
Abstract
Insect–plant interactions may be unintentionally affected when introducing genetically modified (GM) crops into an agro-ecosystem. Our aim was to test the non-target effects of a late blight-resistant GM potato on Myzus persicae in greenhouse and climate room experiments and understand how position and number of R gene insertions can affect non-targets in GM events. We also aimed to compare results to baseline differences among three conventional potato varieties varying in resistance to late blight. Aphid development and survival were affected by some GM events in the first generation, though effects disappeared in the second generation. Effects were not dependent on the presence of a marker gene or the insertion of a second resistance gene. Positional effects of gene insertion influenced aphid performance on certain GM events. However, aphid fitness varied considerably more between conventional potato varieties than between Désirée and the GM events. Comparing different GM events to the non-transformed variety is relevant, since unintended effects of insertion can occur. Our protocols can be recommended for in planta risk assessments with aphids. Ecological perspective is gained by selecting several measured endpoints and by comparing the results with a baseline of conventional cultivars.
Similar content being viewed by others


Key message
-
We investigated the hypothesis that characteristics of a GM potato may influence the non-target aphid Myzus persicae.
-
Aphid performance was affected by the resistance gene position but not by the number of resistance genes or the presence of an antibiotic resistance marker gene.
-
Aphid performance varied considerably more between conventional cultivars than between the unmodified and the GM potato.
-
These findings support future protocols for risk assessments of GM crops on non-target insects.
Introduction
To be considered for cultivation in agriculture, genetically modified (GM) crops must be subject to environmental risk assessment (ERA). The biodiversity and ecology of organisms in the agro-ecosystem are considered important in ERA. Plants are the primary producers supporting the trophic webs of agro-ecosystems, and the direct and indirect consequences of introducing genetically modified crops are therefore a relevant concern (Arpaia 2010; EFSA 2010). Risk assessments should be done in several stages or tiers, starting with experiments that have a high likelihood of detecting effects on non-targets to more complex and realistic field conditions (Andow and Hilbeck 2004; Andow and Zwahlen 2006; Houshyani 2012; Kos et al. 2009; Romeis et al. 2011). Each consecutive tier in the ERA should use the feedback acquired in previous steps. Trials in confined conditions are important in early tiers of ERA to establish whether direct effects occur on the life history of particularly important members of the agro-ecosystem or representatives of important functional groups (Andow et al. 2013; Birch et al. 2007; Houshyani 2012; Romeis et al. 2011, 2013).
Before the introduction of GM plants into the ecosystem, testing for non-target effects of a GM crop in the greenhouse first requires a thorough and transparent selection of appropriate non-target organisms (NTOs) (Carstens et al. 2014; EFSA 2010). These tests should be reproducible and reliable and are an important step in the ERA process. A selection procedure of relevant functional groups and endpoints to test must also be included in the ERA. In this study, we based the selection on the protocol outlined in the EFSA guidance document on ERA of GM plants (EFSA 2010) as well as on several other sources (Andow et al. 2013; Gillund et al. 2013; Romeis et al. 2013, 2014; Scholte and Dicke 2005). We selected the aphid Myzus persicae Sulzer to test in planta the non-target effects of a genetically modified potato expressing resistance to late blight.
Most conventional potato cultivars are susceptible to late blight which is caused by the widespread pathogen Phytophthora infestans (Mont.) de Bary, a hemibiotrophic oomycete which colonizes potato leaves, stems and tubers. Genetic modification of the cultivar Désirée conferred resistance to P. infestans through the insertion of one or two resistance genes (R genes) from crossable potato (Solanum tuberosum L.) relatives, Solanum venturii Hawkes & Hjert., (Rpi-vnt1), and Solanum stoloniferum Schltdl & Bouché (Rpi-sto1) (Haesaert et al. 2015; Haverkort et al. 2016). R genes code for receptor proteins which recognize distinct pathogen effectors (in this case from P. infestans). This recognition initiates signal transduction cascades leading to callose deposits and cell death in infected and surrounding cells preventing the pathogen from further spread, which is macroscopically visible as a hypersensitivity response (HR) (Kamoun et al. 1999; Vleeshouwers et al. 2000, 2011).
Late blight R genes can be co-inserted with a selectable marker gene from a bacterium coding for resistance to an antibiotic (transgenesis) or using a marker-free transformation protocol. Because the R genes used in this study are derived from crossable species and the transformation events contain no ‘foreign’ DNA, the latter protocol is referred to as cisgenesis. We tested two transgenic and two cisgenic events containing the same single R gene (Rpi-vnt1). Also we tested two transgenic events harbouring two R genes (Rpi-vnt1 and Rpi-sto1). The location of the R gene insertion in the genome may have an impact on other plant functions and indirectly on non-target aphids. By testing two transformation events of each construct, position effects could be assessed. We also assessed the reproducibility of the experimental protocol by performing the assays on the same plant clones in two laboratories each maintaining their own M. persicae colonies.
In order to compare the magnitude of the effects of these modifications with the variation among commercially available conventional potato varieties, we compared a cisgenic event (also used in concurrent field experiments) with four conventional varieties (including Désirée) varying in their susceptibility to P. infestans (Table 1).
Selection of non-target species Myzus persicae for in planta testing
Many species may be exposed to GM plants in any agro-ecosystem. Since not all species can be tested, a representative subset of NTOs should be selected for consideration in the risk assessment of each GM plant. The GMO Panel of the European Food Safety Authority (EFSA) proposes a species selection approach (EFSA 2010). M. persicae Sulzer (Hemiptera: Aphididae) was chosen based on a final ranking using the aforementioned approach, which includes several important factors. First, it is listed as the most collected phloem feeder in the EFSA arthropod database (Riedel et al. 2016) and second most collected species on potato giving it high relevance as a focal NTO. Second, the species is amenable for rearing in many laboratories, which allows for the measurement of survival and intrinsic rate of increase, which can be used to estimate the population dynamics of this pest.
Aphids are the most important insect pests of potato (Meissle et al. 2012; Radcliffe 1982), and the polyphagous M. persicae is the most prevalent and studied among those. Aphids cause direct damage through piercing and sucking from the plant’s phloem. More problematic is the fact that M. persicae is a vector of over one hundred plant viruses, with about twelve directly affecting potato crops, including several leaf-roll viruses (Kennedy et al. 1962; Ng and Perry 2004; Van Emden et al. 1969). Aphids are a major prey species host many parasitoids (Müller et al. 1999) and are prey to predators such as larval syrphid flies (Raj 1989), ladybugs (Majerus 1994), lacewings, spiders and others (Van Emden et al. 1969). Despite the specificity of an R gene for resistance against P. infestans, it is nevertheless important to understand whether the modification can affect the behaviour or performance of an important NTO like M. persicae (Han et al. 2016) and its population dynamics.
Experimental procedures
Plant material
The GM events tested in this study were developed by the Laboratory of Plant Breeding of Wageningen University and Research (Haesaert et al. 2015; Haverkort et al. 2016). They have been created using Agrobacterium tumefaciens-mediated transfer of the native Rpi-vnt1 gene, from Solanum venturii, using marker-assisted (events A13-13, 17) and marker-free transformation methods (events A15-31, 45, 84). Also, two marker-assisted transformation events (A16-02 and A16-24) were used that were generated using a single T-DNA harbouring the native Rpi-vnt1 and Rpi-sto1 (from Solanum stoloniferum) genes. The tested conventional cultivars and GM events (defined here as clones with gene insertions conferring resistance to the target P. infestans) are described in Table 1. Events were selected as apparently ‘true to type’ as they were morphologically indistinguishable from non-transformed Désirée under tuber-sown field conditions (Haverkort et al. 2016).
All GM events and conventional cultivars were maintained in vitro, on agar medium (purified agar 0.8% + 2.2 g/L Murashige & Skoog + Duchefa 4.4 g/L + saccharose 20 g/L + micro-agar 8 g/L; pH 5.8) in sterile containers. Containers were kept in a climate room at 16:8 light/dark conditions, 21 °C during light hours and 15 °C when dark, and 70% relative humidity. Cuttings were transplanted five weeks before the experiments to allow for root growth, seedlings then transplanted to larger pots and allowed to grow for five weeks before being used in experiments.
Aphid rearing and experimental set-up
WUR
Myzus persicae were collected in 2004 from Wageningen, The Netherlands (51°59′11.5″N 5°39′48.4″E), and reared at the Laboratory of Entomology, Wageningen University and Research (WUR). They were originally kept on radish but maintained for several generations on S. tuberosum cultivar Désirée before experiments began under the same climate room conditions described above.
ENEA
The colony was started from a laboratory strain originally reared at the University of Bologna. The strain was maintained on S. tuberosum cultivar Désirée for several generations before experiments began. The M. persicae colony was maintained under 16:8 light/dark conditions, 24 °C during light hours and 18 °C when dark, and 70% relative humidity.
Testing the GM potato events and conventional potato varieties
First we tested the intrinsic rate of increase and survival of aphids between the non-transformed Désirée and the following GM (from Désirée) events: A15-31, A15-45 (both cisgenic), A13-13, A13-17 (both transgenic), A16-02 and A16-24 (both transgenic with two R genes); all events are described in Table 1. Then, to test reproducibility, WUR and ENEA performed similar experiments comparing specifically the cisgenic events A15-31 and A15-45 to the non-transformed Désirée. Lastly, we compared several conventional potato cultivars: Désirée, Bintje, Première and Sarpo Mira (described in Table 1) with the same measured endpoints as for the aforementioned experiments.
One-day-old aphid nymphs were used in each experiment. Aphid nymphs were placed singly in clip cages (25 mm diameter; 10 mm high) on the abaxial surface of two (at ENEA) or three leaves (WUR) on each plant. Ten (at WUR) to fifteen (at ENEA) plant replicates of each event and the non-transformed Désirée cultivar were used and randomly distributed in the climate room. Due to space limitations, this was split into two or three rounds, each round testing five plants from each event and non-transformed Désirée.
We monitored the fitness of M. persicae for two generations. Aphids were checked every day for mortality and for offspring production; neonate nymphs were counted and removed daily. At WUR, once the first generation produced its first nymphs, one of these was caged on another leaf of the same plant; at ENEA second generations were transferred to a new plant. The parameters collected were: pre-reproductive period and total fecundity, for calculation of intrinsic rate of increase (R m) and aphid mortality of both generations. Intrinsic rate of increase was calculated as described in Wyatt and White (1977): R m = 0.74 (ln Md)/d, where Md is the effective fecundity and d the length of the pre-reproductive period. The means for all aphid parameters used to calculate survival and intrinsic rate of increase are documented in Appendix of Tables 2, 3 and 4.
The same methodology was applied to a second experiment in a greenhouse comparing the first generation of aphid life-history parameters on one cisgenic event (A15-31, highly resistant) and four conventional cultivars varying in their foliar resistance to P. infestans. Cultivar Bintje has a resistance rating of low to very low, cultivar Première and Désirée rate low to medium and Sarpo Mira rates highly resistant to P. infestans (ECPD 2015).
Statistical analysis
Based on a preliminary small-scale experiment (15 individuals), we conducted a prospective power analysis. The measurement endpoint selected was the length of the pre-reproductive period for which a standard deviation of 1.7 days was found. We added a safety margin and set the standard deviation in the power analysis to be 1.9 days. The common within-group standard deviation was set at 2.5, based on the variability registered in the actual experiment. This effect was selected as the smallest relevant effect. The criterion for significance (alpha) was set at 0.050 for a two-tailed test. The analysis was conducted using the Power and Precision 2.1 software (Borenstein et al. 2001). The results indicated a sample size of 28 individuals for each group, and the study will have power of 81.3% to yield a statistically significant result for the differences indicated.
Intrinsic rate of increase was tested with a mixed linear model or generalized linear mixed model when data did not meet the assumptions of normality, with fixed factors being ‘potato event’ and ‘aphid generation’ and random factors including the ‘plant or pot number’ (since there were three clip cages per plant), nested within ‘round’ (experiment was replicated in two rounds). The model was chosen by backwards selection comparing AIC values of simpler models (Burnham et al. 2010). The fixed factor ‘aphid generation’ (first or second generation) proved to have an influence on aphid intrinsic rates of increase (p = 0.0034). For some events, there was an interaction effect between ‘generation’ and the ‘potato event’. For this reason, we separated the two aphid generations and used separate models for each using the same random factors as above. Analysis for comparisons to baseline cultivars was done in a similar way as above, though the experiment was conducted in one round, for one aphid generation, and the only random effect included in the model was ‘plant or pot number’. Analyses for intrinsic rates of increase were conducted using R Statistical Software (R Core Team 2014), with the ‘nlme’ package.
Survival analyses were conducted using a Cox proportional hazards regression model. This was also separated by generation, which played an important role in aphid survival (p = 0.0005) and interacted with the fixed effect of ‘potato event’. This model included the same nested random effects as above and was performed using R Statistical Software (R Development Core Team 2014), with the ‘survival’ package.
Results
Désirée compared to GM events
Comparison of events
In the first generation, aphid intrinsic rate of increase was generally higher on all GM events than on the non-transformed Désirée plants, though the only events significantly differing from Désirée were the transgenic event A13-17 (p = 0.0122) and the cisgenic event A15-31 (p = 0.0198; Fig. 1a). The trend of higher intrinsic rate of increase was no longer observed in the second generation, the events no longer differed from non-transformed Désirée (Figs. 1b, 2).

Mean aphid intrinsic rate of increase (±SE) on Solanum tuberosum isogenic cultivar Désirée, compared to several genetically modified events for two aphid generations. Two events of cisgenic, transgenic and stacked transgenic potatoes were compared. Asterisk (*) indicates significant differences from the isogenic cultivar within the generation

Mean aphid intrinsic rate of increase (±SE) on Solanum tuberosum isogenic cultivar Désirée, compared to genetically modified events A15-31 and A15-45, for two aphid generations in a at WUR Laboratory of Entomology and b at ENEA laboratory. Asterisk (*) indicates significant differences from the isogenic cultivar in the first generation
Reproducibility between laboratories
The higher rate of intrinsic increase in the aphid population in the first generation on the cisgenic event A15-31 was observed in the laboratories at WUR (Fig. 2a; p = 0.0138) and at ENEA (Fig. 2b; p = 0.0243). However, at WUR, aphids generally had a lower intrinsic rate of increase in the second generation (Fig. 2a; p = 0.0223); whereas in ENEA, it was generally higher in the second generation (Fig. 2b; p = 0.0177).
Aphid survival
Probability of aphid survival over time also tended to be higher on the GM events as compared to the non-transformed Désirée, though only in the first generation significant differences were observed in one transgenic event A13-13 (p = 0.028) with a single R gene and one transgenic event with two R genes, A16-02 (p = 0.039) (Fig. 3a). In the second generation, there were no longer differences between the probabilities of survival of aphids on GM events compared to non-transformed Désirée (Fig. 3b).

Probability of aphid survival per generation on Solanum tuberosum isogenic cultivar Désirée, compared to several genetically modified events. (Color figure online)
No differences were found in the survival of aphids on Désirée compared to A15-31 or A15-45 at either WUR or ENEA (Appendix of Table 1).
Baseline comparison with commercially available cultivars
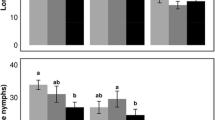
In order to put these results into context of the differences found among conventionally bred and commercially available potato varieties, we tested aphids on three different varieties known to differ in level of resistance against P. infestans. Compared to Désirée, on the other three conventionally bred varieties, aphids had a lower intrinsic rate of increase (Désirée vs. Bintje: p = 0.002, and Désirée compared to Première and Sarpo Mira: p < 0.0001). When put into context of the conventionally bred varieties, there was no longer any difference between aphid rate of increase on the cisgenic event (A15-31) and Désirée (p = 0.1282). Although not different from the isogenic P. infestans-susceptible Désirée, the highly resistant cisgenic event (A15-31) also did not differ from the highly susceptible conventional variety Bintje (p = 0.1198) but aphids had significantly higher intrinsic rate of increase than on the highly P. infestans-resistant conventional variety Sarpo Mira (p < 0.0001; Fig. 4).

Mean aphid intrinsic rate of increase (±SE) on Solanum tuberosum isogenic cultivar Désirée, compared to a cisgenically modified event (A15-31), and three conventional cultivars Bintje, Première and Sarpo Mira. Different letters indicate significant differences between bars
Probability of aphid survival did not differ between Désirée, Bintje and the cisgenic-resistant event A15-31 (Désirée vs. Bintje, p = 0.2919; Désirée vs. A15-31, p = 0.2225). However, aphid survival was significantly lower on Première (p = 0.0096) and Sarpo Mira (p < 0.0001; Fig. 5).

Probability of aphid survival per generation on Solanum tuberosum isogenic cultivar Désirée, compared to cisgenically modified event (A15-31), and three conventional cultivars Bintje, Première and Sarpo Mira. Bold red and blue bold lines indicate significant differences from the isogenic cultivar (Désirée). (Color figure online)
Discussion
Influence of selection markers, number of R genes, collateral effects and endpoint choice on detection of non-target effects
The results of our experiments show that genetic modification in potato for resistance to P. infestans through R gene insertion may have effects on non-target aphids in the first generation, yet these effects were no longer evident in the second generation of aphids. These effects cannot be attributed to marker gene use in the modification, since intrinsic rate of increase was higher both in a cisgenic and in a transgenic event. The differences found between events cannot be attributed to the number of R genes either, since survival probability was increased in events with both one and two R genes.
Interestingly, on the same event intrinsic rate of increase could be significantly higher, whereas survival did not differ. In our findings, significant effects on aphid life-history traits were never seen on both events transformed with the same construct. This brings to light the issue that detection of non-target effects depends on the measured endpoint (Charleston and Dicke 2008; Lövei et al. 2009). For example, in the case of the variety Bintje, it differed from Désirée in terms of aphid intrinsic rate of increase, yet had similar survival probability. Similarly, aphids on Désirée plants transformed to express enhanced chitinolytic activities showed increased population growth, while survival probability did not differ (Saguez et al. 2005). In the GM events, aphids had higher intrinsic rates of increase on A15-31 and A13-17, yet these were not the same events on which survival differed. Therefore, it is important to carefully select biologically relevant endpoints for testing in the greenhouse that can most closely translate to effect differences in the field. Considering several selected measurement endpoints when testing for environmental risk and non-target testing can be misleading if not all endpoints lead to differences in the same events. This considered, for the events tested at both WUR and ENEA, we came to comparable results with regard to both endpoints. Testing multiple endpoints in several events considerably strengthens the reliability of results of early tier risk assessments, but would require separate testable hypotheses and protection goals specific to each in order to reliably inform the assessment.
The location of the inserted R gene in the genome is the only difference between events transformed with the same construct. Since one event can influence aphid life-history traits, whereas another does not, we conclude that these are unintended effects associated with the location of insertion. These are known as position effects (Miki et al. 2009). These insertions may have occurred in a location that can affect interactions with insects such as defence response pathways. However, insertions usually result in loss of function rather than gain of function (Wang 2008). Loss of function effects are complemented by the three remaining copies in the tetraploid potato genome. A more likely explanation of the observed position effects could be a difference in expression level of the inserted R gene. Substantial differences in the expression level of the Rpi-vnt1 gene are observed among different transformation events (J.H. Vossen, unpublished data). In this case, overexpression of a late blight R gene may have a trade-off with resistance to aphids. Generally, these results emphasize the usefulness of a pre-screening for position effects on relevant non-target insects before proceeding with an entire environmental risk assessment on a single modified event. These early tests can help detect possible position effects resulting from genetic modification.
Detection of non-target effects over two insect generations
Our findings show that differences could be detected in the first generation of aphids feeding on GM events; however, these differences had disappeared in the second generation of aphids. Although transgenic resistance based on the expression of Bacillus thuringiensis (Bt) proteins has a very different mode of action, Rhopalosiphum padi aphids on Bt (transgenic) maize had higher performance in the first generation (Lumbierres et al. 2004). Aphis gossypii aphids also had higher intrinsic rates of increase on Bt cotton in the first, but not in the second or third generation (Liu et al. 2005). Since aphids were reared on the untransformed cultivar Désirée, it is possible that the effects seen in the first generation are a consequence of the aphids switching host plants rather than an effect of the transformation itself. This possibility can be tested in future experiments by rearing insects on an alternative host or on each of the test events separately.
The second generation of aphids was kept on the sample plants at WUR, yet at ENEA second-generation aphids were transferred to new plants. Although there were no differences in intrinsic rates of increase between genotypes detected in the second generation of aphids in either laboratory, the difference in performance of the second-generation aphids between experiments conducted at ENEA and Wageningen may have been caused by induced defence mechanisms since both generations were kept on the same plant in Wageningen. Feeding by conspecifics on the same plant can have negative effects on the life-history traits of M. persicae, due to systemic defence mechanisms of the plant (Dugravot et al. 2007).
Aphids are considered as good model organisms for understanding epigenetic effects (Srinivasan and Brisson 2012). The formation of winged offspring is a well-known epigenetic effect in aphids and can be triggered both pre- and post-natally by appropriate environmental cues (Brisson 2010; Sutherland 1969). The formation of sexual aphids is another example of epigenetic responses (Halkett et al. 2004). Although rapid epigenetic responses to changes in plant quality have not yet been studied, this could be an explanation for the changes we observed between rates of increase in two generations.
In aphids it is a natural situation for two generations (or more) to be present on the same plant. In our statistical models, we found in some cases that survival and rate of increase are significantly affected by the interaction of the factors ‘generation’ and ‘event’, which may also explain why observed effects are significant in the first, though not in the second generation. Additionally, the present paper allowed the set-up of a protocol that proved to be sensitive and reproducible and can be suggested as a standard for in planta studies with aphids in ERA.
Significant effects in non-target tests should be compared to variation among conventionally bred varieties
Furthermore, our results point to the importance of comparing the differences found between GM events and the non-transformed variety to the variation among available conventional varieties in the agro-ecosystem. The concept of baseline variation has been documented before and is considered a necessary part of environmental risk assessment (EFSA 2010; Houshyani 2012). We show that when conventional cultivars are included in the comparison of the intrinsic rate of increase, the non-transformed and GM events no longer significantly differ, and rather the variation between conventionally bred varieties is much greater than between a non-transformed cultivar and derived GM events. Though significant effects may be found between the GM potato and its non-transformed progenitor when compared pairwise, this may be insignificant compared to the extent of variation already found between different conventionally bred potato varieties. In the case of our blight-resistant events, despite our sensitive assays, no biological relevance was detected for the non-target effect on aphids, since it proved to be in the range of effects present among available commercial varieties.
Author contributions
JL, SA and JJAvL designed the research. JL, FB, PB, SM and EM conducted experiments. JHV contributed materials. JL and SA analysed data. JL, JHV, SA and JJAvL wrote, and all authors read and approved the manuscript.
References
Andow DA, Hilbeck A (2004) Science-based risk assessment for nontarget effects of transgenic crops. Bioscience 54:637–649
Andow DA, Zwahlen C (2006) Assessing environmental risks of transgenic plants. Ecol Lett 9:196–214
Andow DA et al (2013) An ecologically-based method for selecting ecological indicators for assessing risks to biological diversity from genetically-engineered plants. J Biosaf 22:141–156
Arpaia S (2010) Genetically modified plants and “non-target” organisms: analysing the functioning of the agro-ecosystem. Collect Biosaf Rev 5:12–80
Birch ANE, Griffiths BS, Caul S, Thompson J, Heckmann LH, Krogh PH, Cortet J (2007) The role of laboratory, glasshouse and field scale experiments in understanding the interactions between genetically modified crops and soil ecosystems: a review of the ECOGEN project. Pedobiologia 51:251–260
Borenstein M, Rothstein H, Cohen J (2001) Power and precision (version 2). Biostat Inc., Englewood
Brisson JA (2010) Aphid wing dimorphisms: linking environmental and genetic control of trait variation. Philos Trans R Soc B Biol Sci 365:605–616
Burnham KP, Anderson DR, Huyvaert KP (2010) AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35
Carstens K et al (2014) Surrogate species selection for assessing potential adverse environmental impacts of genetically engineered insect-resistant plants on non-target organisms. GM Crops Food 5:11–15
Charleston DS, Dicke M (2008) Designing experimental protocols to investigate the impact of GM crops on non-target arthropods. COGEM research report CGM2008-1, Bilthoven
Dugravot S, Brunissen L, Létocart E, Tjallingii WF, Vincent C, Giordanengo P, Cherqui A (2007) Local and systemic responses induced by aphids in Solanum tuberosum plants. Entomol Exp Appl 123:271–277
EFSA (2010) Panel on genetically modified organisms, guidance on the environmental risk assessment of genetically modified plants. EFSA J 8:1879
Gillund FT, Nordgaard L, Bøhn T, Wikmark OG, Konestabo HS, Hilbeck A (2013) Selection of nontarget testing organisms for ERA of GM potato with increased resistance to late blight. Potato Res 56:293–324
Haesaert G et al (2015) Transformation of the potato variety Désirée with single or multiple resistance genes increases resistance to late blight under field conditions. Crop Prot 77:163–175
Halkett F, Harrington R, Hullé M, Kindlmann P, Menu F, Rispe C, Plantegenest M (2004) Dynamics of production of sexual forms in aphids: theoretical and experimental evidence for adaptive “coin-flipping” plasticity. Am Nat 163:E112–E125
Han P, Concepcion Velasco-Hernandez M, Ramirez-Romero R, Desneux N (2016) Behavioral effects of insect-resistant genetically modified crops on phytophagous and beneficial arthropods: a review. J Pest Sci 89:859–883
Haverkort A et al (2016) Durable late blight resistance in potato through dynamic varieties obtained by cisgenesis: scientific and societal advances in the DuRPh project. Potato Res 59:35–66
Houshyani B (2012) Application of omics technologies for environmental risk assessment of genetically modified plants: Arabidopsis and modified defence mechanisms as a model study. Ph.D. thesis, Wageningen University
Kamoun S, Huitema E, Vleeshouwers VGAA (1999) Resistance to oomycetes: a general role for the hypersensitive response? Trends Plant Sci 4:196–200
Kennedy JS, Day MF, Eastop VF (1962) A conspectus of aphids as vectors of plant viruses. Commonwealth Institute of Entomology, London
Kos M, van Loon JJA, Dicke M, Vet LEM (2009) Transgenic plants as vital components of integrated pest management. Trends Biotechnol 27:621–627
Liu XD, Zhai BP, Zhang XX, Zong JM (2005) Impact of transgenic cotton plants on a non-target pest, Aphis gossypii Glover. Ecol Entomol 30:307–315
Lövei GL, Andow DA, Arpaia S (2009) Transgenic insecticidal crops and natural enemies: a detailed review of laboratory studies. Environ Entomol 38:293–306
Lumbierres B, Albajes R, Pons X (2004) Transgenic Bt maize and Rhopalosiphum padi (Hom., Aphididae) performance. Ecol Entomol 29:309–317
Majerus MEN (1994) Ladybirds. Harper Collins Academic, London
Meissle M, Álvarez-Alfageme F, Malone LA, Romeis J (2012) Establishing a database of bio-ecological information on non-target arthropod species to support the environmental risk assessment of genetically modified crops in the EU. EFSA J 9:1–170
Miki B, Abdeen A, Manabe Y, MacDonald P (2009) Selectable marker genes and unintended changes to the plant transcriptome. Plant Biotechnol J 7:211–218
Müller CB, Adriaanse ICT, Belshaw R, Godfray HCJ (1999) The structure of an aphid–parasitoid community. J Anim Ecol 68:346–370
Ng JCK, Perry KL (2004) Transmission of plant viruses by aphid vectors. Mol Plant Pathol 5:505–511
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Radcliffe EB (1982) Insect pests of potato. Annu Rev Entomol 27:173–204
Raj BT (1989) Seasonal abundance of natural enemies of aphids infesting potato crop. J Aphidol 3:157–161
Riedel J, Romeis J, Meissle M (2016) Update and expansion of the database of bio-ecological information on non-target arthropod species established to support the environmental risk assessment of genetically modified crops in the EU. EFSA Support Publ 13:956E
Romeis J et al (2011) Recommendations for the design of laboratory studies on non-target arthropods for risk assessment of genetically engineered plants. Transgenic Res 20:1–22
Romeis J, Raybould A, Bigler F, Candolfi MP, Hellmich RL, Huesing JE, Shelton AM (2013) Deriving criteria to select arthropod species for laboratory tests to assess the ecological risks from cultivating arthropod-resistant genetically engineered crops. Chemosphere 90:901–909
Romeis J et al (2014) Potential use of an arthropod database to support the non-target risk assessment and monitoring of transgenic plants. Transgenic Res 23:995–1013
Saguez J et al (2005) Unexpected effects of chitinases on the peach-potato aphid (Myzus persicae Sulzer) when delivered via transgenic potato plants (Solanum tuberosum Linné) and in vitro. Transgenic Res 14:57–67
Scholte E-J, Dicke M (2005) Effects of insect-resistant transgenic crops on non-target arthropods: first step in pre-market risk assessment studies. COGEM research report CGM 2005-6, Bilthoven
Srinivasan DG, Brisson JA (2012) Aphids: a model for polyphenism and epigenetics. Genet Res Int 2012:12
Sutherland ORW (1969) The role of crowding in the production of winged forms by two strains of the pea aphid, Acyrthosiphon pisum. J Insect Physiol 15:1385–1410
The European Cultivated Potato Database (2015) Scottish agricultural science agency, European Cooperative Programme for crop genetic resources
Van Emden HF, Eastop VF, Hughes RD, Way MJ (1969) The ecology of Myzus persicae. Annu Rev Entomol 14:197–270
Vleeshouwers VGAA, van Dooijeweert W, Govers F, Kamoun S, Colon LT (2000) The hypersensitive response is associated with host and nonhost resistance to Phytophthora infestans. Planta 210:853–864
Vleeshouwers VGAA et al (2011) Understanding and exploiting late blight resistance in the age of effectors. Ann Rev Phytopathol 49:507–531
Wang YH (2008) How effective is T-DNA insertional mutagenesis in Arabidopsis? J Biochem Technol 1:11–20
Wyatt I, White P (1977) Simple estimation of intrinsic increase rates for aphids and tetranychid mites. J Appl Ecol 14:757–766
Acknowledgements
Many thanks to the partners of the AMIGA project (Assessing and Monitoring the Impacts of Genetically modified plants on Agro-ecosystems); whose main aim is to provide scientific data to support the foundations of risk assessment protocols for GM crops in European environments.
Funding
This study was funded by the European Commission as a Large-scale Integrating Project within FP 7 under Grant Agreement No. 289706.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors, Jenny Lazebnik, Salvatore Arpaia, Ferdinando Baldacchino, Paolo Banzato, Stefania Moliterni, Jack H. Vossen, Els M. van de Zande and Joop J.A. van Loon declare that they have no conflict of interest.
Ethical approval
All applicable international, national and/or institutional guidelines for the care and use of animals were followed. This article does not contain any studies with human participants performed by any of the authors.
Additional information
Communicated by Y. Gao.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Lazebnik, J., Arpaia, S., Baldacchino, F. et al. Effects of a genetically modified potato on a non-target aphid are outweighed by cultivar differences. J Pest Sci 90, 855–864 (2017). https://doi.org/10.1007/s10340-017-0831-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-017-0831-6