Point Mutations in the 14-α Sterol Demethylase Cyp51A or Cyp51C Could Contribute to Azole Resistance in Aspergillus flavus

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Molecular Identification
2.2. Antifungal Susceptibility Testing (AFST)
2.3. Susceptibility Testing to 14-α Sterol Demethylase Inhibitors (DMIs)
2.4. PCR Amplification and Sequence Analysis of the cyp51A, cyp51B and cyp51C Genes
2.5. RNA Isolation, Reverse Transcription and Quantitative PCR (RT-qPCR)
2.6. Sequence Computer Analysis
2.7. Phylogenetic Analysis
3. Results
3.1. Antifungal Susceptibility Testing

{kind=link}
{kind=link}
| Strains | MICs Ranges (mg/L) | ||||
|---|---|---|---|---|---|
| AmB | ITC | VRC | POS | ISV | |
| ATCC2004304 | 1 | 0.5 | 2 | 0.125 | 1 |
| CM7668 * | 0.5 | >8 | 0.25 | 1–2 | 0.5 |
| CM8087 | 8–>16 | 4–8 | 8 | 0.5 | 8 |
| CM9165 | 1 | 0.25 | 0.5 | 0.125 | 0.25 |
| CM9174 | 2 | 0.5–1 | 1–4 | 0.125–0.25 | 2–4 |
| CM9189 | 1 | 0.5 | 1 | 0.25 | 1 |
| CM9195 | 1 | 0.25 | 1 | 0.125 | 1 |
| CM9228 | 1 | 0.5 | 0.5 | 0.25 | 2 |
| CM9267 | 2 | 0.5 | 1 | 0.25 | 1 |
| CM9298 | 1 | 0.5 | 1 | 0.125 | 1 |
| CM9326 * | 1–2 | 4–8 | 8 | 1–2 | >8 |
| CM9329 | 2 | 0.5 | 1 | 0.25 | 1 |
| CM9331 | 1 | 0.5 | 1 | 0.25 | 2 |
| CM9684 | 0.5 | 1 | >8 | 0.25 | 2 |
| TP642 | 1 | 1 | 2 | 0.25 | 2 |
| TP968 | 1 | 1–2 | 4 | 0.5–1 | 4 |
| TP992 | 1 | 0.5 | 2 | 0.25 | 1 |
| TP1004 | 1 | 1 | 4 | 0.5 | 4 |
| TP1115 | 1 | 1 | 4 | 0.5 | 4 |
| TP1179 | 1 | 1 | 4 | 0.5 | 4 |
3.2. Susceptibility Testing to 14-α Sterol Demethylase Inhibitors (DMIs) Fungicides
3.3. Sequence Analysis of cyp51A, cyp51B and cyp51C Genes
3.3.1. Aspergillus flavus Cyp51A
3.3.2. Aspergillus flavus Cyp51B
3.3.3. Aspergillus flavus Cyp51C
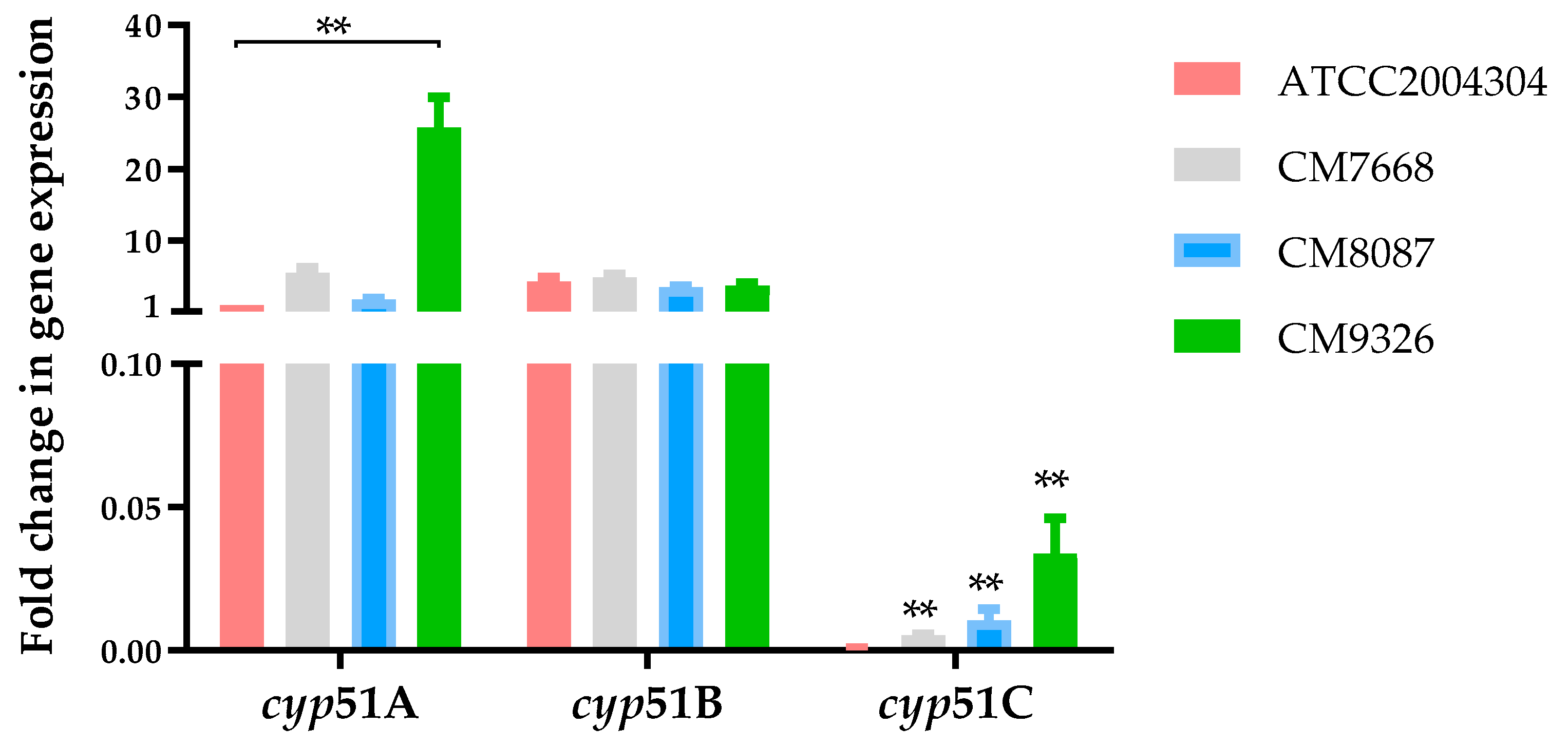
3.4. Aspergillus flavus cyp51 Genes Expression
3.5. Sequence Analysis: Alignments and Similarity
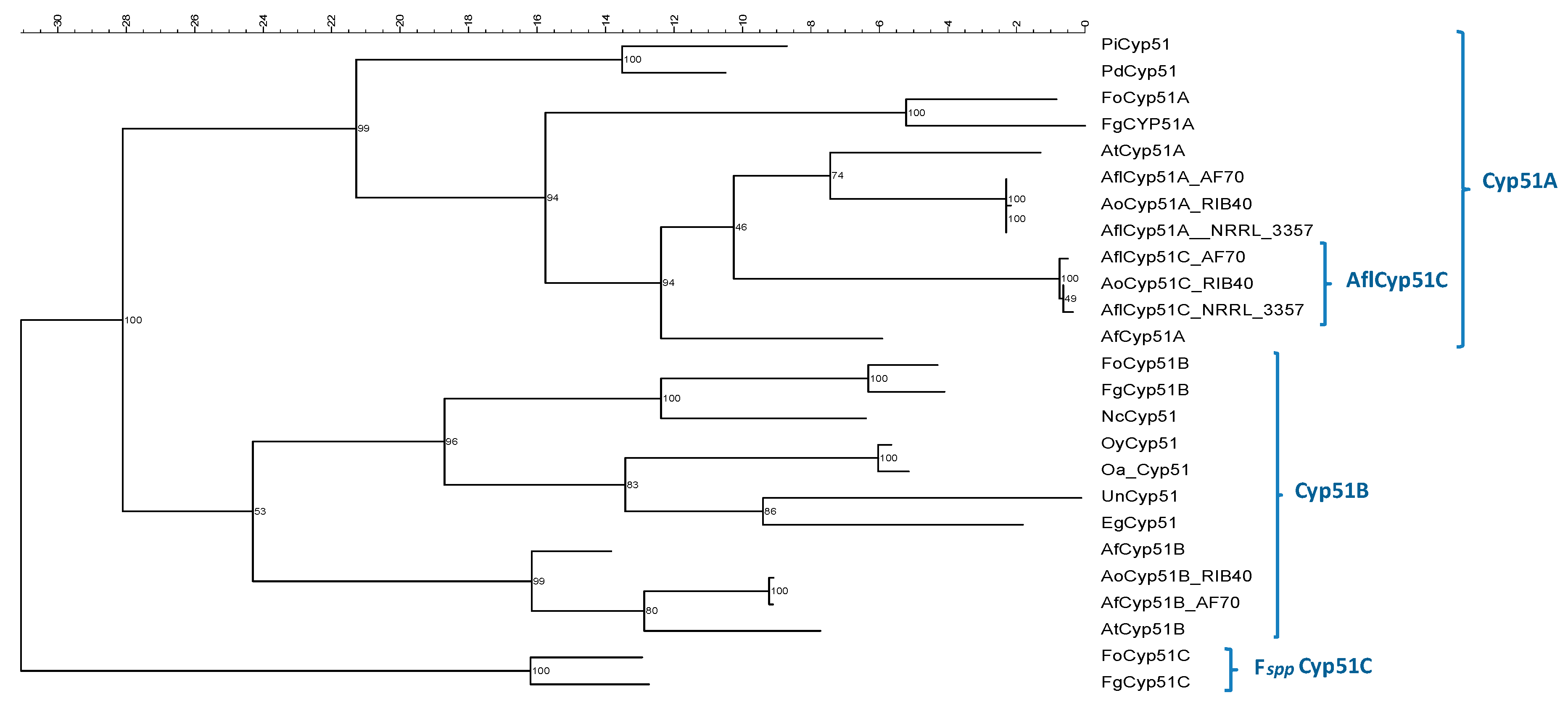
3.6. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Richardson, M.; Lass-Flörl, C. Changing epidemiology of systemic fungal infections. Clin. Microbiol. Infect. 2008, 14, 5–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulussen, C.; Hallsworth, J.E.; Álvarez-Pérez, S.; Nierman, W.C.; Hamill, P.G.; Blain, D.; Rediers, H.; Lievens, B. Ecology of aspergillosis: Insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb. Biotechnol. 2017, 10, 296–322. [Google Scholar] [CrossRef] [Green Version]
- Low, C.Y.; Rotstein, C. Emerging fungal infections in immunocompromised patients. F1000 Med. Rep. 2011, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nivoix, Y.; Velten, M.; Letscher-Bru, V.; Moghaddam, A.; Natarajan-Amé, S.; Fohrer, C.; Lioure, B.; Bilger, K.; Lutun, P.; Marcellin, L.; et al. Factors Associated with Overall and Attributable Mortality in Invasive Aspergillosis. Clin. Infect. Dis. 2008, 47, 1176–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taccone, F.S.; Van den Abeele, A.M.; Bulpa, P.; Misset, B.; Meersseman, W.; Cardoso, T.; Paiva, J.A.; Blasco-Navalpotro, M.; De Laere, E.; Dimopoulos, G.; et al. AspICU Study Investigators. Epidemiology of invasive aspergillosis in critically ill patients: Clinical presentation, underlying conditions, and outcomes. Crit. Care 2015, 19, 7. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, P.; Soubani, A. Invasive pulmonary aspergillosis in patients with COPD: A report of five cases and systematic review of the literature. Chron. Respir. Dis. 2008, 5, 19–27. [Google Scholar] [CrossRef]
- Garcia-Giraldo, A.M.; Mora, B.L.; Loaiza-Castaño, J.M.; Cedano, J.A.; Rosso, F. Invasive fungal infection by Aspergillus flavus in immunocompetent hosts: A case series and literature review. Med. Mycol. Case Rep. 2018, 23, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Manavathu, E.K.; Chandrasekar, P.H. Aspergillus flavus: An emerging non-fumigatus Aspergillus species of significance. Mycoses 2009, 52, 206–222. [Google Scholar] [CrossRef]
- Hedayati, M.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Mellado, E.; Peláez, T.; Pemán, J.; Zapico, S.; Alvarez, M.; Rodríguez-Tudela, J.L.; Cuenca-Estrella, M.; FILPOP Study Group. Population-based survey of filamentous fungi and antifungal resistance in Spain (FILPOP Study). Antimicrob. Agents Chemother. 2013, 57, 3380–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Rivero-Menéndez, O.; Ayats, J.; Castro, C.; García-Rodríguez, J.; Goterris-Bonet, L.; Ibáñez-Martínez, E.; Linares-Sicilia, M.J.; Martin-Gomez, M.T.; et al. The FILPOP2 Project from GEMICOMED (SEIMC) and REIPI. Molecular Identification and Susceptibility Testing of Molds Isolated in a Prospective Surveillance of Triazole Resistance in Spain (FILPOP2 Study). Antimicrob. Agents Chemother. 2018, 62, e00358-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudramurthy, S.M.; Paul, R.A.; Chakrabarti, A.; Mouton, J.W.; Meis, J.F. Invasive Aspergillosis by Aspergillus flavus: Epidemiology, Diagnosis, Antifungal Resistance, and Management. J. Fungi 2019, 5, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and Management of Aspergillus Diseases: Executive Summary of the 2017 ESCMID-ECMM-ERS Guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14alpha-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophys. Acta 2007, 1770, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole Resistance in Aspergillus Species: An Emerging Problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef]
- Zhang, J.; van den Heuvel, J.; Debets, A.J.M.; Verweij, P.E.; Melchers, W.J.G.; Zwaan, B.J.; Schoustra, S.E. Evolution of cross-resistance to medical triazoles in Aspergillus fumigatus through selection pressure of environmental fungicides. Proc. Biol. Sci. 2017, 284, 20170635. [Google Scholar] [CrossRef] [Green Version]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant. Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef]
- Varga, J.; Frisvad, J.C.; Samson, R.A. Two new aflatoxin producing species, and an overview of Aspergillus section Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef]
- Mellado, E.; Diaz-Guerra, T.M.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Identification of two different 14-a sterol demethylase-related genes (cyp51A and cyp51B) in Aspergillus fumigatus and other Aspergillus species. J. Clin. Microbiol. 2001, 39, 2431–2438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The Fungal CYP51s: Their Functions, Structures, Related Drug Resistance, and Inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Won, E.J.; Joo, M.Y.; Park, Y.J.; Kim, S.H.; Shin, M.G.; Shin, J.H. Microsatellite typing and resistance mechanism analysis of voriconazole-resistant Aspergillus flavus isolates in South Korean hospitals. Antimicrob. Agents Chemother. 2019, 63, e01610-18. [Google Scholar] [CrossRef] [Green Version]
- Krishnan-Natesan, S.; Chandrasekar, P.H.; Alangaden, G.J.; Manavathu, E.K. Molecular characterisation of cyp51A and cyp51B genes coding for P450 14α-lanosterol demethylases A (CYP51Ap) and B (CYP51Bp) from voriconazole-resistant laboratory isolates of Aspergillus flavus. Int. J. Antimicrob. Agents. 2008, 32, 519–524. [Google Scholar] [CrossRef]
- Liu, W.; Sun, Y.; Chen, W.; Liu, W.; Wan, Z.; Bu, D.; Li, R. The T788G mutation in the cyp51C gene confers voriconazole resistance in Aspergillus flavus causing aspergillosis. Antimicrob. Agents Chemother. 2012, 56, 2598–2603. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.A.; Rudramurthy, S.M.; Meis, J.F.; Mouton, J.W.; Chakrabarti, A. A Novel Y319H Substitution in CYP51C Associated with Azole Resistance in Aspergillus flavus. Antimicrob. Agents Chemother. 2015, 59, 6615–6619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, C.; Kumar, R.; Kumar, N.; Masih, A.; Gupta, D.; Chowdhary, A. Investigation of Multiple Resistance Mechanisms in Voriconazole-Resistant Aspergillus flavus Clinical Isolates from a Chest Hospital Surveillance in Delhi, India. Antimicrob. Agents Chemother. 2018, 62, e01928-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.M.; Cohen, J.; Holden, D.W. An Aspergillus fumigatus alkaline protease mutant constructed by gene disruption is deficient in extracellular elastase activity. Mol. Microbiol. 1992, 6, 1663–1671. [Google Scholar] [CrossRef]
- Alcazar-Fuoli, L.; Mellado, E.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Aspergillus section Fumigati: Antifungal susceptibility patterns and sequence-based identification. Antimicrob. Agents Chemother. 2008, 52, 1244–1251. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W.W. European Committee on Antimicrobial Susceptibility Testing Subcommittee on Antifungal Susceptibility Testing (EUCAST-AFST). EUCAST technical note on Aspergillus and amphotericin B, itraconazole, and posaconazole. Clin. Microbiol. Infect. 2012, 18, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W.W. Breakpoints for antifungal agents: An update from EUCAST focussing on echinocandins against Candida spp. and triazoles against Aspergillus spp. Drug Resist. Updat. 2013, 16, 81–95. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs for Antifungal Agents, Version 10.0. 2020. Available online: http://www.eucast.org/astoffungi/clinicalbreakpointsforantifungals/ (accessed on 16 April 2020).
- Diaz-Guerra, T.M.; Mellado, E.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. A point mutation in the 14alpha-sterol demethylase gene cyp51A contributes to itraconazole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2003, 47, 1120–1124. [Google Scholar] [CrossRef] [Green Version]
- Cove, D.J. The induction and repression of nitrate reductase in the fungus Aspergillus nidulans. Biochim. Biophys. Acta 1966, 113, 51–56. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Hess, J.; Goldman, N. Addressing inter-gene heterogeneity in maximum likelihood phylogenomic analysis: Yeasts revisited. PLoS ONE 2011, 6, e22783. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Menendez, O.; Soto-Debran, J.C.; Medina, N.; Lucio, J.; Mellado, E.; Alastruey-Izquierdo, A. Molecular Identification, Antifungal Susceptibility Testing, and Mechanisms of Azole Resistance in Aspergillus Species Received within a Surveillance Program on Antifungal Resistance in Spain. Antimicrob. Agents Chemother. 2019, 63, e00865-19. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, M.; Izumikawa, K.; Hirano, K.; Ide, S.; Mihara, T.; Hosogaya, N.; Takazono, T.; Morinaga, Y.; Nakamura, S.; Kurihara, S.; et al. Correlation between triazole treatment history and susceptibility in clinically isolated Aspergillus fumigatus. Antimicrob. Agents Chemother. 2012, 56, 4870–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef] [Green Version]
- Nierman, W.C.; Yu, J.; Fedorova-Abrams, N.D.; Losada, L.; Cleveland, T.E.; Bhatnagar, E.; Bennett, J.W.; Dean, R.; Payne, G.A. Genome Sequence of Aspergillus flavus NRRL 3357, a Strain That Causes Aflatoxin Contamination of Food and Feed. Genome Announc. 2015, 3, e00168-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of Azole-Resistant Aspergillus fumigatus Strains due to Agricultural Azole Use Creates an Increasing Threat to Human Health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Hollomon, D. Does Agricultural Use of Azole Fungicides Contribute to Resistance in the Human Pathogen Aspergillus fumigatus? Pest. Manag. Sci. 2017, 73, 1987–1993. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.; Meis, J.F. Azole Resistance in Aspergillus fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, N.J.; Cools, H.J.; Sierotzki, H.; Shaw, M.W.; Knogge, W.; Kelly, S.L.; Kelly, D.E.; Fraaije, B.A. Paralog re-emergence: A novel, historically contingent mechanism in the evolution of antimicrobial resistance. Mol. Biol. Evol. 2014, 7, 1793–1802. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Urban, M.; Parker, J.E.; Brewer, H.C.; Kelly, S.L.; Hammond-Kosack, K.E.; Fraaije, B.A.; Liu, X.; Cools, H.J. Characterization of the sterol 14α-demethylases of Fusarium graminearum identifies a novel genus-specific CYP51 function. New Phytol. 2013, 198, 821–835. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Yan, L.; Liang, W.; Yang, Q. Paralogous Cyp51s Mediate the Differential Sensitivity of Fusarium Oxysporum to Sterol Demethylation Inhibitors. Pest. Manag. Sci. 2019, 75, 396–404. [Google Scholar] [CrossRef]
- Payne, G.A.; Nierman, W.C.; Wortman, J.R.; Pritchard, B.L.; Brown, D.; Dean, R.A.; Bhatnagar, D.; Cleveland, T.E.; Machida, M.; Yu, J. Whole genome comparison of Aspergillus flavus and A. oryzae. Med. Mycol. 2006, 44, S9–S11. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.K.; Mack, B.M.; Moore, G.G.; Downey, D.L.; Lebar, M.D.; Joardar, V.; Losada, L.; Yu, J.; Nierman, W.C.; Bhatnagar, D. Whole genome comparison of Aspergillus flavus L-morphotype strain NRRL 3357 (type) and S-morphotype strain AF70. PLoS ONE 2018, 13, e0199169. [Google Scholar] [CrossRef] [PubMed]
- Camps, S.M.; van der Linden, J.W.; Li, Y.; Kuijper, E.J.; van Dissel, J.T.; Verweij, P.E.; Melchers, W.J. Rapid induction of multiple resistance mechanisms in Aspergillus fumigatus during azole therapy: A case study and review of the literature. Antimicrob. Agents Chemother. 2012, 56, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Madison, V.; Chau, A.S.; Loebenberg, D.; Palermo, R.E.; McNicholas, P.M. Three-dimensional models of wild-type and mutated forms of cytochrome P450 14alpha-sterol demethylases from Aspergillus fumigatus and Candida albicans provide insights into posaconazole binding. Antimicrob. Agents Chemother. 2004, 48, 568–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabas, Y.; Xess, I.; Bakshi, S.; Mahapatra, M.; Seth, R. Emergence of Azole-Resistant Aspergillus fumigatus from Immunocompromised Hosts in India. Antimicrob. Agents Chemother. 2018, 62, e02264-17. [Google Scholar] [CrossRef] [Green Version]
- Ballard, E.; Melchers, W.J.G.; Zoll, J.; Brown, A.J.P.; Verweij, P.E.; Warris, A. In-host microevolution of Aspergillus fumigatus: A phenotypic and genotypic analysis. Fungal Genet. Biol. 2018, 113, 1–13. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Seyedmousavi, S.; Dhaliwal, M.; Chakrabarti, A.; Meis, J.F.; Mouton, J.W. Pharmacodynamics of Voriconazole against Wild-Type and Azole-Resistant Aspergillus flavus Isolates in a Nonneutropenic Murine Model of Disseminated Aspergillosis. Antimicrob. Agents Chemother. 2016, 61, e01491-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, R.A.; Rudramurthy, S.M.; Dhaliwal, M.; Singh, P.; Ghosh, A.K.; Kaur, H.; Varma, S.; Agarwal, R.; Chakrabarti, A. Magnitude of voriconazole resistance in clinical and environmental isolates of Aspergillus flavus and investigation into the role of multidrug efflux pumps. Antimicrob. Agents Chemother. 2018, 62, e01022-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Cantero, A.; López-Fernández, L.; Guarro, J.; Capilla, J. Azole resistance mechanisms in Aspergillus: Update and recent advances. Int. J. Antimicrob. Agents. 2020, 55, 105807. [Google Scholar] [CrossRef]
- Forastiero, A.; Mesa-Arango, A.C.; Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Bernal-Martinez, L.; Pelaez, T.; Lopez, J.F.; Grimalt, J.O.; Gomez-Lopez, A.; Cuesta, I.; et al. Candida tropicalis antifungal cross-resistance is related to different azole target (Erg11p) modifications. Antimicrob. Agents Chemother. 2013, 57, 4769–4781. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| A. flavus strains | MICs Ranges (mg/L) | |||||||
|---|---|---|---|---|---|---|---|---|
| Imidazole | Triazole | |||||||
| Imazalil | Prochloraz | Metconazole | Tebuconazole | Epoxiconazole | Bromuconazole | Difenoconazole | Myclobutanil | |
| ATCC2004304 | 1–2 | 2–4 | 4–8 | 8 | 8–16 | 4–16 | 8–>32 | >32 |
| CM7668 | 0.25 | 1 | 1 | 1 | 0.5–1 | 0.25–1 | 0.5–1 | 4 |
| CM8087 | 16 | >32 | >32 | >32 | >32 | >32 | >32 | >32 |
| CM9165 | 2 | 1 | 8 | 8 | 8 | 8 | 16 | >32 |
| CM9174 | 4 | 2 | 16 | 16 | 32 | 8 | 16 | >32 |
| CM9189 | 2 | 1 | 8 | 8 | 8 | 8 | 16 | >32 |
| CM9195 | 2 | 1 | 8 | 4 | 8 | 8 | 16 | >32 |
| CM9228 | 2 | 1 | 8 | 4 | 8 | 8 | 16 | 32 |
| CM9267 | 2 | 2 | 8 | 8 | 8 | 16 | 16 | >32 |
| CM9298 | 2 | 2 | 8 | 4 | 8 | 8 | 16 | >32 |
| CM9326 | 32 | >32 | >32 | >32 | >32 | >32 | >32 | >32 |
| CM9329 | 1 | 1 | 4 | 4 | 8 | 8 | 16 | 32 |
| CM9331 | 2 | 1 | 8 | 8 | 16 | 8 | 32 | >32 |
| CM9684 | 8 | 2–4 | 16 | 16 | 16–>32 | 32–>32 | 16–>16 | >32 |
| TP642 | 8 | 8 | 16 | 16 | >32 | 32 | >32 | >32 |
| TP968 | 8 | 32 | 16 | 32 | >32 | 16 | 32 | >32 |
| TP992 | 2 | 2 | 8 | 4 | 16 | 8 | >32 | >32 |
| TP1004 | 8 | 8 | 16 | 16 | >32 | >32 | >32 | >32 |
| TP1115 | 8 | 8 | 16 | 16 | >32 | >32 | >32 | >32 |
| TP1179 | 8 | 8 | 16 | 32 | >32 | >32 | >32 | >32 |
| Strains | A * | Aspergillus flavus Cyp51A | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| c132t | c165t | c342t | g390a | t546c | g595a | c641t | t723c | c738t | t907c | t927c | g966t | a985g | c1065t | t1164c | c1368t | g1371a | ||
| F44 | P55 | G114 | K130 | F182 | A199T | P214L 1 | Y241 | N246 | L303 | P309 | K322N | T329A | H355 | P388 | N456 | L456 | ||
| CM7668 | R | c165t | g390a | t546c | c641t | t1164c | ||||||||||||
| CM8087 | R | c165t | g390a | t546c | a985g | t1164c | ||||||||||||
| CM8098 | S | c165t | g390a | t546c | a985g | t1164c | ||||||||||||
| CM9165 | S | c165t | t546c | t723c | t1164c | |||||||||||||
| CM9174 | S | c165t | g390a | t546c | a985g | t1164c | ||||||||||||
| CM9189 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9195 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9228 | S | t546c | t907c | g966t | t1164c | |||||||||||||
| CM9267 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9298 | S | c132t | c165t | c342t | g390a | t546c | t927c | t1164c | c1368t | |||||||||
| CM9326 | R | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9329 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9331 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| CM9684 | S | c165t | g390a | t546c | a985g | t1164c | ||||||||||||
| TPH642 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| TPH968 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| TPH992 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| TPH1004 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| TPH1115 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| TPH1179 | S | c165t | g390a | t546c | t1164c | |||||||||||||
| ATCC2004304 | S | c165t | g390a | t546c | g595a | c1065t | t1164c | c1368t | g1371a | |||||||||
| A. oryzae RIB40 | - | c165t | g390a | t546c | g595a | c1065t | t1164c | c1368t | g1371a | |||||||||
| A. flavus AF70 | - | c165t | g390a | t546c | c738t | t1164c | c1368t | |||||||||||
| Protein | F44 | P55 | G114 | K130 | F182 | A199T | P214L 1 | Y241 | N246 | L303 | P309 | K322N | T329A | H355 | P388 | N456 | L456 | |
| Strains | A * | Aspergillus flavus Cyp51B | |||
|---|---|---|---|---|---|
| c237t | c498t | t699c | a799g | ||
| C79 | I166 | N233 | K267 | ||
| CM7668 | R | ||||
| CM8087 | R | ||||
| CM9165 | S | c237t | c498t | a799g | |
| CM9174 | S | ||||
| CM9189 | S | c498t | |||
| CM9195 | S | c498t | |||
| CM9228 | S | c498t | |||
| CM9267 | S | ||||
| CM9298 | S | c498t | |||
| CM9326 | R | c498t | |||
| CM9329 | S | ||||
| CM9331 | S | c498t | |||
| CM9684 | S | t699c | |||
| TPH642 | S | ||||
| TPH968 | S | ||||
| TPH992 | S | ||||
| TPH1004 | S | ||||
| TPH1115 | S | ||||
| TPH1179 | S | ||||
| ATCC2004304 | S | ||||
| A. oryzae RIB40 | - | ||||
| A. flavus AF70 | - | c498t | a799g | ||
| Protein | C79 | I166 | N233 | K267 | |
| Strains | A * | Aspergillus flavus Cyp51C | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t161c | c174g | t718g | c748t | g760a | a761g | c826a | a853g | t876g | t894a | g915a | c978t | a1044g | a1046g | c1158t | g1196t | a1263g | a1267g | a1455g | ||
| M54T | G58G | S240A | R250ST | D254N | D254G | P276T | I285V | A292A | S298S | M305I | L326 | L348 | H349R 1 | S386 | S399I | E421 | N423D | V485V | ||
| CM7668 | R | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| CM8087 | R | t161c | c174g | t718g | ||||||||||||||||
| CM9165 | S | t161c | c174g | t718g | ||||||||||||||||
| CM9174 | S | t161c | c174g | t718g | ||||||||||||||||
| CM9189 | S | t161c | c174g | t718g | a761g | c826a | a853g | t876g | t894a | c1158t | a1263g | a1267g | ||||||||
| CM9195 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| CM9228 | S | t161c | c174g | t718g | c748t | g1196t | ||||||||||||||
| CM9267 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| CM9298 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| CM9326 | R | t161c | c174g | t718g | a1046g | |||||||||||||||
| CM9329 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| CM9331 | S | t161c | c174g | t718g | g760a | c826a | a853g | t876g | t894a | c1158t | a1263g | a1267g | ||||||||
| CM9684 | S | t161c | c174g | t718g | ||||||||||||||||
| TPH642 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| TPH968 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| TPH992 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| TPH1004 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| TPH1115 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| TPH1179 | S | t161c | c174g | t718g | a761g | c978t | a1044g | c1158t | a1263g | a1267g | ||||||||||
| ATCC ** | S | t161c | c174g | t718g | g760a | a853g | t876g | t894a | c1158t | a1455g | ||||||||||
| RIB40 *** | - | t161c | c174g | t718g | ||||||||||||||||
| AF70 | - | t161c | c174g | t718g | g760a | g915a | c1158t | a1263g | a1267g | |||||||||||
| Protein | M54T | G58G | S240A | R250ST | D254N | D254G | P276T | I285V | A292A | S298S | M305I | L326 | L348 | H349R 1 | S386 | S399I | E421 | N423D | V485V | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucio, J.; Gonzalez-Jimenez, I.; Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Pelaez, T.; Alcazar-Fuoli, L.; Mellado, E. Point Mutations in the 14-α Sterol Demethylase Cyp51A or Cyp51C Could Contribute to Azole Resistance in Aspergillus flavus. Genes 2020, 11, 1217. https://doi.org/10.3390/genes11101217
Lucio J, Gonzalez-Jimenez I, Rivero-Menendez O, Alastruey-Izquierdo A, Pelaez T, Alcazar-Fuoli L, Mellado E. Point Mutations in the 14-α Sterol Demethylase Cyp51A or Cyp51C Could Contribute to Azole Resistance in Aspergillus flavus. Genes. 2020; 11(10):1217. https://doi.org/10.3390/genes11101217
Chicago/Turabian StyleLucio, Jose, Irene Gonzalez-Jimenez, Olga Rivero-Menendez, Ana Alastruey-Izquierdo, Teresa Pelaez, Laura Alcazar-Fuoli, and Emilia Mellado. 2020. "Point Mutations in the 14-α Sterol Demethylase Cyp51A or Cyp51C Could Contribute to Azole Resistance in Aspergillus flavus" Genes 11, no. 10: 1217. https://doi.org/10.3390/genes11101217