropriate for industrial and biotechnological applications. Interestingly, this enzyme presented high xylanolytic activity with oats spelt xylan, and was very effective in the pulp bleaching industry, thus offering a potential promising candidate for application in biotechnological bioprocesses. Accordingly, further studies, some of which are currently underway, are needed to investigate the purification to homogeneity and encoding gene, perform site-directed mutagenesis, and determine its structure–function relationships.
References
Amore A, Parameswaran B, Kumar R, Birolo L, Vinciguerra R, Marcolongo L, Ionata E, La Cara F, Pandey A, Faraco V (2015) Application of a new xylanase activity from Bacillus amyloliquefaciens XR44A in brewer’s spent grain saccharification. J Chem Technol Biotechnol 90:573–581
Badhan AK, Chadha BS, Kaur J, Saini HS, Bhat MK (2007) Production of multiple xylanolytic and cellulolytic enzymes by thermophilic fungus Myceliophthora sp. IMI 387099. Bioresour Technol 98:504–510
Baek CU, Lee SG, Chung YR, Cho I, Kim JH (2012) Cloning of a family 11 xylanase gene from Bacillus amyloliquefaciens CH51 isolated from Cheonggukjang. Indian J Microbiol 52:695–700
Bastawde KB (1992) Xylan structure, microbial xylanases, and their mode of action. World J Microbiol Biotechnol 8:353–368
Bataillon M, Nunes Cardinali A, Castillon N, Duchiron F (2000) Purification and characterization of a moderately thermostable xylanase from Bacillus sp. strain SPS-0. Enzyme Microb Technol 26:187–192
Blanco A, Vidal T, Colom JF, Pastor FI (1995) Purification and properties of xylanase A from alkali-tolerant Bacillus sp. strain BP-23. Appl Environ Microbiol 61:4468–4470
Bouacem K, Bouanane-Darenfed A, Boucherba N, Joseph M, Gagaoua M, Ben Hania W, Kecha M, Benallaoua S, Hacene H, Ollivier B, Fardeau ML (2014) Partial characterization of xylanase produced by Caldicoprobacter algeriensis, a new thermophilic anaerobic bacterium isolated from an Algerian hot spring. Appl Biochem Biotechnol 174:1969–1981
Bouanane-Darenfed A, Boucherba N, Bouacem K, Gagaoua M, Joseph M, Kebbouche-Gana S, Nateche F, Hacene H, Ollivier B, Cayol J-L, Fardeau M-L (2016) Characterization of a purified thermostable xylanase from Caldicoprobacter algeriensis sp. nov. strain TH7C1T. Carbohyd Res 419:60–68
Boucherba N, Gagaoua M, Copinet E, Bettache A, Duchiron F, Benallaoua S (2014) Purification and characterization of the xylanase produced by Jonesia denitrificans BN-13. Appl Biochem Biotechnol 172:2694–2705
Chang S, Guo Y, Wu B, He B (2017) Extracellular expression of alkali tolerant xylanase from Bacillus subtilis Lucky9 in E. coli and application for xylooligosaccharides production from agro-industrial waste. Int J Biol Macromol 96:249–256
Collins T, Gerday C, Feller G (2005) Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol Rev 29:3–23
Dheeran P, Nandhagopal N, Kumar S, Jaiswal YK, Adhikari DK (2012) A novel thermostable xylanase of Paenibacillus macerans IIPSP3 isolated from the termite gut. J Ind Microbiol Biotechnol 39(6):851–860
Elgharbi F, Hlima HB, Farhat-Khemakhem A, Ayadi-Zouari D, Bejar S, Hmida-Sayari A (2015a) Expression of A. niger US368 xylanase in E. coli: purification, characterization and copper activation. Int J Biol Macromol 74:263–270
Elgharbi F, Hmida-Sayari A, Zaafouri Y, Bejar S (2015b) Expression of an Aspergillus niger xylanase in yeast: application in breadmaking and in vitro digestion. Int J Biol Macromol 79:103–109
Ellis JT, Magnuson TS (2012) Thermostable and alkalistable xylanases produced by the thermophilic bacterium Anoxybacillus flavithermus TWXYL3. ISRN microbiology
Fujimoto H, Ooi T, Wang S-L, Takizawa T, Hidaka H, Murao S, Arai M (1995) Purification and properties of three xylanases from Aspergillus aculeatus. Biosci Biotechnol Biochem 59:538–540
Gagaoua M, Hafid K (2016) Three phase partitioning system, an emerging non-chromatographic tool for proteolytic enzymes recovery and purification. Biosens J 5(1):100134
Gagaoua M, Boucherba N, Bouanane-Darenfed A, Ziane F, Nait-Rabah S, Hafid K, Boudechicha HR (2014) Three-phase partitioning as an efficient method for the purification and recovery of ficin from Mediterranean fig (Ficus carica L.) latex. Sep Purif Technol 132:461–467
Gagaoua M, Hoggas N, Hafid K (2015) Three phase partitioning of zingibain, a milk-clotting enzyme from Zingiber officinale Roscoe rhizomes. Int J Biol Macromol 73:245–252
Gagaoua M, Hafid K, Hoggas N (2016) Data in support of three phase partitioning of zingibain, a milk-clotting enzyme from Zingiber officinale Roscoe rhizomes. Data in brief 6:634–639
Gagaoua M, Ziane F, Nait Rabah S, Boucherba N, El-Hadef El-Okki Ait Kaki A, Bouanane-Darenfed A, Hafid K (2017) Three phase partitioning, a scalable method for the purification and recovery of cucumisin, a milk-clotting enzyme, from the juice of Cucumis melo var. reticulatus. Int J Biol Macromol 102:515–525
Gang GJ, Zbijewski W, Webster Stayman J, Siewerdsen JH (2012) Cascaded systems analysis of noise and detectability in dual-energy cone-beam CT. Med Phys 39:5145–5156
Garg G, Dhiman SS, Mahajan R, Kaur A, Sharma J (2011) Bleach-boosting effect of crude xylanase from Bacillus stearothermophilus SDX on wheat straw pulp. N Biotechnol 28:58–64
Gaur R, Tiwari S (2015) Isolation, production, purification and characterization of an organic-solvent-thermostable alkalophilic cellulase from Bacillus vallismortis RG-07. BMC Biotechnol 15:19
Gaur R, Tiwari S, Rai P, Srivastava V (2015) Isolation, production, and characterization of thermotolerant xylanase from solvent tolerant Bacillus vallismortis RSPP-15. Int J Polym Sci 2015:10
Goswami GK, Pathak RR, Krishnamohan M, Ramesh B (2013) Production, partial purification and biochemical characterization of thermostable xylanase from Bacillus brevis. Biomed Pharmacol J 6:435–440
Goswami GK, Krishnamohan M, Nain V, Aggarwal C, Ramesh B (2014) Cloning and heterologous expression of cellulose free thermostable xylanase from Bacillus brevis. SpringerPlus 3(1):20
Gowdhaman D, Manaswini VS, Jayanthi V, Dhanasri M, Jeyalakshmi G, Gunasekar V, Sugumaran KR, Ponnusami V (2014) Xylanase production from Bacillus aerophilus KGJ2 and its application in xylooligosaccharides preparation. Int J Biol Macromol 64:90–98
Guo G, Liu Z, Xu J, Liu J, Dai X, Xie D, Peng K, Feng X, Duan S, Zheng K, Cheng L, Fu Y (2012) Purification and characterization of a xylanase from Bacillus subtilis isolated from the degumming line. J Basic Microbiol 52:419–428
Gurtler V, Stanisich VA (1996) New approaches to typing and identification of bacteria using the 16S-23S rDNA spacer region. Micobiology 142:3–16
Hakulinen N, Turunen O, Janis J, Leisola M, Rouvinen J (2003) Three-dimensional structures of thermophilic beta-1,4-xylanases from Chaetomium thermophilum and Nonomuraea flexuosa. Comparison of twelve xylanases in relation to their thermal stability. Eur J Biochem 270:1399–1412
Hmida-Sayari A, Taktek S, Elgharbi F, Bejar S (2012) Biochemical characterization, cloning and molecular modeling of a detergent and organic solvent-stable family 11 xylanase from the newly isolated Aspergillus niger US368 strain. Process Biochem 47:1839–1847
Irfan M, Nadeem M, Syed Q, Baig S (2012) Effect of medium composition on xylanase production by Bacillus subtilis using various agricultural wastes. Am Eurasian J Agric Environ Sci 12:561–565
Jain A, Krishnan KP (2017) A glimpse of the diversity of complex polysaccharide-degrading culturable bacteria from Kongsfjorden, Arctic Ocean. Ann Microbiol 2:203–214
Kim OS, Cho YJ, Lee K, Yoon SH, Kim M, Na H, Park SC, Jeon YS, Lee JH, Yi H, Won S, Chun J (2012) Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int J Syst Evol Microbiol 62:716–721
Kumar V, Satyanarayana T (2013) Biochemical and thermodynamic characteristics of thermo-alkali-stable xylanase from a novel polyextremophilic Bacillus halodurans TSEV1. Extremophiles 17:797–808
Kumar V, Satyanarayana T (2014) Production of thermo-alkali-stable xylanase by a novel polyextremophilic Bacillus halodurans TSEV1 in cane molasses medium and its applicability in making whole wheat bread. Bioprocess Biosyst Eng 37:1043–1053
Kumar D, Verma R, Sharma P, Rana A, Sharma R, Prakash C, Bhalla TC (2010) Production and partial purification of xylanase from a new thermophilic isolate. In Biol Forum Int J 2:83–87
Kumar L, Nagar S, Mittal A, Garg N, Gupta VK (2014) Immobilization of xylanase purified from Bacillus pumilus VLK-1 and its application in enrichment of orange and grape juices. J Food Sci Technol 51:1737–1749
Lindner C, Stulke J, Hecker M (1994) Regulation of xylanolytic enzymes in Bacillus subtilis. Microbiology 140(Pt 4):753–757
Logan NA, Berkeley RC (1984) Identification of Bacillus strains using the API system. J Gen Microbiol 130:1871–1882
Lv Z, Yang J, Yuan H (2008) Production, purification and characterization of an alkaliphilic endo-β-1,4-xylanase from a microbial community EMSD5. Enzyme Microb Technol 43:343–348
Maalej I, Belhaj I, Masmoudi NF, Belghith H (2009) Highly thermostable xylanase of the thermophilic fungus Talaromyces thermophilus: purification and characterization. Appl Biochem Biotechnol 158:200–212
Mamo G, Hatti-Kaul R, Mattiasson B (2006) A thermostable alkaline active endo-β-1-4-xylanase from Bacillus halodurans S7: purification and characterization. Enzyme Microb Technol 39:1492–1498
Miller GL (1959) Use of dinitrosalycilic acid reagent for determination of reducing sugars. Anal Chem 31:426–428
Nagar S, Gupta VK, Kumar D, Kumar L, Kuhad RC (2010) Production and optimization of cellulase-free, alkali-stable xylanase by Bacillus pumilus SV-85S in submerged fermentation. J Ind Microbiol Biotechnol 37:71–83
Nagar S, Mittal A, Kumar D, Gupta VK (2012) Production of alkali tolerant cellulase free xylanase in high levels by Bacillus pumilus SV-205. Int J Biol Macromol 50(2):414–420
Nakamura S, Nakai R, Namba K, Kubo T, Wakabayashi K, Aono R, Horikoshi K (1995) Structure-function relationship of the xylanase from alkaliphilic Bacillus sp. strain 41M-1. Nucleic Acids Symp Ser 34:99–100
Ozcan BD, Coskun A, Ozcan N, Baylan M (2011) Some properties of a new thermostable xylanase from alkaliphilic and thermophilic Bacillus sp.isolate DM-15. J Anim Vet Adv 10:138–143
Perez-Rodriguez N, Oliveira F, Perez-Bibbins B, Belo I, Torrado Agrasar A, Dominguez JM (2014) Optimization of xylanase production by filamentous fungi in solid-state fermentation and scale-up to horizontal tube bioreactor. Appl Biochem Biotechnol 173:803–825
Poosarla VG, Chandra TS (2014) Purification and characterization of novel halo-acid-alkali-thermo-stable xylanase from Gracilibacillus sp. TSCPVG. Appl Biochem Biotechnol 173(6):1375–1390
Prakash P, Jayalakshmi SK, Prakash B, Rubul M, Sreeramulu K (2012) Production of alkaliphilic, halotolerent, thermostable cellulase free xylanase by Bacillus halodurans PPKS-2 using agro waste: single step purification and characterization. World J Microbiol Biotechnol 28:183–192
Pratumteep A, Sansernsuk J, Nitisinprasert S, Apiraksakorn J (2010) Production, characterization and hydrolysation products of xylanase from Bacillus subtilis GN156. KKU Res J 15:343–350
Roy I, Sharma A, Gupta MN (2004) Three phase partitioning for simultaneous renaturation and partial purification of Aspergillus niger xylanase. BBA Proteins Proteom 1698:107–110
Roy I, Sharma A, Gupta MN (2005) Recovery of biological activity in reversibly inactivated proteins by three phase partitioning. Enzyme Microb Technol 37:113–120
Sambrook J, Fritsch E, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Sanghi A, Garg N, Kuhar K, Kuhad RC, Gupta VK (2009) Enhanced production of cellulase-free xylanase by alkalophilic Bacillus subtilis ASH and its application in biobleaching of kraft pulp. BioResources 4:1109–1129
Sanghi A, Garg N, Gupta VK, Mittal A, Kuhad RC (2010) One-step purification and characterization of cellulase-free xylanase produced by alkalophilic Bacillus subtilis ash. Braz J Microbiol 41:467–476
Sardar M, Sharma A, Gupta MN (2007) Refolding of a denatured α-chymotrypsin and its smart bioconjugate by three-phase partitioning. Biocatal Biotransform 25:92–97
Seo JK, Park TS, Kwon IH, Piao MY, Lee CH, Ha JK (2013) Characterization of cellulolytic and xylanolytic enzymes of Bacillus licheniformis JK7 isolated from the rumen of a native Korean goat. Asian Australas J Anim Sci 26:50–58
Shameer S (2016) Haloalkaliphilic Bacillus species from solar salterns: an ideal prokaryote for bioprospecting studies. Ann Microbiol 66:1315–1327
Sharma A, Gupta MN (2002) Macroaffinity ligand-facilitated three-phase partitioning (MLFTPP) for purification of xylanase. Biotechnol Bioeng 80:228–232
Sharma M, Chadha BS, Kaur M, Ghatora SK, Saini HS (2008) Molecular characterization of multiple xylanase producing thermophilic/thermotolerant fungi isolated from composting materials. Lett Appl Microbiol 46:526–535
Subramaniyan S (2012) Isolation, purification and characterisation of low molecular weight xylanase from Bacillus pumilus SSP-34. Appl Biochem Biotechnol 166(7):1831–1842
Subramaniyan S, Prema P (2000) Cellulase-free xylanases from Bacillus and other microorganisms. FEMS Microbiol Lett 183:1–7
Tarayre C, Brognaux A, Brasseur C, Bauwens J, Millet C, Matteotti C, Destain J, Vandenbol M, Portetelle D, De Pauw E, Haubruge E, Francis F, Thonart P (2013) Isolation and cultivation of a xylanolytic Bacillus subtilis extracted from the gut of the termite Reticulitermes santonensis. Appl Biochem Biotechnol 171:225–245
Techapun C, Poosaran N, Watanabe M, Sasaki K (2003) Optimization of aeration and agitation rates to improve cellulase-free xylanase production by thermotolerant Streptomyces sp. Ab106 and repeated fed-batch cultivation using agricultural waste. J Biosci Bioeng 95:298–301
Terrasan CR, Temer B, Duarte MC, Carmona EC (2010) Production of xylanolytic enzymes by Penicillium janczewskii. Bioresour Technol 101:4139–4143
Trajano HL, Pattathil S, Tomkins BA, Tschaplinski TJ, Hahn MG, Van Berkel GJ, Wyman CE (2014) Xylan hydrolysis in Populus trichocarpa × P. deltoides and model substrates during hydrothermal pretreatment. Bioresour Technol 179C:202–210
Viet DN, Kamio Y, Abe N, Kaneko J, Izaki K (1991) Purification and properties of beta-1, 4-xylanase from Aeromonas caviae W-61. Appl Environ Microbiol 57:445–449
Wahyuntari B, Mubarik NR, Setyahadi S (2009) Effect of pH, temperature and medium composition on xylanase production by Bacillus sp. AQ-1 and partial characterization of the crude enzyme. Microbiology 3:17–22
Wang W, Wang Z, Cheng B, Zhang J, Li C, Liu X, Yang C (2014) High secretory production of an alkaliphilic actinomycete xylanase and functional roles of some important residues. World J Microbiol Biotechnol 30:2053–2062
Wong KK, Martin LA, Gama FM, Saddler JN, de Jong E (1997) Bleach boosting and direct brightening by multiple xylanase treatments during peroxide bleaching of kraft pulps. Biotechnol Bioeng 54:312–318
Xie Z, Lin W, Luo J (2015) Genome sequence of Cellvibrio pealriver PR1, a xylanolytic and agarolytic bacterium isolated from freshwater. J Biotechnol 214:57–58
Yang YL, Zhang W, Huang JD, Lin L, Lian HX, Lu YP, Wu JD, Wang SH (2010) Purification and characterization of an extracellular xylanase from Aspergillus niger C3486. Afr J Microbiol Res 4:2249–2256
Zhang J, Viikari L (2014) Impact of xylan on synergistic effects of xylanases and cellulases in enzymatic hydrolysis of lignocelluloses. Appl Biochem Biotechnol 174:1393–1402
Zhang J, Wang J, Fang C, Song F, Xin Y, Qu L, Ding K (2010) Bacillus oceanisediminis sp. nov., isolated from marine sediment. Int J Syst Evol Microbiol 60:2924–2929
Zheng HC, Sun MZ, Meng LC, Pei HS, Zhang XQ, Yan Z, Sun JS (2014) Purification and characterization of a thermostable xylanase from Paenibacillus sp. NF1 and its application in xylooligosaccharides production. J Microbiol Biotechnol 24:489–496
Zouari Ayadi D, Hmida Sayari A, Ben Hlima H, Ben Mabrouk S, Mezghani M, Bejar S (2015) Improvement of Trichoderma reesei xylanase II thermal stability by serine to threonine surface mutations. Int J Biol Macromol 72:163–170
Authors’ contributions
NB, MG, and SB designed this research plan and discussed with ABD. NB, MG, CB, KB, MYC performed all the research experiments and NB and MG wrote the draft paper. ABD, YM, and MG helped in the research experiments. MG conducted Three Phase Partitioning experiments. All authors have participated in the interpretation of the results during preparation of the manuscript. All authors read and approved the final manuscript.
Acknowledgements
This work was funded by the Algerian Ministry of Higher Education and Scientific Research under the National Research Program Project (Grant Number F00620110001).
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Boucherba, N., Gagaoua, M., Bouanane-Darenfed, A. et al. Biochemical properties of a new thermo- and solvent-stable xylanase recovered using three phase partitioning from the extract of Bacillus oceanisediminis strain SJ3. Bioresour. Bioprocess. 4, 29 (2017). https://doi.org/10.1186/s40643-017-0161-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-017-0161-9